硒通過調控小白菜幼苗根內NO與H2S信號誘導BrRbohG1/BrRbohG2表達
2015-06-09 14:20:33呂文靜李麗娜楊立飛
江蘇農業學報 2015年1期
關鍵詞:植物
陳 義, 陳 健, 呂文靜, 李麗娜, 楊立飛
(1.南京農業大學園藝學院,江蘇南京 210095;2.江蘇省農業科學院食品質量安全與檢測研究所,江蘇南京 210014)
硒通過調控小白菜幼苗根內NO與H2S信號誘導BrRbohG1/BrRbohG2表達
陳 義1,2, 陳 健2, 呂文靜1,2, 李麗娜1,2, 楊立飛1
(1.南京農業大學園藝學院,江蘇南京 210095;2.江蘇省農業科學院食品質量安全與檢測研究所,江蘇南京 210014)
為了研究硒對農作物脅迫的機理,本試驗以小白菜(Brassica rapa chinensis)幼苗為材料,研究了亞硒酸鈉(Na2SeO3)處理下,根中內源信號分子一氧化氮(NO)和硫化氫(H2S)的變化與BrRbohG1和BrRbohG2表達的關系。結果表明:0.03~0.46 mmol/L的Na2SeO3顯著抑制根的生長,并呈現濃度效應;隨著Na2SeO3處理濃度的升高,內源NO含量逐漸上升,而內源H2S逐漸下降,兩者呈顯著負相關;添加NO合成抑制劑(L-NMMA和Tungstate)或NO清除劑(cPTIO)能夠顯著抑制Na2SeO3對內源H2S的誘導作用;Na2SeO3處理能夠顯著誘導根內BrRbohG1和BrRbohG2的表達,分別添加L-NMMA、Tungstate、cPTIO、H2S供體NaHS均能顯著抑制Na2SeO3對上述兩基因表達的誘導作用。說明Na2SeO3處理下,內源NO介導了Na2SeO3對BrRbohG1和BrRbohG2的誘導表達,而H2S則可能抑制了兩個基因的表達;NO也可能作用于H2S信號的上游。
小白菜;亞硒酸鈉;硫化氫;一氧化氮;BrRbohG1/BrRbohG2
硒是動植物必需的微量元素之一,兼具營養、毒性、解毒三重生物學功能,是許多重要生物酶的主要成分[1-4]。研究發現通過對施硒濃度的調整,選擇蔬菜作為生物載體可以保證安全的有機硒攝入量[5-6]。因此近年來,通過施用富硒肥提高農作物中的硒含量已成為重要的農事操作之一。但這亦導致了大量硒在農業環境中的過量累積,成為一種新的重金屬污染源[7-8]。硒在動植物中的營養劑量和毒性劑量范圍非常狹窄[9]。低濃度的硒對植物的生長發育具有保護和促進作用,但是過量的硒會對植物產生脅迫作用,致使產量和品質下降[10]。目前為止,硒對植物產生脅迫的機理主要集中于其通過誘導活性氧(Reactive oxygen species,ROS)的過量產生,進而誘發氧化脅迫反應。但植物硒毒害深入的生化機理尚不明晰。已有研究結果顯示,由Rboh(Respiratory burst oxidase homologues)基因編碼的膜蛋白NADPH氧化酶是植物在多數非生物脅迫下產生ROS的主要來源[11-13]。但硒通過何種途徑誘導Rboh的表達還不清楚。
一氧化氮(Nitric oxide,NO)是調控植物響應環境脅迫的重要氣體信號分子[14]。最近的研究結果顯示,植物體內NO既能通過啟動防衛反應幫助植物抵抗重金屬脅迫[15],亦能通過促進ROS的過量產生進而介導重金屬的毒害效應[16-17]。已有研究結果顯示,過量硒在抑制擬南芥幼苗生長的同時,能夠誘導根內NO的過量產生[18]。但這些過量產生的NO在調控植物硒毒害反應中發揮何種作用尚不明確。硫化氫(Hydrogen sulfide,H2S)是最近被鑒定出的調控植物多種生理過程的一種全新氣體信號分子[19]。H2S可通過減少植物體內ROS的過量產生緩解植物的重金屬毒害反應[19]。另外,最近的研究結果顯示,H2S與NO的互作參與植物對鎘脅迫的響應和調控側根發育[20-21]。因此,明確硒通過調控H2S與NO互作的下游靶標將有助于進一步揭示硒毒害誘導植物產生過量ROS的分子機理。
小白菜屬十字花科蕓薹屬常見蔬菜,在實際生產中常作為富硒載體,但是外源添加無機硒易造成小白菜硒中毒。本試驗以小白菜幼苗根為材料,研究小白菜幼苗硒處理后根內源NO和H2S的變化情況,以及這種變化與小白菜體內2個Rboh基因(BrRbohG1和BrRbohG2)表達的相關性。以期為進一步闡明植物硒毒害的分子機理提供部分理論基礎。
1 材料與方法
1.1 材料與試劑
供試小白菜品種為南京綠領種業生產的五月慢。總NOS抑制劑L-NMMA、NO清除劑cPTIO、NO熒光染料DAF-DA均購自海門碧云天生物科技有限公司;NR抑制劑Na2WO4、H2S供體硫氫化鈉(NaHS)購自Sigma公司;TaKaRa Prime Script RT reagent購自TaKaRa公司;Trizol Reagent購自Invitrogen公司;H2S熒光染料WPS-1[3′-methoxy-3-oxo-3H-spiro(isobenzofuran-1,9′-xanthen)-6′-yl2-(pyridin-2-yldisulfanyl)benzoate],由美國華盛頓大學生化系Prof.Ming Xian惠贈。其余試劑均購自國藥集團。
1.2 試驗方法
1.2.1 幼苗培養和處理 種子經過1%NaClO消毒10 min后,用蒸餾水沖洗3次,在25℃條件下避光催芽12 h,然后轉入1/2 Hoagland營養液,在光照培養箱(RDN-300B-3)中培養,培養條件為25℃/22℃(晝/夜),光照周期14 h/10 h(光/暗),相對濕度75%。當幼苗根長至0.3 cm時開始處理。挑選生長健壯的幼苗到1/2 Hoagland營養液中。設置營養液中Na2SeO3最終濃度分別為:0 mmol/L、0.03mmol/L、0.06mmol/L、0.12 mmol/L、0.23 mmol/L、0.46 mmol/L,處理48 h時檢測根尖中內源NO、H2S;處理72 h后統計根長,每個處理10棵苗,3次重復,選取抑制率為50%左右的處理濃度作為后續試驗條件。為探討內源NO與內源H2S上下游關系設置以下處理: Na2SeO3(0 mmol/L)、Na2SeO3(0.06 mmol/L)、Na2SeO3+L-NMMA(200 μmol/L)、Na2SeO3+Tungstate(30 μmol/L)、Na2SeO3+cPTIO(100 μmol/L),處理48 h后檢測根尖中H2S含量。然后探討內源NO和內源H2S與BrRbohG1/BrRbohG2表達量關系,設計以下處理:Na2SeO3(0 mmol/L)、Na2SeO3(0.06mmol/L)、Na2SeO3+L-NMMA(200 μmol/L)、Na2SeO3+Tungstate(30μmol/L)、Na2SeO3+cPTIO(100 μmol/L)、Na2SeO3+NaHS (0.5 mmol/L)、NaHS(0.5 mmol/L)處理48 h后,取下根部保存于液氮中供后續試驗使用。
1.2.2 H2S組織熒光檢測與分析 根內H2S采用特異性熒光探針WSP-1進行原位檢測[21]。將不同處理的小白菜幼苗根部用蒸餾水沖洗3次,再吸干表面水分,將根系浸入到20 μmol/L WSP-1探針溶液中,在25℃、避光條件下裝載40 min。探針裝載完后用蒸餾水沖洗3次,通過在熒光顯微鏡(ECLIPSE,TE2000-S,Nikon)下以465 nm為激發波長、515 nm為發射波長,并結合使用Image-Pro Plus 6.0軟件統計根尖中WSP-1熒光密度,以此來反映H2S的相對含量。
1.2.3 NO組織熒光檢測與分析 根內NO采用特異性熒光探針DAF-DA進行原位檢測[22]。將不同處理的小白菜幼苗根部用蒸餾水沖洗3次,再吸干表面水分,將根系浸入到15 μmol/L DAF-DA探針溶液中,在25℃、避光條件下裝載15 min。探針裝載完后用蒸餾水沖洗3次,通過在熒光顯微鏡(ECLIPSE,TE2000-S,Nikon)下以490 nm為激發波長、525 nm為發射波長,并結合使用Image-Pro Plus 6.0軟件統計根尖中DAF-DA熒光密度,以此來反映NO的相對含量。
1.2.4BrRbohG1/BrRbohG2表達分析 首先以NCBI(http://www.ncbi.nlm.nih.gov/)中擬南芥的RbohG序列在小白菜基因組Brassica database(BRAD)(http://brassicadb.org/brad/index.php)中進行BLAST檢索,獲得相似性較高的小白菜BrRbohG類似基因。然后通過同源比對和序列結構分析進行確認。多重比對采用DNAMAN5.2. 2;蛋白質氨基酸序列結構分析采用SMART(http://smart. embl-heidelberg.de/)[23];蛋白質亞細胞定位預測采用WoLF PSORT(http://wolfpsort.org/)[24];啟動子調控區域分析采用PLACE(http://www.dna. affrc.go.jp/PLACE/signalscan.html)[25]。
基因表達分析采用RT-PCR。采用Trizol法提取根總RNA。使用TaKaRa PrimeSriptTMRT reagent試劑盒合成cDNA模板,保存于4℃條件下。分別以BrRbohG1(Bra019189)、BrRbohG2(Bra19191)、Actin序列設計特異性引物,引物序列分別為:BrRbohG1-F:5′-CATGACGGCTATCTGAGCGT-3′,BrRbohG1-R:5′-CGTGCTGCGATTTCGTTCAT-3′;BrRbohG2-F:5′-TTGGCTTGCGATTTTCCACG-3′,BrRbohG2-R:5′-GGATAAACCGCAGCGTTCAC-3′;Actin-F: 5′-GTGACAATGGAACTGGAATG-3′,Actin-R:5′-AGACGGAGGATAGCGTGAGG-3′。PCR反應體系:模板1 μl,前后引物各1 μl,PCR Mix 12 μl,超純水10 μl。PCR反應條件:94℃5 min;94℃30 s,BrRbohG1、BrRbohG2(50℃30 s),Actin(58℃30 s);72℃1 min,28個循環;72℃10 min。PCR產物用1%瓊脂糖凝膠電泳進行檢測,使用凝膠成像系統(Tanon-3500,上海天能)觀察拍照。
1.3 統計分析
每個結果采用3次重復的平均值±標準差表示,首先采用SPSS 2.0對試驗數據進行單因素方差分析(ANOVA),然后進行F檢驗檢測不同處理之間的差異顯著性。
2 結果與分析
2.1 Na2SeO3對小白菜幼苗根長的影響
Na2SeO3處理顯著抑制了小白菜根的生長(圖1)。在Na2SeO3濃度為0.06 mmol/L處理時平均根長約為對照組的36.9%,隨著Na2SeO3增加,根長逐漸變短,當Na2SeO3濃度為0.46 mmol/L時根長最短,抑制率達到93.0%,小白菜根部幾乎停滯生長,說明,在一定范圍內,Na2SeO3對小白菜幼苗根部生長有抑制作用,且抑制率與Na2SeO3濃度呈正相關。
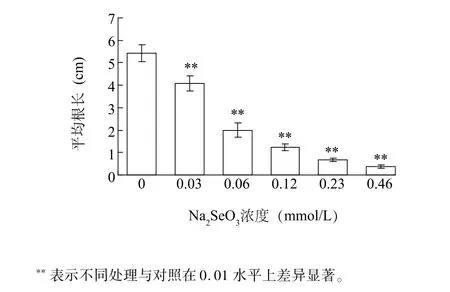
圖1 不同濃度Na2SeO3對小白菜根生長的影響Fig.1 Effects of Na2SeO3concentrations on the growth of Brassica rapa root
2.2 Na2SeO3對小白菜幼苗根尖內源NO和H2S含量的影響
圖2為DAF-DA熒光密度統計圖。隨著Na2SeO3濃度增加,小白菜根尖中DAF-DA熒光密度逐漸增加(圖2)。低濃度Na2SeO3(0.03 mmol/L)處理時,DAF-DA熒光密度與對照組差異不顯著;在0.06 mmol/L處理時DAF-DA熒光密度顯著高于對照組,比對照組增加42.9%。Na2SeO3濃度為0.12 mmol/L、0.23 mmol/L、0.46 mmol/L處理時DAF-DA熒光密度分別為對照組的2.18倍、2.34倍、2.89倍,與對照組均有極顯著差異。說明Na2SeO3處理下小白菜根尖內源NO含量增加,且隨著Na2SeO3濃度增加NO含量逐漸增加。
圖3為WSP-1熒光密度統計。隨著Na2SeO3濃度增加,WSP-1熒光密度逐漸降低(圖3)。0.03 mmol/L的Na2SeO3處理時,根尖WSP-1熒光密度與對照差異不顯著;當Na2SeO3為0.06 mmol/L時, WSP-1熒光密度顯著低于對照組,為對照的71.1%。當Na2SeO3濃度分別為0.12 mmol/L、0.23 mmol/L、0.46 mmol/L處理時,WSP-1熒光密度分別為對照組的57.5%、58.1%、39.4%,差異達到極顯著。由此發現Na2SeO3處理后小白菜根尖內源H2S含量下降,且隨著Na2SeO3濃度增加H2S含量逐漸下降。
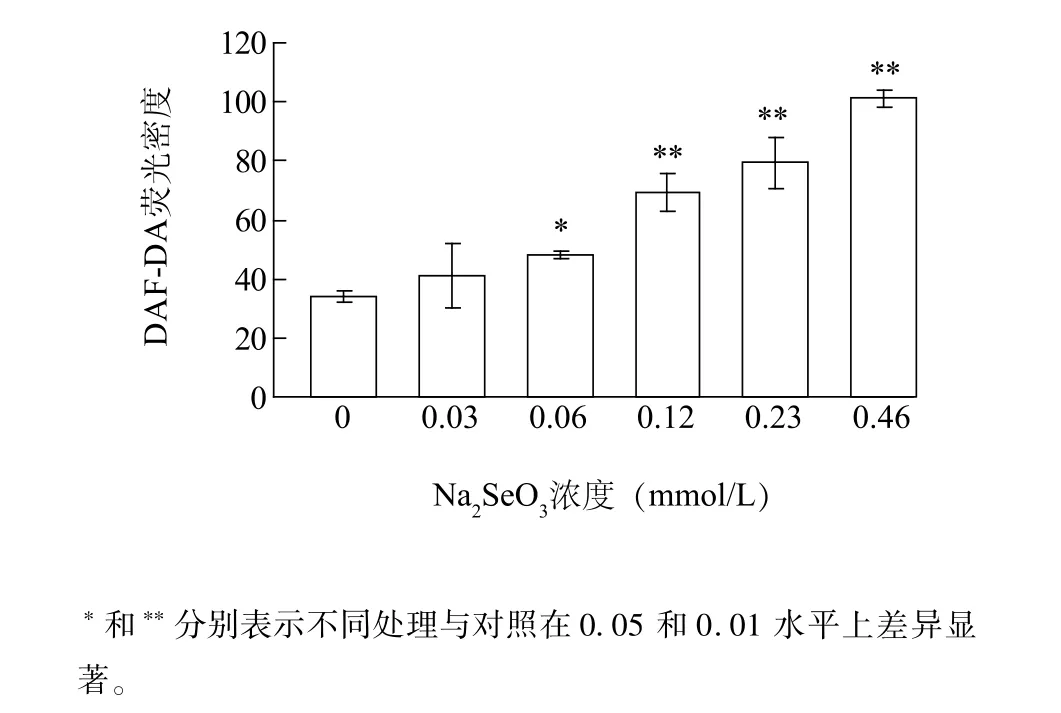
圖2 不同濃度Na2SeO3處理下小白菜幼苗根尖內源NO含量Fig.2 Effects of Na2SeO3concentrations on the content of endogenous NO in B.rapa root
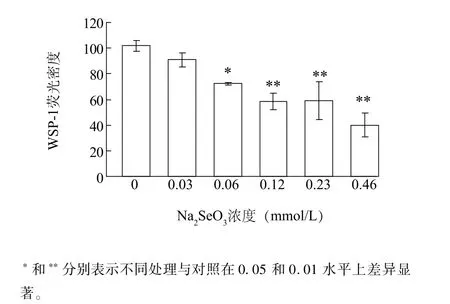
圖3 不同濃度Na2SeO3處理下小白菜根尖內源H2S含量Fig.3 Effects of Na2SeO3concentrations on the content of endogenous H2S in B.rapa root
2.3 Na2SeO3脅迫下小白菜根內源NO與H2S的互作
相關性分析結果顯示,不同濃度Na2SeO3處理過程中,根內源NO與H2S含量變化成顯著負相關,即:隨著NO含量的上升,H2S逐漸下降(圖4)。我們的前期研究結果顯示,H2S可作為NO的下游信號調控側根發育[21]。為了進一步探究NO與H2S在介導Na2SeO3對小白菜幼苗氧化損傷過程中的上下游關系,本試驗外源添加NO產生抑制劑(L-NMMA和Tungstate)與NO清除劑(cPTIO),再檢測根尖中內源H2S含量。由圖5可知, Na2SeO3處理后根尖WSP-1熒光密度與對照組相比顯著下降,為對照的43.7%;但是加入NO抑制劑(L-NMMA和Tungstate)和清除劑(cPTIO)后,根尖中WSP-1熒光密度分別為Na2SeO3處理組的1.43倍、1.37倍、1.78倍,差異均達到顯著水平。由此發現,通過抑制或清除根尖中NO,能夠增加根尖內源H2S含量,即外源硒處理條件下,根尖中內源NO抑制H2S產生。
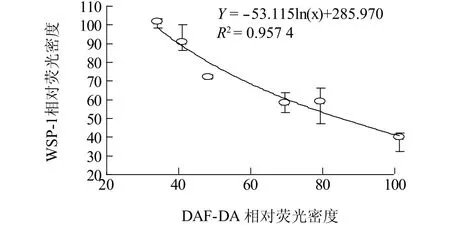
圖4 不同濃度Na2SeO3處理下小白菜根尖內源H2S含量(WSP-1熒光強度)與內源NO(DAF-DA熒光強度)相關性分析Fig.4 The correlation analysis between H2S content(WSP-1 fluorescent density)and NO content(DAF-DA fluorescent density)in B.rapa root under the stress of different concentration of Na2SeO3
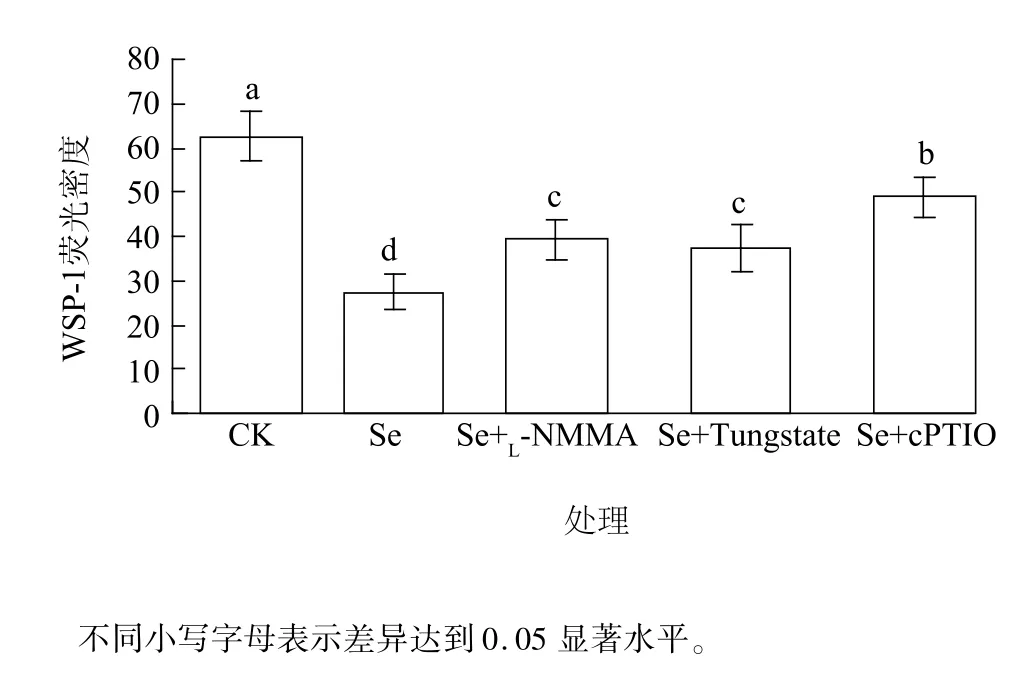
圖5 不同NO抑制劑和清除劑對小白菜幼苗根尖內源H2S的影響Fig.5 Effects of NO inhibitors and scavenger on the endogenous H2S in B.rapa root
2.4 Na2SeO3脅迫下小白菜根內源NO與H2S互作對BrRbohG1/BrRbohG2基因表達的影響
目前植物中報道的Rboh基因家族主要包含11個成員(RbohA-J)。相比較其他成員,RbohG的研究相對較少,其參與調控生理過程的報道也相對缺乏。因此,我們以擬南芥中的AtRbohG基因序列在小白菜基因組中進行BLAST,檢索到兩條同源序列:Bra019189(BrRbohG1)和Bra019191 (BrRbohG2)。亞細胞定位預測結果顯示,他們均定位于質膜或類囊體膜,屬于膜蛋白。根據已報道的植物氧化酶結構[26],我們進行了進一步的組成結構分析,結果表明:BrRbohG1和BrRbohG2中含有植物NADPH氧化酶典型的保守結構特征,如在EF-1和EF-2手型結構域、在EF-1上游N端方向的Ca2+互作結構域、Fe3+還原結構域、FAD結合域、NAD結合域(圖6)。多重比對結果顯示,上述特征性區域的關鍵作用位點在BrRbohG1、BrRbohG2和AtRbohG中相對保守一致(圖7)。
圖8顯示為不同處理對BrRbohG基因表達量的影響。與對照相比,經Na2SeO3處理的小白菜根系中BrRbohG1、BrRbohG2基因表達量均顯著上調,分別為對照的4.33倍和1.63倍。由于Na2SeO3處理能夠誘導內源NO的產生,因此我們在Na2SeO3處理時外源添加NO合成抑制劑L-NMMA、Tungstate,或NO清除劑cPTIO后,BrRbohG1和BrRbohG1的表達量分別比Na2SeO3單獨處理時下降了46.7%~61.7%和47.9%~83.6%。另外,由于Na2SeO3處理能夠抑制內源H2S的產生,因此在Na2SeO3處理時通過外源添加H2S供體NaHS來增加內源H2S含量。結果顯示,BrRbohG1和BrRbohG1的表達量分別比Na2SeO3單獨處理時下降了57.4%和78.8%。由此發現,在Na2SeO3處理條件下,減少根中的NO或增加H2S均能降低BrRbohG1和BrRbohG2的表達量。
2.5 BrRbohG1和BrRbohG2基因啟動子區域分析
在BRAD中定位BrRbohG1和BrRbohG2基因,下載起始密碼子上游2 000 bp序列并通過PLACE(http://www.dna.affrc.go.jp/PLACE/)進行啟動子區域預測分析。結果(表1)顯示,BrRbohG1和BrRbohG2的上游啟動子區域包含響應金屬離子調控元件MRE(TGCACAC)響應NO調控元件,如與bZIP轉錄因子結合的ACGT框(ACGT)等;響應生長素調控元件,如生長素響應因子ARFAT(TGTCTC))等。啟動子區域的不同響應元件說明在Na2SeO3脅迫下,BrRbohG1和BrRbohG2基因可能受金屬離子、NO、生長素等調控。
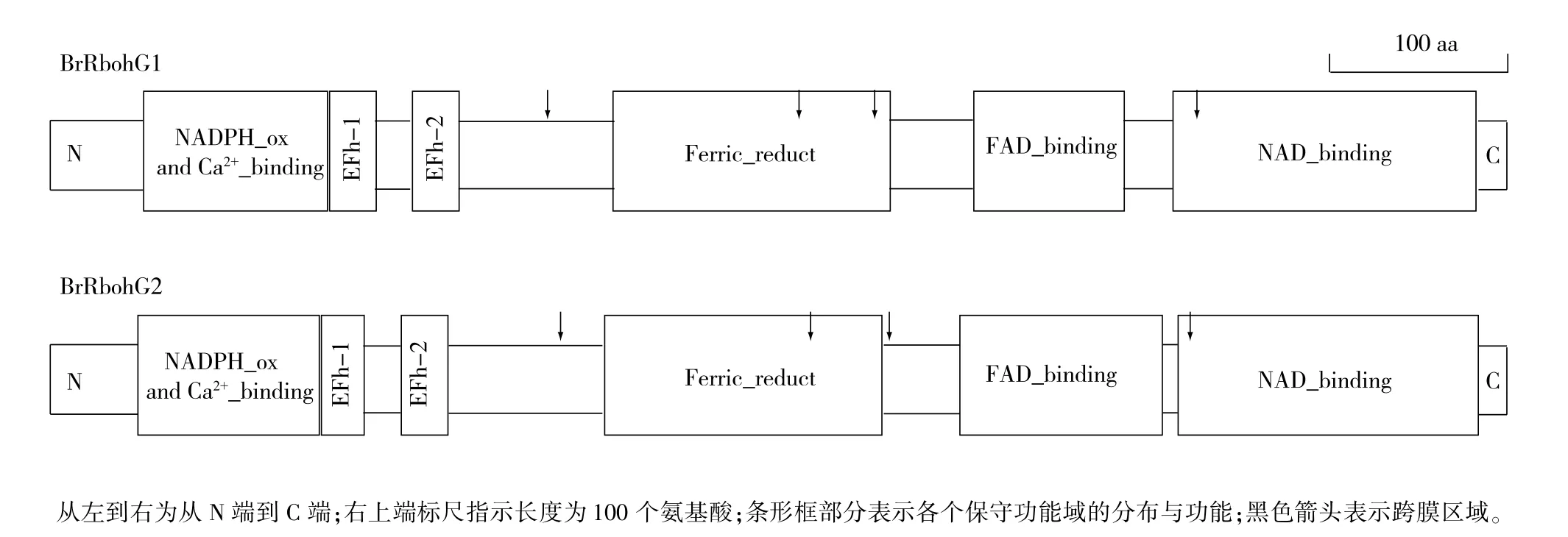
圖6 小白菜中BrRbohG1和BrRbohG2蛋白質序列保守域SMART分析Fig.6 SMART analysis for the conserved domains in BrRbohG1 and BrRbohG2 protein sequence

圖7 擬南芥中AtRbohG與小白菜中BrbohG1和BrRbohG2的多重比對分析Fig.7 The multi-alignment of AtRbohG in Arabdopsis and BrRbohG1 and BrRbohG2 in B.rapa
3 討論

圖8 抑制、清除NO或外源添加H2S對小白菜幼苗根部BrRbohG1和BrRbohG2表達量變化的影響Fig.8 Expression levels of BrRbohG1 and BrRbohG1 in B.rapa root treated with NO inhibitors,scavenger and H2S donor

表1 BrRbohG1與BrRbohG2基因上游啟動子區域不同信號響應元件Table 1 Signal responsive elements in the promoters of BrRbohG1 and BrRbohG2
目前關于硒在植物上的研究大多集中于硒在植物體受到外界脅迫時起到保護作用[27-28],而本試驗則關注在一定濃度時硒會導致植物生長受到抑制。植物遭受多數外界脅迫都會導致ROS積累,打破原有氧化與抗氧化系統的平衡[29],造成植物體細胞膜損傷、蛋白質合成受阻和含金屬的酶失活等,抑制植物生長。目前在擬南芥和水稻中分別發現10條和9條Rboh基因,它們都與ROS產生有關。前期試驗發現在小白菜中有12條具有NADPH氧化酶功能的基因,并發現在硒脅迫下根中BrRbohG1和BrRbohG2的表達量明顯上調。通過對BrRbohG1和BrRbohG2兩基因的啟動子區域分析得出二者啟動子區域包含NO、生長素和金屬離子響應元件,結合本試驗中硒抑制小白菜根生長并導致BrRbohG1和BrRbohG2基因表達上調,說明硒可能通過直接或間接的方式使小白菜中Rboh基因的表達量上調,繼而導致ROS積累。
NO和H2S是當下植物生理學的研究熱點。經過近20年的研究,發現NO幾乎在植物生長過程中的各個時期、部位都有調節作用[30]。NO在植物中有信號和毒害雙重作用,大部分文獻闡述了NO作為信號分子對植物受逆境脅迫時的保護作用。有文獻從NO與ROS互作方面解釋了NO存在的毒害作用[31]。在本試驗中發現硒處理后內源NO含量明顯上升,繼而通過一系列信號傳遞調節下游的Rboh基因最終導致植物受到脅迫作用,這一發現可能是內源NO對植物產生毒害的又一個新的途徑。相比于NO,H2S作為氣體信號分子在植物中的研究時間較短,通過外源添加或清除H2S證明了H2S參與了植物氣孔運動、光合作用、衰老以及對生物和非生物脅迫的調節作用[32-33]。對于H2S的研究也大都集中于其正面效應,在本試驗中外源硒導致小白菜根中內源H2S含量下降,當外源補充H2S時,BrRbohG1和BrRbohG2表達量相比硒處理有明顯下降,說明低濃度的H2S可能介導了硒誘導植物Rboh基因表達造成植物受到抑制。已有研究發現NO與H2S有上下游關系,Lisjak[34]發現H2S在調節擬南芥氣孔張開過程中使NO含量下降;而Li[35]發現NO能夠誘導下游H2S產生提高玉米幼苗耐熱性,說明在不同物種或不同處理條件下NO的上下游關系不是固定的。本試驗中降低內源NO含量能夠緩解Na2SeO3對H2S產生的抑制作用,且增加內源NO或降低內源H2S均能誘導BrRbohG1/G2表達上調,說明H2S的產生受上游NO含量調控,而H2S能夠直接或間接的調控BrRbohG1/BrRbohG2表達。
至于植物中NO與H2S間的相互調控機理還不是很清晰。研究發現在人體中發現NO和H2S能夠發生生物化學反應,形成RSNO類化合物[36];而且在大鼠中外源添加NO能夠改變CBS(Cystathionineβ-lyase)活性和H2S水平[37]。那么植物中NO與H2S也可能有類似的生化反應和調節機制,即過量的NO可能與H2S反應降低了內源H2S水平,也有可能抑制LCD或DCD基因表達,使內源H2S含量保持較低水平。因此NO與H2S在植物中的相互作用還需要更深入研究。
總之,本研究結果表明,Na2SeO3處理下,小白菜根尖產生過量的內源NO可能抑制下游內源H2S產生,且低濃度的H2S能夠誘導BrRbohG1和BrRbohG2的表達。
[1] ZHU Y G,PILON SMITS,ZHAO F J,et al.Selenium in higher plants:understanding mechanisms for biofortification and phytoremediation[J].Trends in Plant Science,2009,14(8):436-442.
[2] 方 勇,陳 曦,陳 悅,等.外源硒對水稻籽粒營養品質和重金屬含量的影響[J].江蘇農業學報,2013,29(4):760-765.
[3] 陳劍俠.葡萄葉片和果實對硒的吸收和累積特性[J].江蘇農業科學,2013,41(7):154-156.
[4] 吳麗軍,孫小風,張 榮,等.硒對不同品種春油菜含硒量、生物量及產量的影響[J].江蘇農業科學,2013,41(4):77-79.
[5] 張現偉,鄭家奎,張 濤,等.富硒水稻的研究意義與進展[J].雜交水稻,2009,24(2):5-9.
[6] DE T L,WAEGENEERS N,TACK F,et al.Selenium content of belgian cultivated siols and its uptake by field crops and vegetables [J].Science of the Total Environment,2014,468:77-82.
[7] HUANG S,HUA M,FENG J,et al.Assessment of selenium pollution in agricultural soils in the Xuzhou District,Northwest Jiangsu[J].China J Environ Sci,2009,21:481-487.
[8] NAVARRO-ALARCON M,CABRERA-VIQUE C.Selenium in food and the human body:a review[J].Sci Total Environ,2008,400:115-141.
[9] MACFARQUHAR J K,BROUSSARD D L,MELSTROM P,et al.Acute selenium toxicity associated with a dietary supplement [J].Archives of Internal Medicine,2010,170(3):256-261.
[10]MROCZEK-ZDYRSKA M,WóJCIK M.The influence of selenium on root growth and oxidative stress induced by lead inVicia faba L.minor plants[J].Biological Trace Element Research,2012,147 (1-3):320-328.
[11]李 秀,鞏 彪,徐 坤.外源NO對高溫脅迫下姜葉片活性氧代謝的影響[J].園藝學報,2014,41(2):277-284.
[12]魏志琴,陳志勇,秦 蓉,等.Cu2+對擬南芥根的局部毒性及誘導DNA損傷和細胞死亡[J].植物學報,2013,48(3):303-312.
[13]伏 毅,戴 媛,譚曉榮,等.干旱對小麥幼苗脂類和蛋白質氧化損傷的影響[J].作物雜志,2010(3):45-50.
[14]BESSON-BARD A,PUGIN A,WENDEHENNE D.New insights into nitric oxide signaling in plants[J].Annual Review of Plant Biology,2008,59:21-39.
[15]GALLEGO S M,PENA L B,BARCIA R A,et al.Unravelling cadmium toxicity and tolerance in plants:Insight into regulatory mechanisms[J].Environ Exp Bot,2012,83:33-46.
[16]BESSON-BARD A,GRAVOT A,RICHAUD P,et al.Nitric oxide contributes to cadmium toxicity inArabidopsisby promoting cadmium accumulation in roots and by up-regulating genes related to iron uptake[J].Plant Physiol,2009,149:1302-1315.
[17]CHEN M,CUI W,ZHU K,et al.Hydrogen-rich water alleviates aluminum-induced inhibition of root elongation in alfalfa via decreasing nitric oxide production[J].J Hazard Mater,2014,267: 40-47.
[18]LEHOTAI N,PET? A,ERDEI L,et al.The effect of selenium (Se)on development and nitric oxide levels inArabidopsis thalianaseedlings[J].Acta Biologica Szegediensis,2011,55:105-107.
[19]LISJAK M,TEKLIC T,WILSON I D,et al.Hydrogen sulfide: environmental factor or signalling molecule?[J].Plant Cell Environ,2013,36:1607-1616.
[20]SHI H,YE T,CHAN Z.Nitric oxide-activated hydrogen sulfide is essential for cadmium stress response in bermudagrass[Cynodon dactylon(L.)Pers.][J].Plant Physiology and Biochemistry. 2014,74:99-107.
[21]LI Y J,CHEN J,XIAN M,et al.In site bioimaging of hydrogen sulfide uncovers its pivotal role in regulating nitric oxide-induced lateral root formation[J].PLoS ONE,2014,9:e90340.
[22]GUO K,XIA K,YANG Z M.Regulation of tomato lateral root development by carbon monoxide and involvement in auxin and nitric oxide[J].Journal of Experimental Botany,2008,59:3443-3452.
[23]LETUNIC I,DOERKS T,BORK P.SMART 7:recent updates to the protein domain annotation resource[J].Nucleic Acids Res, 2012,40:302-305.
[24]HORTON P,PARK K J,OBAYASHI T,et al.WoLF PSORT: protein localization predictor[J].Nucleic Acids Res,2007,35: 585-587.
[25]HIGO K,UGAWA Y,IWAMOTO M,et al.Plant cis-acting regulatory DNA elements(PLACE)database[J].Nucleic Acids Res, 1999,27:297-300.
[26]D MAARINO,C DUNAND,A PUPPO,et al.A burst of plant NADPH oxidase[J].Trends in Plant Science,2012,17(1):9-15.
[27]SOLEIMANZADEH H.Response of sunflower(Helianthus annuusL.)to selenium application under water stress[J].World Applied Sciences Journal,2012,17(9):1115-1119.
[28]DJANAGUIRAMAN M,PRASAD P,SEPPANEN M.Selenium protects sorghum leaves from oxidative damage under high temperature stress by enhancing antioxidant defense system[J].Plant Physiology and Biochemistry,2010,48(12):999-1007.
[29]王東明,賈 媛,崔繼哲.鹽脅迫對植物的影響及植物鹽適應性研究進展[J].中國農學通報,2009,25(4):124-128.
[30]史慶華,賴齊賢,朱祝軍,等.一氧化氮在植物中的生理功能[J].細胞生物學雜志,2005,27(1):39-42.
[31]WENDEHENNE D,PUGIN A,KLESSIG D F,et al.Nitric oxide:comparative synthesis and signaling in animal and plant cells [J].Trends in Plant Science,2001,6(4):177-183.
[32]崔為體,沈文飚.植物中硫化氫的生理功能及其分子機理[J].生命的化學,2012,32(4):385-389.
[33]LI L,WANG Y,SHEN W.Roles of hydrogen sulfide and nitric oxide in the alleviation of cadmium-induced oxidative damage in alfalfa seedling roots[J].Biometals,2012,25(3):617-631.
[34]LISJAK M,SRIVASTAVA N,TEKLIC T,et al.A novel hydrogen sulfide donor causes stomatal opening and reduces nitric oxide accumulation[J].Plant Physiology and Bochemistry,2010,48 (12):931-935.
[35]LI Z G,YANG S Z,LONG W B,et al.Hydrogen sulphide may be a novel downstream signal molecule in nitric oxide-induced heat tolerance of maize(Zea maysL.)seedlings[J].Plant,Cell& Environment,2013,36(8):1564-1572.
[36]WHITEMAN M,LI L,KOSTETSKI I,et al.Evidence for the formation of a novel nitrosothiol from the gaseous mediators nitric oxide and hydrogen sulphide[J].Biochemical and Biophysical Research Communications,2006,343(1):303-310.
[37]MOK Y Y P,ATAN M,BIN M S,et al.Role of hydrogen sulphide in haemorrhagic shock in the rat:protective effect of inhibitors of hydrogen sulphide biosynthesis[J].British Journal of Pharmacology,2004,143(7):881-889.
(責任編輯:陳海霞)
Selenium-induced expression of BrRbohG1/BrRbohG2 in Brassica rapa root through nitric oxide and hydrogen sulfide signalling
CHEN Yi1,2, CHEN Jian2, Lü Wen-jing1,2, LI Li-na1,2, YANG Li-fei1
(1.College of Horticulture,Nanjing Agricultural University,Nanjing 210095,China;2.Institute of Food Quality Safety and Detection,Jiangsu Academy of Agricultural Sciences,Nanjing 210014,China)
Selenium(Se)has been becoming an emergent pollutant in the agricultural environment.In order to explore the mechanism of Se-induced stress responses in crops,the present study investigated the effect of Na2SeO3treatment on two important endogenous signalling molecules,hydrogen sulfide(H2S)and nitric oxide(NO),on the roots ofBrassica rapaand the regulation of the expression ofBrRbohG1andBrRbohG2by H2S and NO.Results were achieved as follows.The root growth was significantly inhibited by Na2SeO3varying from 0.03 mmol/L to 0.46 mmol/L in a dose-dependent manner.Compared to the control group,the root elongation was decreased by almost 60%in the treatment of Na2SeO3at 0.06 mmol/L.Treatment with Na2SeO3resulted in the increase in endogenous NO content and the decrease in endogenous H2S content in root in dose-dependent manners.The correlation analysis suggested that there was asignificantly negative correlation between NO and H2S in root with Na2SeO3treatment.The inhibition of H2S induced by 0.06 mmol/L Na2SeO3could be significantly attenuated by the addition of both NO synthesized inhibitors (L-NMMA and Tungstate)and NO scavenger(cPTIO).The expressions ofBrRbohG1andBrRbohG2could be induced significantly by Na2SeO3at 0.06 mmol/L,which could be inhibited by the addition ofL-NMMA,Tungstate, cPTIO,and H2S donor sodium hydrosulfide(NaHS),respectively.In conclusion,both endogenous NO and H2S are probably involved in the regulation of the expressions ofBrRbohG1andBrRbohG2in the root ofB.rapaunder Na2SeO3stress.The endogenous NO mediated Na2SeO3-induced expressions ofBrRbohG1andBrRbohG2while the endogenous H2S might inhibit the expressions ofBrRbohG1andBrRbohG2in the roots ofB.rapaunder Na2SeO3stress. And NO might have functioned the upstream of H2S in the above process.
Brassica rapa;selenium;hydrogen sulfide;nitric oxide;BrRbohG1/BrRbohG2
S634.3
A
1000-4440(2015)01-0149-09
陳 義,陳 健,呂文靜,等.硒通過調控小白菜幼苗根內NO與H2S信號誘導BrRbohG1/BrRbohG2表達[J].江蘇農業學報, 2015,31(1):149-157.
10.3969/j.issn.1000-4440.2015.01.024
2014-08-14
國家自然科學基金項目(31401857);江蘇省農業科技自主創新基金項目[CX(13)5052]
陳 義(1990-),男,安徽六安人,碩士研究生,研究方向為蔬菜生理與生物技術。(E-mail)2012104082@njau.edu. cn
楊立飛,(E-mail)lfy@njau.edu.cn
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
