科爾沁草甸生態(tài)系統(tǒng)凈碳交換特征及其驅(qū)動(dòng)因子
2015-06-26 11:00:43劉廷璽雷慧閩張圣微段利民
草業(yè)學(xué)報(bào) 2015年11期
王 婧,劉廷璽*,雷慧閩,張圣微,段利民
(1.內(nèi)蒙古農(nóng)業(yè)大學(xué)水利與土木建筑工程學(xué)院,內(nèi)蒙古呼和浩特010018;2.清華大學(xué)水利水電工程系,北京100084)
科爾沁草甸生態(tài)系統(tǒng)凈碳交換特征及其驅(qū)動(dòng)因子
王 婧1,劉廷璽1*,雷慧閩2*,張圣微1,段利民1
(1.內(nèi)蒙古農(nóng)業(yè)大學(xué)水利與土木建筑工程學(xué)院,內(nèi)蒙古呼和浩特010018;2.清華大學(xué)水利水電工程系,北京100084)
以2013年10月-2014年9月連續(xù)觀測(cè)的CO2通量數(shù)據(jù)為基礎(chǔ),分析了科爾沁草甸生態(tài)系統(tǒng)凈碳交換量(NEE)的時(shí)間變化特征及其驅(qū)動(dòng)因素。結(jié)果表明,NEE日變化季節(jié)差異明顯,生長(zhǎng)季變化幅度大,凈CO2日吸收速率7月>8月>9月>6月>5月;生長(zhǎng)季內(nèi),NEE主要受控于葉面積指數(shù)和光合有效輻射。NEE與光合有效輻射(PAR)之間的關(guān)系可用直角雙曲線方程來(lái)描述,擬合得到的表觀初始光能利用率α為0.0015μmol CO2/μmol PAR,最大光合速率Pmax為0.65μmol CO2/(m2·s)。葉面積指數(shù)(LAI)對(duì)NEE的影響可由分段函數(shù)表示,當(dāng)LAI>3.08時(shí),表現(xiàn)為漸進(jìn)飽和型,且LAI越大NEE對(duì)PAR的響應(yīng)越明顯;當(dāng)高飽和水汽壓差(VPD)在1.5~2.0 kPa時(shí),光合作用開(kāi)始降低,NEE明顯受到VPD值的抑制;短暫強(qiáng)降雨(累計(jì)降雨量>40 mm/d)對(duì)晝間NEE有一定的抑制,而持續(xù)低強(qiáng)度降雨(降雨時(shí)長(zhǎng)>15 h)對(duì)夜間NEE存在激發(fā)作用;夜間NEE隨土壤溫度呈指數(shù)增長(zhǎng),溫度敏感系數(shù)(Q10)為2.63。
凈生態(tài)系統(tǒng)交換量;驅(qū)動(dòng)因子;渦度相關(guān)系統(tǒng);科爾沁沙地
陸地生態(tài)系統(tǒng)的碳循環(huán)及其對(duì)環(huán)境因子的響應(yīng)研究是理解生態(tài)系統(tǒng)碳平衡的關(guān)鍵。草地生態(tài)系統(tǒng)作為陸地生態(tài)系統(tǒng)的重要組成部分,幾乎覆蓋了陸地面積的20%,其凈初級(jí)生產(chǎn)力約占全球陸地生物區(qū)凈初級(jí)生產(chǎn)力的1/3。美歐科學(xué)家于1996年初步建立起全球CO2通量網(wǎng),利用渦度技術(shù)在全球范圍內(nèi)開(kāi)展了不同緯度、不同類(lèi)型生態(tài)系統(tǒng)CO2通量的全球性研究,但其中70%是針對(duì)不同森林植被類(lèi)型的,對(duì)草原生態(tài)系統(tǒng)的研究主要集中在低海拔的平原地區(qū)[1-2],且研究較少。Dugas等[2]對(duì)德高草原碳收支狀況研究發(fā)現(xiàn)葉面積指數(shù)(LAI)、光合有效輻射(PAR)及土壤含水量影響碳通量大小,Shaver等[3]應(yīng)用Pan-Arctic模型對(duì)生態(tài)系統(tǒng)凈生態(tài)系統(tǒng)碳交換(以下簡(jiǎn)稱(chēng)“NEE”)研究表明,NEE的變異約75%可由LAI、氣溫(Ta)、PAR函數(shù)的單一回歸模型解釋。我國(guó)自20世紀(jì)90年代開(kāi)始應(yīng)用渦度相關(guān)法開(kāi)展了陸面過(guò)程中碳循環(huán)通量的研究,盡管起步稍晚,但也取得了部分有價(jià)值的研究成果。伏玉玲等[4]研究了中國(guó)草地生態(tài)系統(tǒng)碳收支及其對(duì)環(huán)境的響應(yīng),郝彥賓等[5]研究了內(nèi)蒙古羊草(Leymus chinensis)草原碳交換的季節(jié)變異規(guī)律,發(fā)現(xiàn)水分和PAR是影響碳交換的兩個(gè)重要生態(tài)因子。Zhao等[6]研究了青藏高寒草甸凈碳交換特征,表明葉面積指數(shù)(GLAI)、土壤含水量(θ)、Ta是3個(gè)對(duì)NEE季節(jié)變異影響最大的指標(biāo)。
科爾沁沙地草甸區(qū)作為我國(guó)北方典型的半干旱草原區(qū),具有極其脆弱的草原生態(tài)資源。與其他下墊面相比,對(duì)氣候變化的響應(yīng)更為敏感。近年來(lái),有關(guān)該地區(qū)的碳通量研究主要局限于短期、不連續(xù)的觀測(cè)數(shù)據(jù)[7]及碳排放的觀測(cè)[8],針對(duì)NEE的研究比較缺乏。而研究半干旱區(qū)草甸生態(tài)系統(tǒng)NEE的日、季變化及其與氣象因子的關(guān)系,對(duì)于了解碳的源匯過(guò)程、開(kāi)展草原碳循環(huán)的模擬和氣候變化情景下的預(yù)測(cè)研究具有十分重要的意義。本文依據(jù)渦度相關(guān)系統(tǒng)2013年10月-2014年9月的連續(xù)觀測(cè)數(shù)據(jù),對(duì)科爾沁草甸生態(tài)系統(tǒng)的碳收支及其過(guò)程機(jī)制進(jìn)行定量評(píng)價(jià),分析各氣象要素對(duì)碳交換的影響,揭示草甸草原NEE的日、季變化規(guī)律,解析主要環(huán)境因子對(duì)碳源匯的影響及其差異,為評(píng)估該地區(qū)碳收支和明晰全球碳循環(huán)提供更為完善的試驗(yàn)數(shù)據(jù)和理論基礎(chǔ)。
1 材料與方法
1.1 研究區(qū)概況
試驗(yàn)研究區(qū)隸屬內(nèi)蒙古通遼市科爾沁左翼后旗,面積約55 km2。該區(qū)多年平均氣溫約6.6℃,1月平均氣溫最低,為-13.3℃,7月平均氣溫最高,為23.8℃;多年平均降水量389 mm,降水主要集中在6-8月,占年總降水量的69.3%;多年平均蒸發(fā)量1412 mm,蒸發(fā)主要集中在4-9月。此間蒸發(fā)量占全年的17%。試驗(yàn)區(qū)為沙丘-草甸相間分布地區(qū),草甸面積約占整個(gè)研究區(qū)面積的26.6%,本文選取草甸試驗(yàn)站(122°39′15″E,43°20′14″N)作為研究對(duì)象,該試驗(yàn)站地面相對(duì)高程約188.5 m,南臨小型湖泊,地勢(shì)平坦。天然植被主要有羊草、蘆葦(Phragmites australis)等。6-8月份植被生長(zhǎng)旺季,植被平均高度為145 cm,平均葉面積指數(shù)為2.47。
1.2 觀測(cè)儀器與方法
草甸試驗(yàn)點(diǎn)主要安裝有渦度相關(guān)系統(tǒng)傳感器、微氣象觀測(cè)系統(tǒng)。分別在5,10,20,40,80,160 cm處埋有6層土壤溫度、土壤含水量觀測(cè)傳感器。每30 d進(jìn)行一次數(shù)據(jù)采集與處理,渦度原始數(shù)據(jù)與氣象數(shù)據(jù)同步下載,本研究主要以2013年10月-2014年9月連續(xù)觀測(cè)的CO2通量數(shù)據(jù)及微氣象數(shù)據(jù)為基礎(chǔ)。
1.2.1 渦度相關(guān)系統(tǒng)和微氣象系統(tǒng)觀測(cè) 渦度相關(guān)系統(tǒng)傳感器安裝在距地面2.6 m處,主要用于觀測(cè)湍流通量。該系統(tǒng)主要包括三維超聲風(fēng)速儀(CSAT-3,Campbell Scientific,USA)及紅外氣體分析儀(LI-7500,Li-COR Inc,Nebraska,USA),分別用于測(cè)定三維風(fēng)速與大氣CO2和H2O密度,由數(shù)據(jù)采集器(CR3000,Campbell Scientific,USA)記錄并存儲(chǔ)系統(tǒng)同步計(jì)算的二氧化碳和水熱通量,采樣頻率為10 Hz,數(shù)據(jù)記錄時(shí)間間隔為30 min。
微氣象觀測(cè)系統(tǒng)主要包括安裝在渦度觀測(cè)塔上的四分量?jī)糨椛鋬x(NR-LITE,Campbell Scientific,USA)、光合有效輻射儀(LI190SB,Campbell Scientific,USA),以及3層(1.5,2.4,10.0 m)溫濕度測(cè)量?jī)x器(HMP45C,Campbell Scientific,US)、土壤熱通量板(HFP01,Campbell Scientific,USA)、120 cm口徑水面蒸發(fā)量(255,Nova Lynx,US)、降水量(TE525MM,Campbell Scientific,US)等。這些監(jiān)測(cè)要素由數(shù)據(jù)采集器(CR1000,Campbell Scientific,USA)按10 min時(shí)間間隔計(jì)算其平均值并存儲(chǔ)。
1.2.2 葉面積指數(shù)觀測(cè) 對(duì)草甸群落葉面積指數(shù)LAI的動(dòng)態(tài)測(cè)定也與碳通量觀測(cè)同步進(jìn)行,利用植物冠層分析儀(LAI-2200,Li-COR Nebraska,USA)通過(guò)測(cè)定冠層上方(簡(jiǎn)稱(chēng)“A”值)、冠層下方(簡(jiǎn)稱(chēng)“B”值)光照輻射的衰減來(lái)計(jì)算葉面積指數(shù)(葉面積指數(shù)=葉面積/地面積)。草甸植被生長(zhǎng)初期的4-5月份,每隔15~20 d測(cè)量一次,6-9月份每20~30 d測(cè)量一次。選取3個(gè)1 m×1 m的樣方,每個(gè)樣方測(cè)定1個(gè)A值,4個(gè)B值,采用90度遮蓋帽,取3組葉面積指數(shù)的平均值作為代表值。
1.3 數(shù)據(jù)處理
1.3.1 CO2通量的計(jì)算公式 CO2通量即凈生態(tài)系統(tǒng)碳交換量,由CO2(μmol/m2·s)/H2O(mmol/m2·s)的濃度與垂直風(fēng)速的原始數(shù)據(jù)經(jīng)過(guò)協(xié)方差計(jì)算來(lái)確定。式中,w'為垂直湍流速度脈動(dòng),c'為二氧化碳濃度。NEE由渦度相關(guān)系統(tǒng)測(cè)定的CO2通量直接獲得,NEE為正表示生態(tài)系統(tǒng)向大氣釋放CO2,反之則表示從大氣吸收CO2。

1.3.2 通量數(shù)據(jù)的質(zhì)量控制與插補(bǔ) 對(duì)渦動(dòng)相關(guān)法觀測(cè)到的10 Hz原始湍流數(shù)據(jù)運(yùn)用Logger Net軟件進(jìn)行格式轉(zhuǎn)換(TOB3-TOB1),然后采用美國(guó)LI-COR公司研發(fā)的Eddy Pro軟件[9]進(jìn)行系列校正計(jì)算,包括異常值及野值點(diǎn)剔除、坐標(biāo)旋轉(zhuǎn)修正、頻率損失修正、超聲虛溫修正、空氣密度效應(yīng)修正(WPL修正)等基本處理,得到采樣間隔為30 min的通量數(shù)據(jù)。
對(duì)30 min數(shù)據(jù),若野點(diǎn)數(shù)大于100,則剔除該時(shí)次數(shù)據(jù);或根據(jù)LI-7500紅外氣體分析儀診斷異常標(biāo)志值(diag≠0)[10],以及相關(guān)監(jiān)測(cè)數(shù)據(jù)超出CO2濃度閾值(100~1000 mg/m3),對(duì)夜間觀測(cè),用給定的一個(gè)臨界閾值uc*加以檢驗(yàn)。當(dāng)u*<uc*時(shí),則舍棄該通量數(shù)據(jù)不進(jìn)行分析[11]。uc*值一般取0.1~0.2 m/s,本文取0.1 m/s。
對(duì)原始通量數(shù)據(jù)進(jìn)行質(zhì)量控制后,剔除的低質(zhì)量數(shù)據(jù)會(huì)影響數(shù)據(jù)的連續(xù)性。因此,本研究對(duì)夜間和晝間缺失的NEE數(shù)據(jù),采用分別的方法進(jìn)行插補(bǔ)。針對(duì)夜間缺失的NEE數(shù)據(jù),利用5 cm深度處的土壤溫度(Ts)采用Vants Hoff擬合方程加以插補(bǔ)[12]。

式中,Reco為夜間生態(tài)系統(tǒng)呼吸速率(由于夜間沒(méi)有光合作用,Reco即為NEE;Reco,ref為生態(tài)系統(tǒng)10℃時(shí)的參考呼吸速率(本文取0.05 mg/m2·s);Q10為呼吸隨溫度的變化速率(本文取2.63),即5至40℃之間,溫度每增加10℃單位時(shí)間土壤呼吸增加的倍數(shù),Raich和Schlesinger[13]通過(guò)收集和分析全球各種生態(tài)系統(tǒng)的土壤呼吸數(shù)據(jù),發(fā)現(xiàn)Q10在1.3~3.3之間波動(dòng),Ts為5 cm深度處土壤實(shí)測(cè)溫度。
針對(duì)晝間缺失的NEE數(shù)據(jù),則利用Michaelis-Menten函數(shù)進(jìn)行插補(bǔ)[13]。

式中,α為生態(tài)系統(tǒng)表觀光量子效率;pmax為生態(tài)系統(tǒng)最大光合速率,即PAR→∞時(shí)NEE的漸進(jìn)值;Reco,d為暗呼吸速率,即PAR→0時(shí)的NEE值。
2 結(jié)果與分析
2.1 凈二氧化碳通量日內(nèi)變化特征
草甸NEE的日變化基本呈現(xiàn)單峰型(圖1),且呈現(xiàn)出上午碳吸收較強(qiáng),下午碳吸收較弱的不對(duì)稱(chēng)現(xiàn)象,在日出(5:00-6:00時(shí))、日落(18:00-19:00時(shí))前后碳通量波動(dòng)較大,此時(shí)的呼吸作用與光合作用達(dá)到一個(gè)平衡狀態(tài)。夜間NEE為正,整個(gè)生態(tài)系統(tǒng)表現(xiàn)為碳排放,白天NEE均為負(fù)值,即隨著光合作用的加強(qiáng),整個(gè)生態(tài)系統(tǒng)表現(xiàn)為凈碳吸收,5-9月日均碳吸收速率分別為0.02,0.12,0.33,0.29,0.24 mg/(m2·s),呈現(xiàn)為7月>8月>9月>6月>5月,且碳吸收出現(xiàn)在凌晨的時(shí)間依次滯后0.25,0.25,0.50,0.50 h,碳排放出現(xiàn)在傍晚的時(shí)間依次提前0.5,0.5,1.0,1.5 h。同時(shí),日出、日落前后碳吸收量與排放量達(dá)到相對(duì)平衡的狀態(tài)。

圖1 生長(zhǎng)季草甸生態(tài)系統(tǒng)碳通量月平均日變化Fig.1 Diurnal changes of CO2fluxes during growing season over the meadow grassland ecosystem
2.2 凈二氧化碳通量及其影響因子的季節(jié)變化特征
為了解NEE與環(huán)境因子的季節(jié)變化特征,本文選取2013年10月-2014年9月生態(tài)系統(tǒng)NEE及其主要環(huán)境因子(飽和水汽壓差VPD、氣溫Ta、土壤溫度Ts、光合有效輻射PAR)的月變化加以分析(圖2)。氣溫與5 cm深度處的土壤溫度最大值均出現(xiàn)在7月中旬,分別為29.3和26.6℃,草甸生長(zhǎng)季節(jié)的氣溫、晝夜溫差、土壤溫度的日平均值分別為19.3,10.0,16.0℃;VPD的日平均值為1.08 kPa,最高、最低值分別出現(xiàn)在7月9日和10月18日;PAR的月平均日變化呈現(xiàn)“單峰型”,最大值出現(xiàn)在7月5日,為1874μmol/(m2·s);8月降水量最高,為245.9 mm,7月次之,為104.6 mm。生長(zhǎng)季平均葉面積指數(shù)LAI為2.57。4-6月LAI迅速增加,6-8月增速放緩,8月中下旬達(dá)到最大值3.34,9月微弱下降,但降幅較小。
NEE在全年表現(xiàn)出“兩峰一谷”的特征(圖2d),其中“峰值”出現(xiàn)在1,10月,均值分別為0.13和0.08 mg/(m2·s),“谷值”出現(xiàn)在7月,均值-0.33 mg/(m2·s),較徐麗君等[14]研究的貝加爾針茅草(Stipa baicalensis)原碳吸收谷值偏小。4月草甸NEE均值開(kāi)始由正轉(zhuǎn)負(fù),生長(zhǎng)季的5-9月,生態(tài)系統(tǒng)主要表現(xiàn)為碳匯,凈CO2吸收總量1.02 mg/(m2·s),5,6,7,8,9月分別貢獻(xiàn)了11.2%,27.0%,30.5%,22.5%,8.4%。但不同緯度的草甸生態(tài)系統(tǒng)NEE正負(fù)轉(zhuǎn)換的時(shí)間有較大差異,如Shi等[15]研究嵩草草甸生態(tài)系統(tǒng)7-9月為碳吸收,其余為碳排放。而本研究區(qū)從10月出現(xiàn)碳排放,10月至翌年3月,凈CO2日排放均值0.11 mg/(m2·s)。同時(shí)生態(tài)系統(tǒng)碳交換因受到環(huán)境因素影響而產(chǎn)生差異(表1),如生長(zhǎng)季NEE與PAR、LAI相關(guān)性顯著(P<0.01),非生長(zhǎng)季NEE主要受控于溫度因素(R2=0.568),生態(tài)系統(tǒng)主要表現(xiàn)為碳源。研究時(shí)段,草甸作為碳匯從大氣中吸收218.52 g CO2/m2。
2.3 葉面積指數(shù)對(duì)NEE的影響
LAI是反映植物長(zhǎng)勢(shì)的重要特征參數(shù),它對(duì)植物光能利用、干物質(zhì)積累等都有明顯影響。生長(zhǎng)初期(LAI<0.97),由于植被稀疏LAI較小,對(duì)散射光的吸收較少,草甸CO2吸收量隨LAI的增加而線性增長(zhǎng);生長(zhǎng)旺期(0.97<LAI<3.08)植被生長(zhǎng)稠密,LAI增加冠層有利于植物吸收來(lái)自各個(gè)方向的散射光,CO2吸收量隨LAI的增加而緩慢增加,之后漸進(jìn)飽和狀態(tài);生長(zhǎng)末期(LAI>3.08)部分葉片開(kāi)始枯黃、凋落,LAI有所下降,碳吸收減少且隨LAI的增加而趨于平穩(wěn)(圖3),這與Suyker等[16]對(duì)玉米(Zea may)生殖生長(zhǎng)研究得出的結(jié)論相似。同時(shí)由于生長(zhǎng)末期群體底層存在大量枯葉,利用冠層分析儀進(jìn)行測(cè)量時(shí),儀器無(wú)法自動(dòng)消除這部分枯葉的影響,導(dǎo)致LAI測(cè)定結(jié)果偏大[17]。
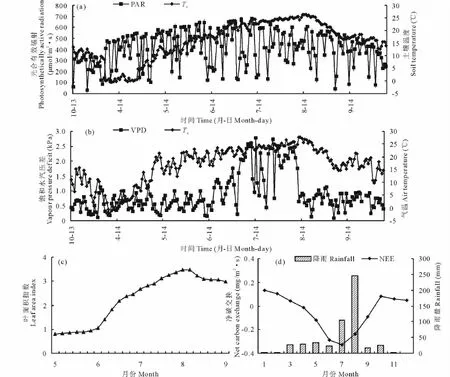
圖2 凈碳交換量NEE與環(huán)境因子的季節(jié)變化Fig.2 Seasonal variations of environmental factors with net carbon exchange(NEE)

表1 凈碳交換NEE與氣象因子標(biāo)準(zhǔn)化回歸分析Table 1 Normalized regression analysis between net ecosystem CO2exchange(NEE)and meteorology factors
生長(zhǎng)季LAI與PAR的偏相關(guān)分析(表2)表明,當(dāng)控制PAR時(shí),LAI與NEE相關(guān)性不顯著(R2=0.38,P>0.05),而考慮PAR影響時(shí),LAI與NEE相關(guān)系數(shù)提高了40.6%,二者顯著相關(guān)(P<0.01),LAI與PAR存在明顯的交互作用,共同影響著生態(tài)系統(tǒng)凈碳交換過(guò)程。
2.4 光合有效輻射對(duì)NEE的影響
生長(zhǎng)季的NEE與PAR變化關(guān)系較為分散,通常用一個(gè)直角雙曲線模型表示(公式3)。其中表觀初始光能效率α,利用低光強(qiáng)下NEE-PAR的線性擬合關(guān)系公式反推得到[18](NEE=0.0015PAR+0.264,R2=0.924),即α值為0.0015μmol CO2/μmol PAR。由NEE-PAR的擬合曲線(圖4a)可以估計(jì)光補(bǔ)償點(diǎn)的范圍變化在80~150μmol/(m2·s)之間。
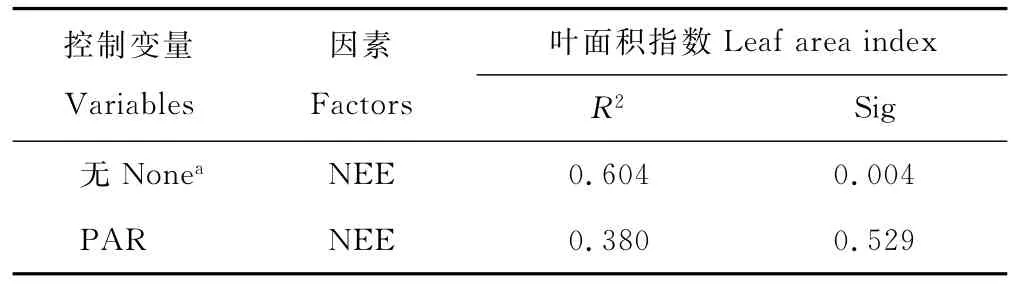
表2 LAI與PAR偏相關(guān)分析Table 2 Partial correlation analysis between LAI and PAR

圖4 凈碳交換NEE與光合有效輻射PAR的關(guān)系Fig.4 Relationships between net ecosystem CO2exchange(NEE)and photosynthetically active radiation(PAR)in growing season
草甸生態(tài)系統(tǒng)NEE對(duì)光強(qiáng)PAR的響應(yīng)分為3個(gè)不同階段(圖4b):PAR<300μmol/(m2·s)時(shí),NEE吸收值隨PAR的升高呈線性較快速增加,這時(shí)光是唯一的環(huán)境限制因素;300<PAR<1200μmol/(m2·s)時(shí),NEE吸收值隨PAR的升高呈曲線式較緩慢增長(zhǎng),此時(shí)溫度、外界CO2濃度等環(huán)境因素成為共同的限制因素;PAR>1200μmol/(m2·s)時(shí),NEE吸收量不再隨著PAR的升高而增長(zhǎng),即達(dá)到光合作用的光飽和階段。這與許大全等[19]的研究結(jié)果相一致。
2.5 水分因素對(duì)NEE的影響
水汽壓差VPD間接地代表空氣的水分狀況,有研究指出高光強(qiáng)和高水汽飽和差是植物光合作用降低的兩個(gè)條件[20]。將生長(zhǎng)季VPD觀測(cè)數(shù)據(jù)劃分為3個(gè)區(qū)間:低區(qū)(0.5~1.0 k Pa)、中區(qū)(1.0~1.5 k Pa)、高區(qū)(1.5~2.0 kPa),VPD由低區(qū)到高區(qū)曲線斜率依次增大-減小-增大(圖5)。VPD在低區(qū)時(shí),隨著VPD值的升高,碳吸收速率增加,即更有利于生態(tài)系統(tǒng)的碳蓄積;VPD在中區(qū)時(shí),NEE吸收速率隨VPD的增加繼續(xù)升高,但增加幅度越來(lái)越小,二者變化趨于飽和;VPD處于高區(qū)時(shí),NEE吸收速率隨VPD增加開(kāi)始降低,不利于生態(tài)系統(tǒng)的碳蓄積,原因可能是高水汽壓導(dǎo)致氣孔部分關(guān)閉,氣孔阻力增大,光合作用降低。進(jìn)而影響草甸生態(tài)系統(tǒng)的生產(chǎn)力[21]。
降水是影響生態(tài)系統(tǒng)碳交換的重要水分因子,Lawerence等[22]在對(duì)加拿大北部溫帶C3草地生態(tài)系統(tǒng)連續(xù)觀測(cè)結(jié)果表明,降水充足的年份CO2日吸收峰值約為干旱條件下的3倍。同時(shí)降雨格局的差異也會(huì)影響生態(tài)系統(tǒng)的碳交換能力,以2014年8月16日(短時(shí)強(qiáng)降雨)、22日(短時(shí)弱降雨)、28日(持續(xù)降雨)為例,探討光照強(qiáng)度、降雨對(duì)NEE的共同影響(圖6)。

圖5 凈碳交換NEE與飽和水汽壓差VPD的關(guān)系Fig.5 Relationships between net ecosystem CO2exchange(NEE)and vapor pressure deficit(VPD)

圖6 降雨對(duì)NEE日變化的影響Fig.6 Diurnal variation of CO2exchange(NEE)under different rainfall conditions
8月16日,降雨開(kāi)始前3 h內(nèi),草甸呼吸速率由0.35 mg/(m2·s)迅速降到0.02 mg/(m2·s),降幅達(dá)94%。受凌晨降雨影響,白天PAR僅在0~600μmol/(m2·s)范圍波動(dòng),而表層土壤含水量出現(xiàn)15.8%的小幅遞增,CO2日吸收峰值為0.79 mg/(m2·s),出現(xiàn)在雨后14:00,夜間NEE與雨前碳排放速率大體持平。8月22日,白天NEE與PAR呈明顯“單峰”變化,PAR極值達(dá)1347μmol/(m2·s),正午12:30左右,到達(dá)碳吸收峰值0.93 mg/(m2·s),而夜間21:00-22:00發(fā)生了5.8 mm的微量降雨,但未對(duì)碳排放造成明顯影響。8月28日,降雨歷時(shí)15.5 h,凈輻射(Rn)與PAR均受其影響,數(shù)值僅在0~50 W/m2和0~300μmol/(m2·s)范圍內(nèi)波動(dòng),表層土壤含水量在降雨開(kāi)始3 h內(nèi)緩慢增加,而后穩(wěn)定。午間13:00左右隨雨量增加,生態(tài)系統(tǒng)碳匯功能降低(-0.48 mg/m2·s)。夜間受降雨影響,生態(tài)系統(tǒng)呼吸速率明顯上升。可以看出,降雨削弱了到達(dá)地面的輻射強(qiáng)度,卻增加了表層土壤水含量,進(jìn)而影響生態(tài)系統(tǒng)碳交換。短暫強(qiáng)降雨過(guò)程使白天生態(tài)系統(tǒng)碳匯功能降低,而連續(xù)低強(qiáng)度降雨雖然使生態(tài)系統(tǒng)處于低光強(qiáng)(0<PAR<300μmol/m2·s)的環(huán)境,但提高了草甸生態(tài)系統(tǒng)夜間的碳交換能力,增加了地溫對(duì)夜間呼吸的控制[23]。
3 討論
生長(zhǎng)季NEE日變化呈明顯“倒鐘型”,全天出現(xiàn)一個(gè)吸收高峰。日出后的6:00-6:30,草甸植被便開(kāi)始吸收CO2,8:00-10:30之間達(dá)到碳吸收高峰,午間的11:00-13:00由于光飽和作用,碳吸收量減小,出現(xiàn)群落“午休”現(xiàn)象,這與張法偉等[24]研究的青海湖高寒草甸光合午休時(shí)間較為一致。
12月至翌年3月草甸植被尚未復(fù)蘇,4月中下旬植被才開(kāi)始返青,草甸系統(tǒng)碳吸收微弱,土壤呼吸作用顯著。曾有研究指出,非生長(zhǎng)季土壤呼吸是生長(zhǎng)季生態(tài)系統(tǒng)釋放CO2的一個(gè)重要組成部分,顯著影響著系統(tǒng)的碳收支[25],陳驥等[26]在對(duì)青海湖北岸高寒草甸碳收支的研究中發(fā)現(xiàn),非生長(zhǎng)季土壤呼吸速率是生長(zhǎng)季的土壤呼吸速率的0.30~0.33倍。而生長(zhǎng)季5-7月生態(tài)系統(tǒng)白天的碳吸收強(qiáng)度主要受控于光合有效輻射,二者表現(xiàn)出良好的直角雙曲線關(guān)系(NEE=+0.304,R2=0.73),而夜間則通過(guò)擬合出Vants Hoff方程(公式2)參數(shù),來(lái)估算草甸生態(tài)系統(tǒng)的呼吸速率,本文利用已獲試驗(yàn)數(shù)據(jù)經(jīng)擬合確定Q10為2.63,從而建立起碳通量與土壤溫度的指數(shù)擬合關(guān)系(Reco=0.05eIn(2.63)(Ts-10)/10,R2=0.67)。
4 結(jié)論
1)科爾沁草甸生態(tài)系統(tǒng)碳通量在全年呈現(xiàn)“兩峰一谷”的變化規(guī)律,草甸生態(tài)系統(tǒng)碳吸收總量218.52 g CO2/m2。非生長(zhǎng)季主要表現(xiàn)為碳源,凈CO2日排放速率均值0.11 mg/(m2·s),生長(zhǎng)季主要表現(xiàn)為碳匯,凈CO2吸收速率均值-0.17 mg/(m2·s),較北美大草原的-0.88 mg/(m2·s)值偏小[27],這可能與海拔及葉面積指數(shù)的差異有關(guān)[17]。NEE日變化呈明顯的“單峰型”,且日出、日落前后達(dá)到碳收支平衡狀態(tài)。7月12日,10月9日分別出現(xiàn)CO2日吸收(-0.89 mg/m2·s)和日釋放(0.56 mg/m2·s)的峰值。
2)葉面積指數(shù)對(duì)草甸草原生長(zhǎng)季NEE的變化影響明顯,LAI<0.97時(shí),草甸CO2吸收量隨LAI的增加而線性增長(zhǎng),當(dāng)0.97<LAI<3.08時(shí),CO2吸收量又隨LAI的增加而漸進(jìn)飽和,PAR與LAI的交互作用顯著(P<0.01),葉面積指數(shù)越大,凈碳交換NEE對(duì)PAR的響應(yīng)也越明顯。
3)光合有效輻射也是影響NEE的重要環(huán)境因子。低光強(qiáng)下,生長(zhǎng)季的NEE-PAR呈顯著線性相關(guān)(R2=0.924),生長(zhǎng)季表觀量子效率均值為0.0015μmol CO2/μmol PAR,而在PAR>1200μmol/(m2·s)時(shí),NEE對(duì)光強(qiáng)的響應(yīng)表現(xiàn)為漸進(jìn)飽和型;同時(shí)草甸生態(tài)系統(tǒng)的碳匯功能受到高VPD值的抑制;短暫強(qiáng)降雨增加了表層土壤水分含量,但削弱了到達(dá)地面的輻射強(qiáng)度,降低了晝間土壤呼吸速率,但對(duì)夜間NEE無(wú)明顯影響。而持續(xù)降雨使表層土壤含水量維持在較高水平,雖引起晝間碳交換能力的降低,但激發(fā)了夜間土壤呼吸速率的增加。
[1] Rank A B,Dugas W A.Carbon dioxide fluxes over a northern,semiarid,mixed grass prairie.Agricultural and Forest Meteorology,2001,108:317-326.
[2] Dugas W A,Heuer M L,Msyeux H S.Carbon dioxide fluxes over Bermudagrass,native prairie,and sorghum.Agricultural and Forest Meteorology,1999,93:121-139.
[3] Shaver G R,Rastetter E B,Salmon V,et al.Pan-Arctic modeling of net ecosystem exchange of CO2.Philosophical Transactions of the Royal Society B-biolonical Sciences,2013,368:1-13.
[4] Fu Y L,Yu G R,Wang Y F,et al.Impact of water stress on photosynthesis and respiration over Leymus chinensis ecosystem in Inner Mongolia.Science in China(Ser.D)Earth Sciences,2006,36(Supp,I):183-193.
[5] Hao Y B,Wang Y F,Sun X M,et al.Seasonal variation in carbon exchange and its ecological analysis over Leymus chinensis steppe in Inner Mongolia.Science in China(Ser.D)Earth Sciences,2006,36:174-182.
[6] Zhao Y P,Zhang X Z,Shi P L,et al.Impact of drought stress on net CO2exchange above an alpine grassland ecosystem in the Gentral Tibetan Plateau.Journal of Resources and Ecology,2013,4(4):327-336.
[7] Li Y Q,Zhao H L,Zhao X Y,et al.Responses of soil respiration to air temperature from June to September in different sanddunes,Horqin sandy land.Journal of Desert Research,2008,28(2):249-254.
[8] Chen N N,Guan D X,Jin C J,et al.Characteristics of soil respiration on Horqin meadow.Chinese Journal of Grassland,2011,33(5):82-87.
[9] LI-COR Biosciences,Eddy Pro 4.0:Help and User’s Guide.Lincoln[EB/OL].[2013-12-02].http://envsupport.licor.com/help/EddyPro4/Default.htm.
[10] Burba G G,McDermitt D K.Addressing the influence of instrument surface heat exchange on the measurements of CO2flux from open-path gas analyzers.Global Change Biology,2008,14:1854-1876.
[11] Li Y J,Xu Z Z,Wang Y L,et al.Latent and sensible heat fluxes and energy balance in a maize agroecosystem.Chinese Journal of Plant Ecology,2007,31(6):1132-1144.
[12] Falge E,Baldocchi D D,Olson R J,et al.Gap filling strategies for defensible annual sums of net ecosystem exchange.Agricultural and Forest Meteorology,2001,107:43-69.
[13] Raich J W,Schlesinger W H.The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate.Tellus,1992,44:81-99.
[14] Xu L J,Tang H J,Yang G X,et al.Variation of net ecosystem carbon flux and its impact factors on Stipa baicalensis steppe in the growing season.Acta Prataculturae Sinica,2011,20(6):287-292.
[15] Shi P L,Sun X M,Xu L L,et al.Net ecosystem CO2exchange and controlling factors in a Steppe-Kobresia meadow on Tibetan Plateau.Science in China(Ser.D),2006,36(Supp,I):194-203.
[16] Suyker A E,Verma S B,Burba G G,et al.Gross primary production and ecosystem respiration of irrigated maize and irrigated soybean during a growing season.Agricultural and Forest Meteorology,2005,131(3/4):180-190.
[17] Chen J L,Wang J,Wang Q,et al.LAI and light transmittance measurement of isolated Vitex shrub using LAI-2200 PCA.Chinese Journal of Agrometeorology,2014,35(4):373-379.
[18] Xu L L,Zhang X Z,Shi P L,et al.Photoionization efficiency in response to temperature under low pressure of alpine meadow on Tibetan Platean.Science in China(Ser.D),2006,36(Supp,II):204-209.
[19] Xu D Q,Li D Y,Shen Y G,et al.On midday depression of photosynthesis of wheat leaf under field conditions.Acta Phytophysiologia Sinica,1984,10:269-276.
[20] Bunce J A.Afternoon inhibition of photosynthesis in maize.Field Crop Research,1990,24:261-271.
[21] Fu Y L,Yu G R,Sun X M,et al.Depression of net ecosystem CO2exchange in semi-arid Leymus chinensis steppe and alpine shrub.Agricultural and Forest Meteorology,2006,137:234-244.
[22] Lawerence B,F(xiàn)lanagan,Linda A,et al.Seasonal and interannual variation in carbon dioxide exchange and carbon balance in a northern temperate grassland.Global Change Biology,2002,8:599-615.
[23] Zhang F W,Li Y N,Zhao X Q,et al.Effect of one precipitation process on CO2flux and themal transportation in alpine meadow of Qinghai-Tibetan plateau.Chinese Journal of Ecology,2008,27(10):1685-1691.
[24] Zhang F W,Li Y N,Cao G M,et al.CO2fluxes and their driving factors over alpine meadow grassland ecosystems in the northern shore of Qinghai Lake,China.Chinese Journal of Plant Ecology,2012,36(3):187-198.
[25] Wang W,Wang T,Peng S S,et al.Review of winter CO2efflux from soils:a key process of CO2exchange between soil and atmosphere.Chinese Journal of Plant Ecology,2007,31(3):394-402.
[26] Chen J,Cao J J,Wei Y L,et al.Effect of grazing exclusion on soil respiration during the dormant in alpine meadow grassland ecosystems on the northern shore of Qinghai Lake,China.Acta Prataculturae Sinica,2014,23(6):78-86.
[27] Verma S B,Kim J,Clement R J,et al.Water vapor and carbon dioxide exchange at a centrally located prairie site during FIFE.Journal of Geophysics Research,1992,97:18629-18639.
參考文獻(xiàn):
[4] 付玉玲,于貴瑞,王艷芬.水分脅迫對(duì)內(nèi)蒙古羊草草原光合和呼吸作用的影響.中國(guó)科學(xué)D輯,地球科學(xué),2006,36(增刊I):183-193.
[5] 郝彥賓,王艷芬,孫曉敏,等.內(nèi)蒙古羊草草原碳交換季節(jié)變異及其生態(tài)學(xué)解析.中國(guó)科學(xué),2006,36:174-182.
[7] 李玉強(qiáng),趙哈林,趙學(xué)勇,等.科爾沁沙地夏秋(6-9月)季不同類(lèi)型沙丘土壤呼吸對(duì)氣溫變化的響應(yīng).中國(guó)沙漠,2008,28(2):249-254.
[8] 陳妮娜,關(guān)德新,金昌杰,等.科爾沁草甸草地土壤呼吸特征.中國(guó)草地學(xué)報(bào),2011,33(5):82-87.
[11] 李祎君,許振柱,王云龍,等.玉米農(nóng)田水熱通量動(dòng)態(tài)與能量閉合分析.植物生態(tài)學(xué)報(bào),2007,31(6):1132-1144.
[14] 徐麗君,唐華俊,楊桂霞,等.貝加爾針茅草原生態(tài)系統(tǒng)生長(zhǎng)季碳通量及其影響因素分析.草業(yè)學(xué)報(bào),2011,20(6):287-292.
[15] 石培禮,孫曉敏,徐玲玲,等.西藏高原草原化嵩草草甸生態(tài)系統(tǒng)CO2凈交換及其影響因子.中國(guó)科學(xué)D輯,地球科學(xué),2006,36(增刊I):194-203.
[17] 陳景玲,王靜,王謙,等.植物冠層分析儀測(cè)定荊條孤立冠層葉面積指數(shù)和透光率的分析.中國(guó)農(nóng)業(yè)氣象,2014,35(4):373-379.
[18] 徐玲玲,張憲洲,石培禮,等.青藏高原低氣壓條件下高寒草甸群落光量子效率對(duì)溫度的響應(yīng).中國(guó)科學(xué)D輯,地球科學(xué),2006,36(增刊I):204-209.
[19] 許大全,李德耀,沈允剛,等.田間小麥葉片光合作用“午睡”現(xiàn)象的研究.植物生理學(xué)報(bào),1984,10:269-276.
[23] 張法偉,李英年,趙新全,等.一次降水過(guò)程對(duì)青藏高原高寒草甸CO2通量和熱量輸送的影響.生態(tài)學(xué)雜志,2008,27(10):1685-1691.
[24] 張法偉,李英年,曹廣民,等.青海湖北岸高寒草甸草原生態(tài)系統(tǒng)CO2通量特征及其驅(qū)動(dòng)因子.植物生態(tài)學(xué)報(bào),2012,36(3):187-198.
[25] 王娓,汪濤,彭書(shū)時(shí),等.冬季土壤呼吸:不可忽視的地氣CO2交換過(guò)程.植物生態(tài)學(xué)報(bào),2007,31(3):394-402.
[26] 陳驥,曹軍驥,魏永林,等.青海湖北岸高寒草甸草原非生長(zhǎng)季土壤呼吸對(duì)溫度和濕度的響應(yīng).草業(yè)學(xué)報(bào),2014,23(6):78-86.
Net ecosystem carbon exchange and its main driving factors in meadow grassland ecosystems in the Horqin Sand Lands
WANG Jing1,LIU Ting-Xi1*,LEI Hui-Min2*,ZHANG Sheng-Wei1,DUAN Li-Min1
1.Water Resources and Hydropower Survey and Design Institude,Inner Mongolia Agricultural University,Hohhot 010018,China;2.Department of Hydraulic Engineering,Tsinghua University,Beijing 100084,China
The objectives of this study were to investigate the main drivers of ecosystem carbon dynamics in meadow grasslands in the Horqin Sand Lands,and to understand the relationship between net ecosystem CO2exchange(NEE)and key meteorological factors in this region.We continuously measured(NEE)and environmental factors in meadow grasslands in the Horqin Sand Lands from October 2013 to September 2014 using the eddy covariance method.The results showed that the diurnal variation in NEE differed significantly among seasons,especially among vegetation growing seasons.The highest seasonal diurnal absorption rate of CO2was in July,followed by August,September,June,and then May.The NEE was controlled by photosynthetically active radiation(PAR)and the leaf area index(LAI)during vegetation growing seasons.The relationship between NEE and PAR was expressed by a rectangular hyperbolic function.The simulated apparent initial average light-use efficiency was 0.0015μmol CO2/μmol PAR,and the maximum photosynthetic rate(Pmax)was 0.65μmol CO2/(m2·s).The NEE was influenced by LAI,which was expressed by piecewise functions.When the LAI was greater than 3.08,NEE-PAR showed a gradual saturation-type relationship.The higher the LAI,the stronger the response of net carbon exchange to the amount of PAR.A light saturation response occurred when 1.5<VPD<2 k Pa,and NEE was significantly inhibited under high VPD.The day-time net CO2exchange was inhibited by heavy rainfall(cumulative rainfall>40 mm/day),while sustained low-intensity rainfall(rainfall length>15 h)stimulated soil respiration at night.The night-time respiration rate of the ecosystem(Reco)increased exponentially as the soil temperature(Ts)increased,and the temperature-sensitive coefficient Q10was 2.63.
net ecosystem exchange(NEE);meteorological factors;eddy covariance;Horqin sand
10.11686/cyxb2014530 http://cyxb.lzu.edu.cn
王婧,劉廷璽,雷慧閩,張圣微,段利民.科爾沁草甸生態(tài)系統(tǒng)凈碳交換特征及其驅(qū)動(dòng)因子.草業(yè)學(xué)報(bào),2015,24(11):10-19.
WANG Jing,LIU Ting-Xi,LEI Hui-Min,ZHANG Sheng-Wei,DUAN Li-Min.Net ecosystem carbon exchange and its main driving factors in meadow grassland ecosystems in the Horqin Sand Lands.Acta Prataculturae Sinica,2015,24(11):10-19.
2014-12-19;改回日期:2015-03-10
國(guó)家自然科學(xué)基金重點(diǎn)資助項(xiàng)目(51139002),教育部創(chuàng)新團(tuán)隊(duì)發(fā)展計(jì)劃(IRT13069)和內(nèi)蒙古農(nóng)業(yè)大學(xué)寒旱區(qū)水資源利用創(chuàng)新團(tuán)隊(duì)(NDTD2010-6)資助。
王婧(1989-),女,內(nèi)蒙古阿拉善人,在讀碩士。E-mail:lebishu1234@163.com
*通訊作者Corresponding author.E-mail:txliu1966@163.com,leihm@m(xù)ail.tsinghua.edu.cn
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
文苑(2018年22期)2018-11-19 02:54:14
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
