雙膦酸鹽對NFATc1及相關因子p-NF-κB、p-c-Jun在破骨細胞分化過程中的影響
2015-06-28 15:41:38董偉彭宏峰梁永強馮曉潔廖囡囡戚孟春
解放軍醫學雜志 2015年10期
關鍵詞:檢測
董偉,彭宏峰,梁永強,馮曉潔,廖囡囡,戚孟春
論 著
雙膦酸鹽對NFATc1及相關因子p-NF-κB、p-c-Jun在破骨細胞分化過程中的影響
董偉,彭宏峰,梁永強,馮曉潔,廖囡囡,戚孟春
目的研究雙膦酸鹽(ALN)對破骨細胞(OC)分化過程中關鍵因子NFATc1及其信號分子p-NF-κB、p-c-Jun的影響。方法采用小鼠單核巨噬細胞RAW264.7誘導培養OC,細胞培養于培養皿,分A組(對照組)、B組(ALN組)。培養第2d采用Western blotting檢測NFATc1、p-NF-κB、p-c-Jun基因表達變化情況,培養第4d采用免疫熒光方法檢測NFATc1表達,培養第7d檢測各組OC生成及數目。采用牙本質磨片接種RAW264.7細胞,分組及處理方法同上述培養皿培養過程,第9d檢測骨吸收功能。結果兩組細胞均有TRAP陽性多核OC生成,并在牙本質磨片上形成吸收陷窩;但A組TRAP陽性多核細胞數目、吸收陷窩數目及陷窩面積均大于B組。免疫熒光檢測NFATc1表達A組高于B組;Western blotting檢測NFATc1、p-NF-κB、p-c-Jun表達B組較A組低,兩組有顯著性差異(P<0.01)。結論雙膦酸鹽通過下調p-NF-κB、p-c-Jun的表達,最終抑制NFATc1的表達,從而抑制OC生成和骨吸收功能。
阿侖膦酸鹽;破骨細胞;NFATc1;NF-κB;基因,jun
破骨細胞的功能分化過程是當今研究的重點[1],許多骨骼疾病如骨質疏松癥[2]、類風濕性關節炎、Paget's病、多發性骨髓瘤、惡性腫瘤的溶骨性骨轉移等,都與破骨細胞(osteoclasts,OC)增殖、骨吸收功能亢進密切相關[3]。阿侖膦酸鹽(alendronate,ALN)作為第三代含氮雙膦酸鹽,是現階段治療上述疾病的首選藥物,除此之外還被用作預防和治療骨密度降低及成骨不全癥等疾病[4]。ALN主要通過抑制破骨細胞骨吸收功能、誘導破骨細胞凋亡來發揮作用,但確切機制及涉及的胞內信號通路目前知之甚少,特別是對破骨細胞分化過程中關鍵信號分子NFATc1及其重要通路如NF-κB通路、MKK通路是否產生影響等。本研究在OC培養過程中加入ALN并檢測NFATc1的表達,分析影響其表達的p-c-Jun、p-NF-κB,以初步揭示雙膦酸鹽抑制OC的分子機制,為ALN的臨床應用提供理論基礎。
1 材料與方法
1.1 實驗材料 DMEM培養基(Gibco,美國),RANKL(receptor activator of nuclear factor κB ligand) (Biovision,美國),阿侖膦酸鹽(Sigma,美國),兔抗鼠NFATc1、p-c-Jun、p-NF-κB多克隆抗體(Santa Cruz,美國),羊抗兔辣根過氧化物酶標記IgG和RIPA(武漢博士德,中國),抗酒石酸酸性磷酸酶(TRAP)染色試劑盒(Sigma,美國),RAW264.7小鼠單核巨噬細胞系(中國科學院細胞庫)。
1.2 OC的培養及實驗分組 RAW264.7細胞以1×105/孔的密度接種于48孔培養板中,DMEM培養基(含15%胎牛血清、100U/ml青霉素、100μg/ ml鏈霉素)培養,細胞貼壁后分為兩組,對照組加入100ng/ml RANKL;處理組加入100ng/ml RANKL+5×10-7mol/L ALN,定期于倒置相差顯微鏡下觀察。
1.3 抗酒石酸酸性磷酸酶(TRAP)染色 接種于48孔板的細胞于處理后第7d收獲,按TRAP染色試劑盒操作步驟進行染色,顯微鏡100倍放大倍數下隨機選取5個視野計數TRAP染色陽性細胞(細胞核≥3 個)數目,5個視野計數取平均值為該細胞爬片的破骨細胞數目。
1.4 牙本質磨片制備及吸收陷窩檢測 將人離體牙制備成直徑約0.5cm,厚度約0.2mm的牙本質磨片,打磨拋光表面,消毒滅菌后將RAW264.7細胞接種其上,細胞貼壁后分為兩組(同1.2細胞處理方法)。取處理后第9天的牙本質磨片,2.5%戊二醛4℃固定7min,1mol/L氫氧化胺中以50Hz超聲清洗5min,再經蒸餾水超聲清洗5min×3次;隨后2.5%戊二醛固定2h,1%鋨酸固定2h,乙醇逐級脫水,醋酸異戊酯置換,CO2臨界點干燥,鍍金,掃描電鏡(HITACHIS-4800)觀察吸收陷窩。在500倍放大倍數下每孔牙本質磨片上隨機選取5個視野,用醫學數碼圖像分析系統Med6.0測量五個視野吸收陷窩總數目及總面積。
1.5 免疫熒光法檢測NFATc1表達 取培養至第4d的RAW264.7細胞,PBS沖洗,4%多聚甲醛固定10min,0.5%Triton穿孔15min,1%BSA封閉30min,PBS沖洗5min×2次。加入5%BSA稀釋的兔抗鼠NFATc1多克隆一抗(1:25)過夜。PBS沖洗,加入5%BSA稀釋的FITC標記的羊抗兔IgG,37℃孵育2h,5μg/ml DAPI染色2min,于共聚焦顯微鏡下觀察。
1.6 Western blotting檢測NFATc1、p-NF-κB、p-c-Jun蛋白表達 兩組細胞培養第2天,提取細胞總蛋白BCA法定量。上樣量35μg,采用12%SDS-PAGE凝膠電泳分離,采用濕法轉膜至PVDF膜,5%BSA室溫封閉1h,分別加入1μg/ml兔抗鼠NFATc1、p-NF-κB、p-c-Jun多克隆一抗孵育過夜,羊抗兔辣根過氧化物酶標記IgG二抗結合室溫孵育1h。TBST洗膜后按化學發光試劑盒說明書制備反應液,室溫孵育5min。用Image J分析軟件對條帶吸光度值(A)進行半定量分析,以目的蛋白條帶(A)值/內參GAPDH蛋白條帶(A)值的比值表示目的蛋白的相對量。
1.7 統計學處理 采用SPSS 11.0軟件進行統計分析。計量數據以表示,組間比較采用SNK-q檢驗,P<0.05為差異有統計學意義。
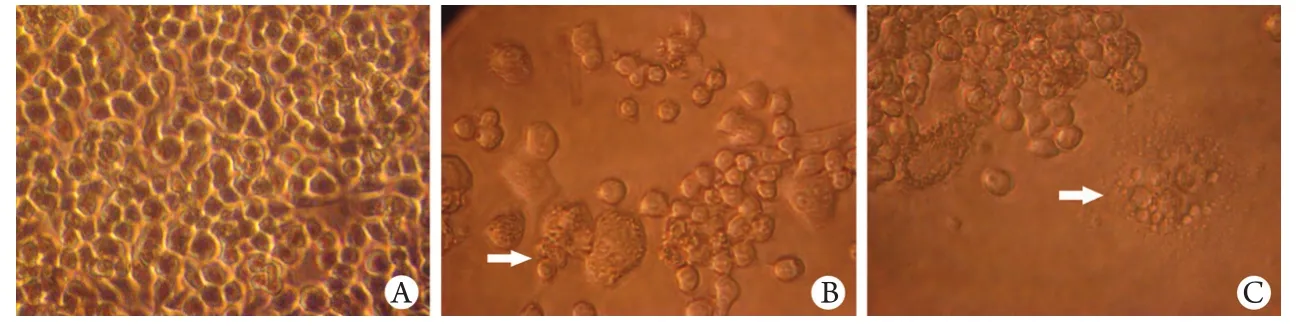
圖1 倒置相差顯微鏡下觀察RAW264.7細胞生長情況(×400)Fig.1 RAW264.7 cell growth observed under inverted phase contrast microscope (×400) A. Incubated for 1 day; B. Incubated for 3 days; C. Incubated for 7 days
2 結 果
2.1 倒置相差顯微鏡觀察OC生成情況 培養開始時,貼壁的RAW264.7呈圓形鋪于培養板底壁(圖1A)。培養至第3天,在誘導因子RANKL的作用下,RAW264.7開始相互融合(圖1B)。培養至第7d,細胞融合成多核OC(圖1C)。
2.2 TRAP染色及定量分析 對照組OC生成數量顯著高于處理組(P<0.01),提示ALN可顯著抑制OC的生成(圖2)。
2.3 牙本質吸收陷窩觀察及定量分析 兩組比較,對照組OC吸收陷窩數目較多,陷窩總面積較大,兩組間吸收陷窩數目及總面積均有顯著性差異(P<0.01,圖3,表1)。上述結果表明,ALN可有效抑制OC的骨吸收功能。
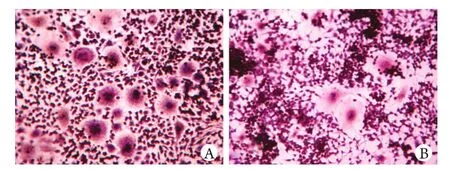
圖2 細胞培養第7天的TRAP染色比較(×200)Fig. 2 TRAP staining of osteoclasts on the 7th day (×200)A. Control group; B. ALN-treated group

圖3 細胞培養第9天骨吸收陷窩形成情況(SEM×500)Fig. 3 Resorption lacunaes formed on dentin slices after culture for 9 days (SEM×500) A. Control group; B. ALN-treated group
2.4 免疫熒光檢測NFATc1表達 兩組細胞培養至4d,免疫熒光檢測見NFATc1表達(圖4),對照組NFATc1熒光強度(5.42±0.63)明顯大于處理組(1.26±0.15)(P<0.01)。
2.5 Western blotting檢測NFATc1,p-NF-κB,p-c-Jun蛋白的表達 Western blotting檢測兩組細胞NFATc1、p-NF-κB、p-c-Jun蛋白含量變化,處理組NFATc1、p-NF-κB、p-c-Jun蛋白表達較對照組明顯減弱,條帶灰度值分別下降47.1%、39.3%、18.4%;結果顯示,ALN可抑制OC細胞NFATc1、 p-NF-κB、p-c-Jun的表達。

表1 TRAP陽性破骨細胞計數及牙本質磨片吸收陷窩計數和陷窩面積Tab.1 The number of TRAP(+) osteoclasts, number and size of resorption lacunaes formed on dentin slices in each group
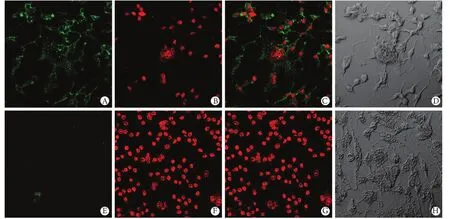
圖4 免疫熒光化學法檢測兩組NFATc1的表達(×400)Fig. 4 Detection of NFATc1 expression by immunofluorescent chemistry (×400) A, E. NFATc1 expression labelled by FITC; B, F. Cell nucleus labelled by DAPI; C, G. Integrated image; D, H. Normal vision without fluorescence

圖5 Western blotting檢測兩組NFATc1、p-NF-κB、p-c-Jun蛋白表達Fig.5 Detection of NFATc1, p-NF-κB, p-c-Jun protein expression in two groups of cells by Western blotting
3 討 論
OC是骨吸收時唯一的效應細胞,由多功能造血干細胞在誘導因子(RANKL)的誘導下相互融合形成的多核巨細胞。在OC形成過程中,眾多因子及信號通路發揮重要作用[5-7],其中活化T細胞核因子NFATc1為破骨細胞分化的重要信號分子[8],而NF-κB及MKK是影響NFATc1較為關鍵的信號通路分子[9-10]。
在OC分化中,當RANKL與其受體即OC前體上的RANK相互作用后,引起了下游一系列信號因子的級聯作用[11]。首先,引發鈣/鈣調磷酸酶/活化T細胞核因子(calcium/calcineurin/NFAT)活化,導致胞質內Ca2+波動,Ca2+/鈣調蛋白依賴的鈣調磷酸酶被激活,使胞質內NFATc1蛋白異位進入細胞核[12];同時,NF-κB參與作用的經典途徑(classical pathway)和替代途徑(alternative pathway)被激活。在經典途徑中,由IKKβ和NEMO/IKKγ組成的IKK復合物是關鍵信號因子,它的作用是誘導在細胞質中與無活性NF-κB形成復合物的抑制蛋白IκB磷酸化,隨后NF-κB被釋放,形成二聚體p65/cRel[13];而替代途徑的關鍵蛋白是NF-κB誘導激酶NIK(NF-κB-inducing kinase),其激活IκB激酶IKKα,促使NF-κB釋放并形成二聚體RelB/p52[13]。上述兩途徑形成的NF-κB二聚體轉移至細胞核中與NFATc2共同作用于NFATc1基因啟動子,誘發NFATc1基因初始表達,隨后NFATc1基因表達進入自發擴增階段;與此同時,在RANKL刺激下,另一通路MKK信號通路啟動,JUN被激活,NF-κB磷酸化,最終c-Jun磷酸化,c-Jun與c-Fos形成催化蛋白-1(AP-1),c-Fos被激活,選擇性地募集到NFATc1基因啟動子上,與NF-κB、NFATc2共同促進NFATc1基因的表達,進而促使多核OC前體的形成[14-15]。隨后,OC前體活化形成成熟的破骨細胞,發揮其骨吸收功能。
ALN是現階段治療骨質疏松等疾病的常用臨床藥物。眾多學者通過動物實驗及細胞實驗兩水平均證實了其對OC分化及骨吸收功能的抑制作用,但其是否對NFATc1表達產生抑制,并影響相關通路的作用,國內外研究并未給出確切結論。本研究結果表明,ALN通過抑制p-NF-κB、p-c-Jun的表達,影響NF-κB參與通路及MKK信號通路,最終下調NFATc1基因表達,從而起到抑制OC生成及骨吸收功能的作用。本實驗部分揭示了雙膦酸鹽對OC生成抑制作用的分子學機制,將更好地為雙膦酸鹽在臨床中的應用提供實驗依據。然而,ALN是否會通過影響鈣/鈣調磷酸酶/活化T細胞核因子(calcium/ calcineurin/NFAT)通路,導致胞質內Ca2+波動受阻,從而阻礙NFATc1發生核異位還需要進一步實驗驗證。
[1]Yang FX, Yang DZ, Zhou JP,et al. Effect of ERK1/2 signal pathway on the expression of OPG/RANKL in cementoblasts under stress stimulation[J]. Med J Chin PLA, 2014, 39(12): 941-945. [楊鳳雪, 楊冬珍, 周建萍, 等. 應力刺激下ERK1/2信號通路對成牙骨質細胞OPG/RANKL表達的影響[J]. 解放軍醫學雜志, 2014, 39(12): 941-945.]
[2]Ma XJ, Kong XL, Meng XJ,et al. Study on the related factors of affecting the adult female osteoporosis[J]. Chin J Pract Intern Med, 2014, 34(S1): 78-81. [馬小靜, 孔祥雷, 孟祥菊, 等. 影響成年女性骨質疏松的相關因素研究[J]. 中國實用內科雜志, 2014, 34(S1): 78-81.]
[3]Ikeda F, Nishimura R, Matsubara T,et al. Critical roles of c-Jun signaling in regulation of NFAT family and RANKL-regulated osteoclast differentiation[J]. J Clin Invest, 2004, 114(4): 475-484.
[4]Zhu HM. Bisphosphonates and osteoporosis[J]. Chin J Pract Intern Med, 2011, 31(7): 517-520. [朱漢民. 雙膦酸鹽和骨質疏松[J]. 中國實用內科雜志, 2011, 31(7): 517-520.]
[5]Kobayashi, N, Kadono Y, Naito A,et al. Segregation of TRAF6-mediated signaling pathways clarifies its role in osteoclastogenesis[J]. EMBO J, 2001, 20(6): 1271-1280.
[6]Kim JH, Kim K, Youn BU,et al. MHC class Ⅱ transactivator negatively regulates RANKL-mediated osteoclast differentiation by downregulating NFATc1 and OSCAR[J]. Cell Signal, 2010, 22(9): 1341-1349.
[7]Asagiri M, Takayanagi H. The molecular understanding of osteoclast differentiation[J]. Bone, 2007, 40(2): 251-264.
[8]Yuroda Y, Hisatsune C, Nakamura T,et al. Osteoblasts induce Ca+ oscillation-independent NFATc1 activation during osteoclastogenesis[J]. Proc Natl Acad Sci USA, 2008, 105(25): 8643-8648.
[9]Franzoso, G, Carlson L, Xing L,et al. Requirement for NF-kappaB in osteoclast and B-cell development[J]. Genes Dev, 1997, 11(24): 3482-3496.
[10] Miyazaki M, Fujikawa Y, Takita C,et al. Tacrolimus and cyclosporine A inhibit human osteoclast formationviatargeting the calcineurin-dependent NFAT pathway and an activation pathway for c-Jun or MITF in rheumatoid arthritis[J]. Clin Rheumatol, 2007, 26(2): 231-239.
[11] Huang H, Ryu J, Ha J,et al. Osteoclast differentiation requires TAK1 and MKK6 for NFATc1 induction and NF-κB transactivation by RANKL[J]. Cell Death Differ, 2006, 13(11), 1879-1891.
[12] Komarova SV, Pilkington MF, Weidema AF,et al. RANK ligand-induced elevation of cytosolic Ca2+accelerates nuclear translocation of NF-kB inosteoclasts[J]. J Biol Chem, 278(10): 8286-8293.
[13] Deborah V N. Role of NF-κB in the skeleton[J]. Cell Research, 2011, 21(1): 169-182.
[14] Asagiri M, Sato K, Usami T,et al. Autoamplification of NFATc1 expression determines its essential role in bone homeostasis[J]. J Exp Med, 202(9): 1261-1269.
[15] Matsuo K, Galson DL, Zhao C,et al. Nuclear factor of activated Tcells (NFAT) rescues osteoclasto- genesis in precursors lacking c-Fos[J]. J Biol Chem, 279(25): 26475-26480.
Effect of bisphosphonates on NFATc1 and correlators p-NF-κB and p-c-Jun in osteoclast differentiation
DONG Wei, PENG Hong-feng, LIANG Yong-qiang, FENG Xiao-jie, LIAO Nan-nan, QI Meng-chun*
School of Stomatology, North China University of Science and Technology, Tangshan 063000, China
*< class="emphasis_italic">Corresponding author, E-mail: qimengchun@163.com
, E-mail: qimengchun@163.com
This work was supported by the National Natural Science Foundation of China (81270965) and the Natural Science Foundation of Hebei Province (C2011401044)
ObjectiveTo study the effect of alendronate (ALN) on NFATc1 and correlated signaling factors p-NF-κB and p-c-Jun in osteoclast differentiation.MethodsOsteoclasts were inductively cultivated with mouse mononuclear macrophage RAW264.7, and they were divided into 2 groups: group A (control group) and group B (ALN-treated group). The protein expressions of NFATc1, p-NF-κB and p-c-Jun were determined with Western blotting at the 2nd day of cultivation; the expression of NFATc1 was assessed by immunofluorescent assay on the 4th day; and the osteoclast formation was examined on the 7th day of cultivation. RAW264.7 cells were inoculated on abrasive dentine disk, and divided into 2 groups and treated as mentioned above. The resorption function of osteoclast was observed on the 9th day of inoculation.ResultsTRAP positive multinuclear osteoclasts were observed, and resorption lacunaes formed in the abrasive dentine disks of the 2 groups. More TRAP positive multinuclear cells and resorption lacunaes in large size were found in group A than those in group B. Immunofluorescence assay showed the expression of NFATc1 was higher in group A than in group B. The gene expressions of NFATc1, p-NF-κB and p-c-Jun were lower in group B than in group A (P<0.01) as determined with Western blotting.ConclusionBy down-regulating the expressions of p-NF-κB and p-c-Jun, ALN may strongly inhibit the osteoclast formation and its resorption function, thus inhibiting NFATc1 expression.
alendronate; osteoclast; NFATc1; NF-κB; genes, jun
R329.24
A
0577-7402(2015)10-0778-04
10.11855/j.issn.0577-7402.2015.10.02
2014-11-25;
2015-08-20)
(責任編輯:張小利,沈寧)
國家自然科學基金(81270965);河北省自然科學基金(C2011401044)
董偉,醫學碩士,講師。主要從事骨組織工程方面的研究
063000 河北唐山 華北理工大學口腔醫學院(董偉、彭宏峰、梁永強、馮曉潔、廖囡囡、戚孟春)
戚孟春,E-mail: qimengchun@163.com
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
