季風(fēng)常綠闊葉林不同恢復(fù)階段群落優(yōu)勢物種的空間分布格局
2015-07-04 11:14:38李智宏徐崇華劉萬德卯光宇
西北植物學(xué)報 2015年2期
關(guān)鍵詞:物種
李智宏,徐崇華,劉萬德,王 妍,卯光宇
(1 云南太陽河省級自然保護區(qū)管理局,云南普洱665000;2 中國林業(yè)科學(xué)研究院資源昆蟲研究所,昆明650224;3 西南林業(yè)大學(xué)環(huán)境科學(xué)與工程學(xué)院,昆明650224)
種群空間格局是決定群落組成、結(jié)構(gòu)與動態(tài)的重要因素,一直是生態(tài)學(xué)研究熱點之一[1]。分析物種的空間格局有助于認(rèn)識其潛在的生態(tài)學(xué)過程(如種子擴散、種內(nèi)和種間競爭、干擾等)、種群的生物學(xué)特性(如生活史策略、喜光、耐陰等)及其與環(huán)境因子之間的相互關(guān)系(如小生境、植物與生長環(huán)境之適合度、環(huán)境異質(zhì)性等)[2-5]。物種(特別是優(yōu)勢物種)的空間格局對物種的生長、繁殖、死亡、再生、資源利用以及林窗的形成等具有顯著影響[2-3,6]。優(yōu)勢種對群落結(jié)構(gòu)塑造和群落環(huán)境形成有明顯的控制作用[7],既是群落骨架,又決定了群落外貌和群落類型[5],制約著群落資源配置的異化過程[8]。因此,分析優(yōu)勢物種的空間格局有助于認(rèn)識群落結(jié)構(gòu)的形成與維持機制,揭示種群分布格局成因及種群生活史對策,闡明種群及群落的動態(tài)特征及種群與環(huán)境互作過程,為種群保護及恢復(fù)措施的制定提供科學(xué)依據(jù)。
季風(fēng)常綠闊葉林是中國最復(fù)雜、生產(chǎn)力最高、生物多樣性最豐富的地帶性植被類型之一,對保護環(huán)境和維持全球碳平衡等都具有極重要的作用,尤其是在中國亞熱帶地區(qū)的生態(tài)環(huán)境建設(shè),乃至全國的可持續(xù)發(fā)展中占據(jù)舉足輕重的地位[9]。近年來,盡管有關(guān)季風(fēng)常綠闊葉林開展了大量的研究工作[9-21],但有關(guān)季風(fēng)常綠闊葉林物種空間分布格局的研究還沒有開展,特別是退化生態(tài)系統(tǒng)中優(yōu)勢物種的空間分布格局研究。本研究通過對云南普洱不同恢復(fù)階段(恢復(fù)15a、30a和老齡林)季風(fēng)常綠闊葉林優(yōu)勢物種的調(diào)查,分析了其空間分布格局特征,探索了其分布格局隨群落恢復(fù)及尺度的變化特征,旨在闡明種群分布格局與生境條件、種群年齡結(jié)構(gòu)的動態(tài)關(guān)系,揭示種群發(fā)育過程中空間動態(tài)與生物學(xué)機理,為保護和合理經(jīng)營利用提供依據(jù)。
1 研究區(qū)概況和研究方法
1.1 研究區(qū)概況
研究區(qū)域位于云南省中南部的普洱市,地理位置為22°02′~24°50′N,99°09′~102°19′E,海拔317~3 370m。該區(qū)地處熱帶北緣向南亞熱帶的過渡地區(qū),氣候主要受印度洋及太平洋季風(fēng)控制,形成夏秋季多雨、冬春季干旱、年溫差小、日溫差大、干濕季明顯的氣候特征。該地區(qū)年平均氣溫17.7℃,年均降雨量1 547.6mm,雨水主要集中在5~10月,占全年降雨量的87.3%,年平均蒸發(fā)量1 590mm,相對濕度82%。土壤以赤紅壤為主。
季風(fēng)常綠闊葉林是該地區(qū)主要的森林植被類型之一。由于森林距離人類活動區(qū)域較近,季風(fēng)常綠闊葉林常被砍伐作為薪炭、改造成茶園及農(nóng)地等。但近年來,隨著各種森林保護措施的出臺,森林得以不斷恢復(fù),形成了不同恢復(fù)時間的季風(fēng)常綠闊葉林群落。恢復(fù)15a群落主要分布在依像鎮(zhèn)的大寨小平壩,海拔范圍在1 400~1500m 之間,坡位中上,坡度在18°~25°之間,喬木層蓋度在65%~70%之間,優(yōu)勢種為短刺栲(Castanopsisechidnocarpa)、刺栲(C.hystrix)、紅木荷(Schimawallichii)、杯狀栲(C.calathiformis),主要組成物種有小果栲(C.fleuryi)、母豬果(Helicianilagirica)、華南石櫟(Lithocarpusfenestratus)、香面葉(Linderacommunis)等;灌木層蓋度在40%~50%之間,主要組成物種以喬木幼樹為主,如杯狀栲;草本層蓋度在15%~25%之間,主要組成物種有毛果珍珠茅(Sclerialevis)、狗 脊 蕨(Woodwardiajaponica)、山菅蘭(Dianellaensifolia)等。恢復(fù)30a群落主要分布在翠云區(qū)信房水庫,海拔范圍在1 350~1 370 m 之間,坡位中上,坡度在20°~25°之間,喬木層蓋度在70%~85%之間,優(yōu)勢種為短刺栲、刺栲、華南石櫟、紅木荷,主要組成物種有紅花木犀欖(Olearosea)、粗壯潤楠(Machilusrobusta)、隱距越桔(Vaccinium exaristatum)等;灌木層蓋度為65%,絕大多數(shù)為喬木樹種的幼樹、幼苗,以短刺栲等占優(yōu)勢;草本層蓋度在65%~70%之間,主要組成物種有毛果珍珠茅、狗脊蕨、芒萁(Dicranopterispedata)等。老齡林(>200a)主要位于菜陽河自然保護區(qū),海拔范圍在1 250~1 600m 之間,坡位中上,坡度在5°~12°之間,喬木層蓋度在85%~95%之間,優(yōu)勢種主要為短刺栲、刺栲、杯狀栲、紅木荷,主要組成物種有截頭石櫟(Lithocarpustruncatus)、粗穗石櫟(Lithocarpusgrandifolius)、粗壯潤楠等;灌木層蓋度在60%~65%,主要組成物種以喬木幼樹為主,如杯狀栲、短刺栲等;草本層蓋度在30%~45%,主要組成物種是云南復(fù)葉耳蕨(Arachniodeshenryi)、毛果珍珠茅、綠苞山姜(Alpiniabracteata)等。
1.2 數(shù)據(jù)收集
通過對資料的收集及群落類型的實地考察,選擇西部季風(fēng)常綠闊葉林皆伐后不同自然恢復(fù)時期(恢復(fù)15a、30a及老齡林)群落設(shè)置調(diào)查樣地。樣地面積為30m×30m,每個類型3個重復(fù),共調(diào)查樣地9塊。利用網(wǎng)格法將樣地分成36個5m×5m 的小樣方,對小樣方內(nèi)所有樹高大于1.3m 的木本植物進行每木調(diào)查。調(diào)查內(nèi)容包括物種名稱、胸徑、樹高及冠幅等。普洱季風(fēng)常綠闊葉林以短刺栲、刺栲、紅木荷為優(yōu)勢物種。因此,在調(diào)查過程中,對以上3種物種進行每木定位。此外,按照機械布點法選擇18個樣方,在每個樣方中心設(shè)置2m×2m 的小樣方進行幼苗調(diào)查及定位。植被調(diào)查的同時,記錄樣地所處生境特征,包括地理位置、海拔、坡度、坡向、坡位、林分郁閉度等。
1.3 數(shù)據(jù)處理
以樣地內(nèi)短刺栲、刺栲、紅木荷個體定位數(shù)據(jù)為依據(jù),應(yīng)用相鄰格子法,分5m×5m、5m×10m、10 m×10m、10m×20m、20m×20m 的樣方格子分別進行統(tǒng)計分析。測定種群空間分布格局的方法很多,本研究采用聚集度指標(biāo)進行測定。具體方法及參數(shù)意義見文獻[22~25]。
2 結(jié)果與分析
2.1 種群實際分布格局
根據(jù)樣地調(diào)查所得數(shù)據(jù),繪制了刺栲、短刺栲、紅木荷的實際分布散點圖(圖1~圖3)。由圖1可見,刺栲種群在恢復(fù)15a群落中僅在2個樣地中出現(xiàn),并且個體數(shù)量相對較少,而在恢復(fù)30a群落中則個體數(shù)量較多,老齡林也有2個樣地中個體數(shù)量相對較少。對于短刺栲種群,除老齡林中一個樣地外,其余樣地中個體數(shù)量均較多,散布于樣地中的絕大多數(shù)地方,并在局部呈現(xiàn)密集分布(圖2)。紅木荷在所有的調(diào)查樣地中均出現(xiàn),但個體數(shù)量少于短刺栲,其實際分布狀態(tài)則如圖3所示。
2.2 不同恢復(fù)階段3種優(yōu)勢物種空間分布格局
利用格子樣方法對季風(fēng)常綠闊葉林優(yōu)勢種群刺栲、短刺栲、紅木荷種群空間分布格局進行了理論擬合和格局強度判定(表1)。從表1 可以看出,在3個恢復(fù)階段,刺栲、短刺栲及紅木荷3種優(yōu)勢種均呈聚集分布,說明聚集分布是刺栲、短刺栲、紅木荷3種優(yōu)勢種群在季風(fēng)常綠闊葉林空間分布的基本狀態(tài)屬性。
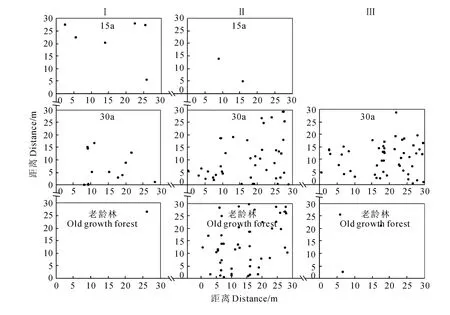
圖1 刺栲種群的實際分布狀態(tài)Fig.1 Spatial distribution status on spots of C.hystrixpopulation
2.3 不同恢復(fù)階段種群空間分布格局隨尺度的變化
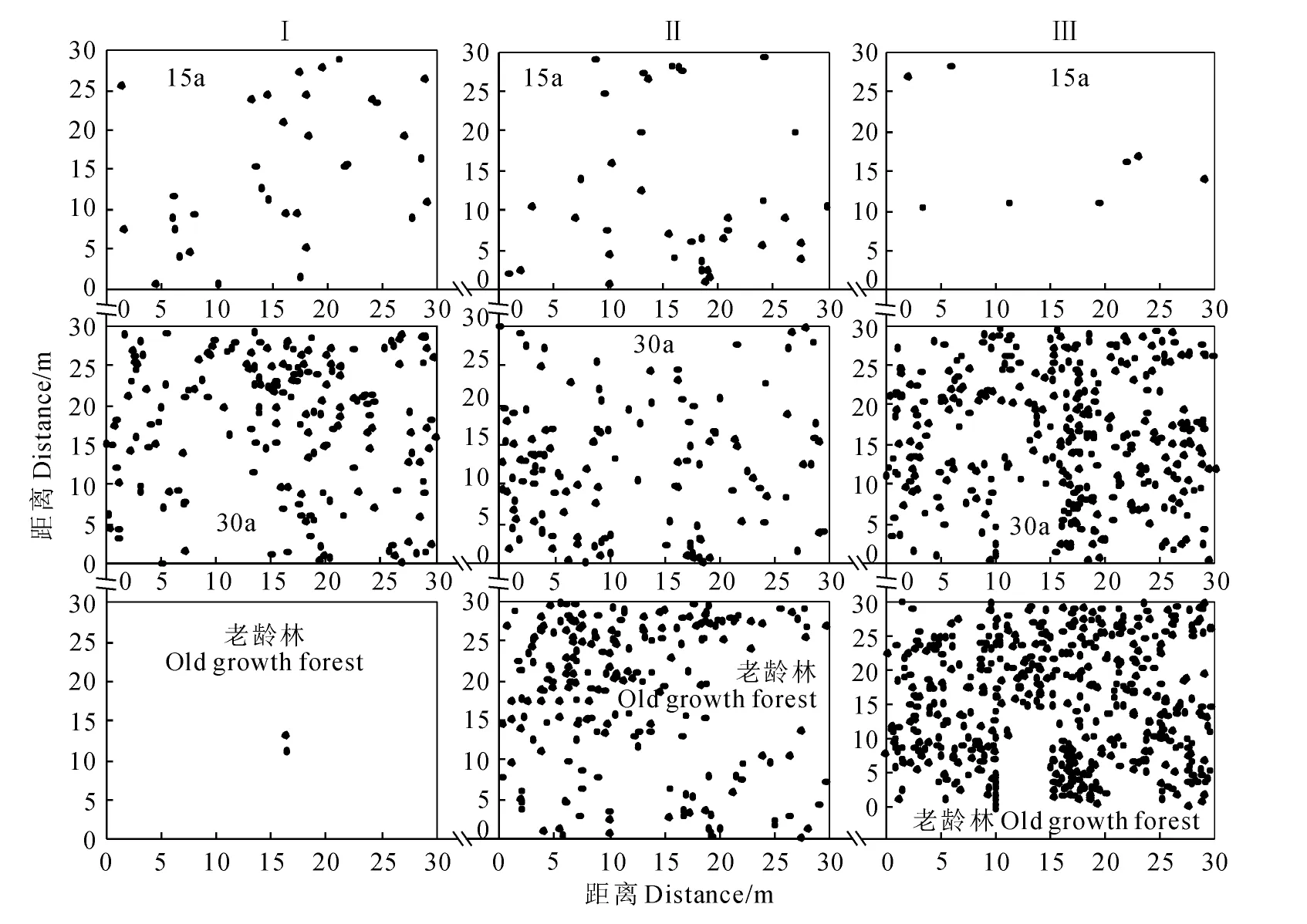
圖2 短刺栲種群的實際分布狀態(tài)Fig.2 Spatial distribution status on spots of C.echidnocarpa population
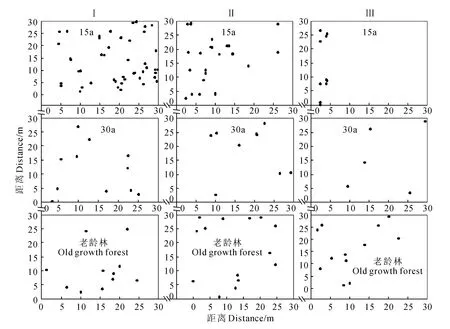
圖3 紅木荷種群的實際分布狀態(tài)Fig.3 Spatial distribution status on spots of S.wallichii population
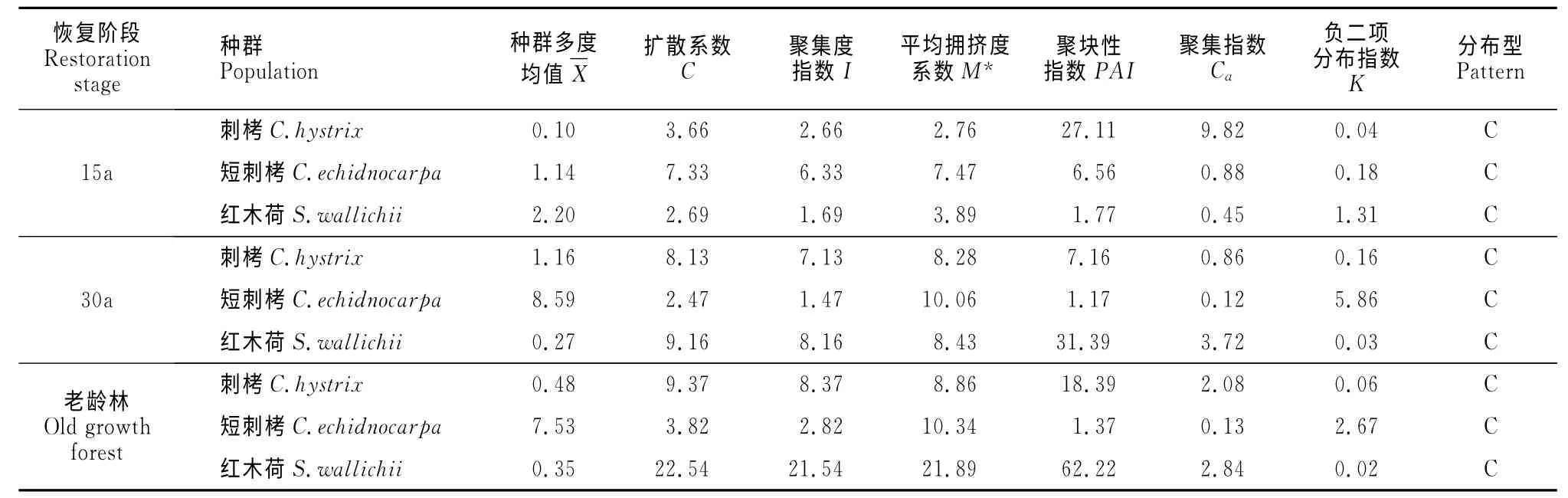
表1 不同恢復(fù)階段3種優(yōu)勢物種種群分布格局Table 1 Spatial distribution patterns of three dominant species at different restoration stages
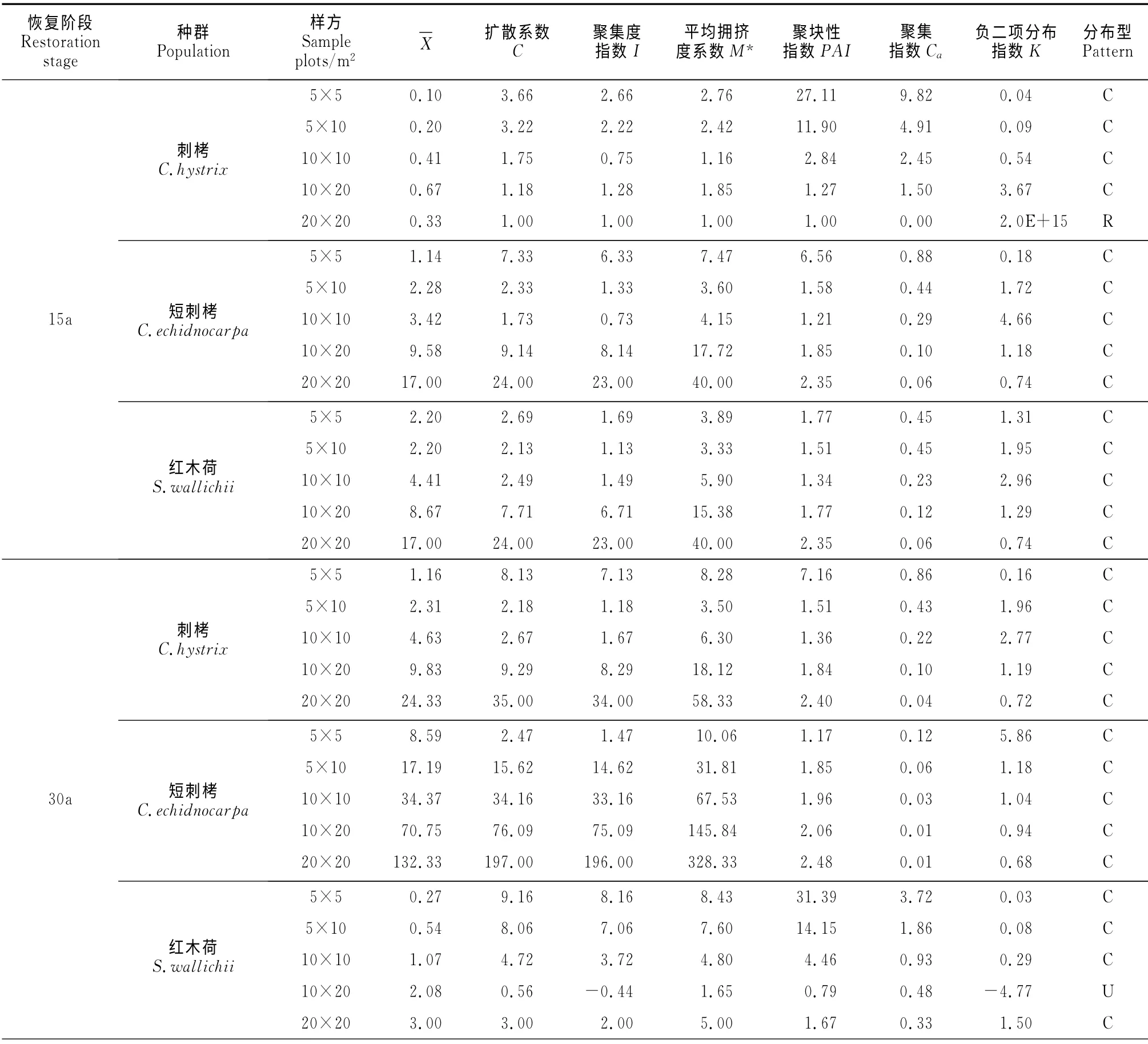
表2 不同取樣面積3種優(yōu)勢物種種群分布格局比較Table 2 Spatial distribution patterns of three dominant species in different sample areas
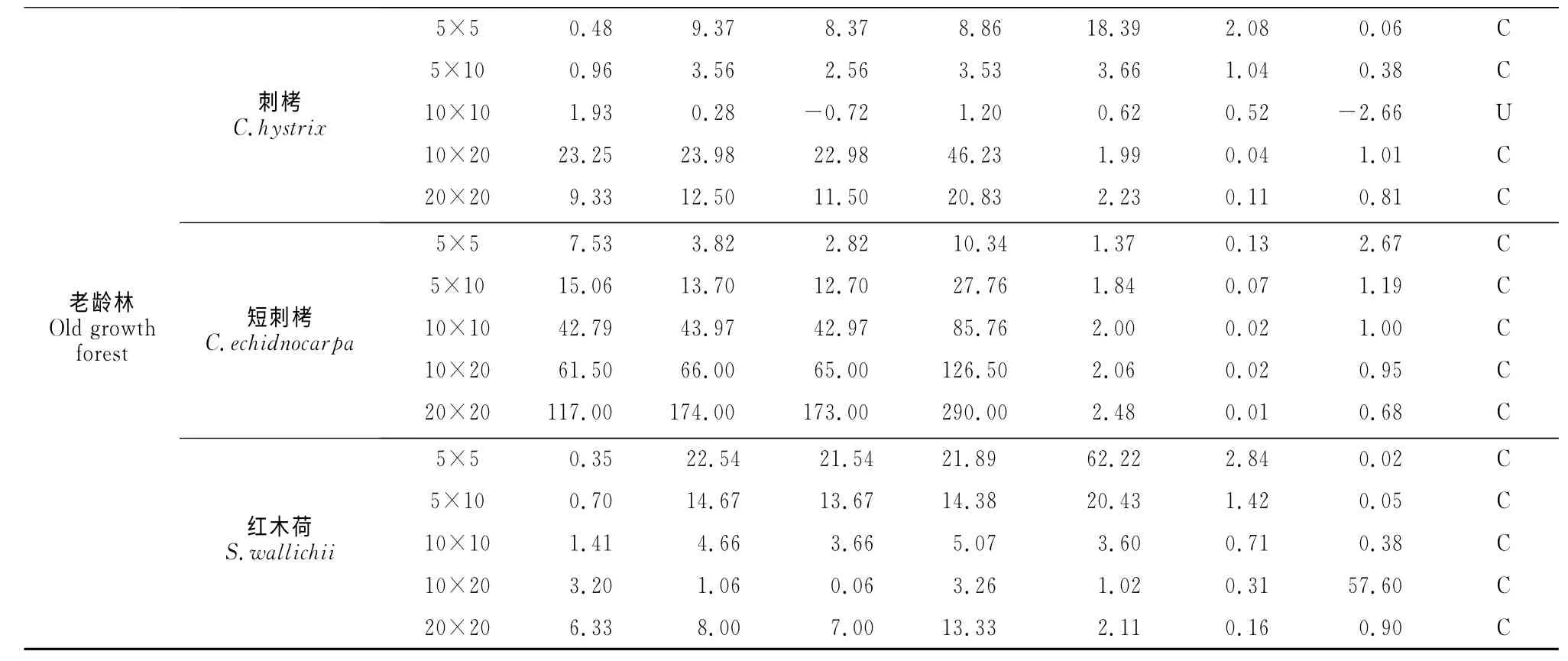
續(xù)表2 Continued Table 2
利用可變尺度相鄰格子樣方法對季風(fēng)常綠闊葉林3種優(yōu)勢物種種群空間分布格局進行理論擬合和格局強度判定(表2)。在所有恢復(fù)階段和尺度下,除刺栲在恢復(fù)15年20m×20m 尺度為隨機分布、原始林10m×10m尺度下為均勻分布、紅木荷在10m×20m 尺度下為均勻分布外,3個物種均為聚集分布(表2)。刺栲和紅木荷在所有群落中聚集強度均隨取樣尺度的增大而變小,而短刺栲在恢復(fù)15年群落中聚集強度均隨取樣尺度的增大而變小,但在恢復(fù)30年及老齡林群落中聚集強度則隨取樣尺度的增大而增大。在相同尺度下,刺栲隨群落恢復(fù),其聚集強度減弱,而短刺栲和紅木荷的聚集強度則隨群落恢復(fù)逐漸增大(表2)。
3 討 論
物種空間分布格局是物種與環(huán)境長期相互適應(yīng)、相互作用的結(jié)果,它不僅決定于物種自身的形態(tài)結(jié)構(gòu)、生理生態(tài)特性,同時也與自然生境條件及其所處群落中其他種群的競爭排斥等生態(tài)效應(yīng)密切相關(guān)[25-26]。不同恢復(fù)階段季風(fēng)常綠闊葉林3種優(yōu)勢物種(刺栲、短刺栲和紅木荷)均呈聚集分布,同時,在不同尺度下3種物種種群空間分布格局也呈現(xiàn)聚集分布。
物種的空間分布格局受多種因素影響,如環(huán)境因素、物種本身生物特性、生態(tài)學(xué)特性、形態(tài)結(jié)構(gòu)、生理特性等等[23,27-30]。然而,由于同一植物種的個體對環(huán)境等條件的要求基本上是一致的,自然環(huán)境條件對格局形成的作用較小,因此,種群生物學(xué)特征在形成物種空間格局過程中發(fā)揮了重要作用[31]。本研究中,刺栲、短刺栲、紅木荷3種物種聚集分布的空間分布格局形式與物種自身的生物學(xué)特性密切相關(guān)。首先,3種物種種子特征是其聚集分布的重要原因之一。作為殼斗科的刺栲和短刺栲,其果實為堅果,而山茶科的紅木荷果實則為蒴果,較重的果實決定了其種子傳播方式為重力傳播。因此,3 種物種種子成熟后,均落于母樹周圍,除少量由于動物取食被帶走外,絕大多數(shù)種子將在母樹周圍發(fā)芽,形成幼苗,從而導(dǎo)致3 種物種個體呈現(xiàn)聚集分布狀態(tài)。其次,3種物種較強的萌生能力也是其個體呈現(xiàn)聚集分布的重要原因。研究表明,3種物種均具有較強的萌生能力,并且萌生方式主要為根萌生和干基萌生,萌生更新是其主要的更新方式之一[15]。由于萌生并沒有改變個體的原有地理位置,同時,盡管萌生個體間存在較強的競爭和自疏作用,但其保留個體數(shù)量仍較多,從而導(dǎo)致個體分布更加密集,其聚集程度更高。可見,植物物種生物學(xué)特性在物種個體分布中具有重要作用。
此外,本研究結(jié)果也發(fā)現(xiàn),群落的恢復(fù)時間并沒有改變物種的分布形式。在所有的3種群落中,刺栲、短刺栲、紅木荷均為聚集分布。這與3種物種的萌生更新具有密切關(guān)系。由于本研究所選的干擾方式為皆伐,而皆伐留下的伐樁,為物種萌生留下了基礎(chǔ)。皆伐后,3種物種伐樁或根部芽原基通過內(nèi)部激素調(diào)整,吸收根部供應(yīng)的養(yǎng)分迅速萌發(fā),形成萌枝,占領(lǐng)采伐跡地。由于萌生植株具有明顯的競爭優(yōu)勢[32],其生長比實生植株更快,能迅速占據(jù)生境、快速恢復(fù)有性繁殖。因此,在恢復(fù)的植被中,其植被更新方式以萌生更新為主。正是皆伐后萌生的這種更新方式,導(dǎo)致了3種物種個體分布格局沒有發(fā)生變化,仍為聚集分布。
由于種群個體數(shù)、所處生境條件的異質(zhì)性以及種內(nèi)、種間關(guān)系的變化等因素均可引起種群分布格局產(chǎn)生相應(yīng)的變化。所以,種群空間分布格局具有尺度依賴性[33]。在研究種群空間分布格局時人為地確定某個單位尺度進行調(diào)查,所得結(jié)果只能代表那個尺度上的格局強度[34]。因此,種群分布格局的研究不應(yīng)該在單一隨機性取樣尺度上進行,集群性發(fā)生的真正尺度只能通過對同一種群進行多次不同樣方大小的反復(fù)取樣來判定[35]。由于本研究過程中將研究對象(刺栲、短刺栲、紅木荷)每一個個體都進行了定位,所以可按任意大小進行取樣,這樣便可以考察任意尺度上的3 種物種種群的空間分布狀況,這樣格局強度和格局規(guī)模的判斷更具有真實性。而本文的研究結(jié)果發(fā)現(xiàn),取樣尺度并沒有改變物種的分布格局,3種物種在3種取樣面積下均為聚集分布(除刺栲在恢復(fù)15年20m×20m、原始林10 m×10m 尺度、紅木荷在10m×20m 尺度下外)。這說明聚集分布格局是刺栲、短刺栲和紅木荷3種種群空間分布的基本屬性。種群的聚集型分布,是生存與發(fā)展的一種策略。聚集分布使種內(nèi)競爭大于種間競爭,激烈的種內(nèi)競爭可以淘汰一些較弱的個體,增強剩余個體的競爭能力,有利于種群的進化;另一方面,剩余個體可以利用充裕的空間和資源,有利于種群的健康發(fā)展。
總之,刺栲、短刺栲、紅木荷在本文研究的恢復(fù)階段及尺度下,其個體分布均為聚集分布,物種生物學(xué)特性在其個體分布方式上起到了決定性作用。
[1]YANG H(楊 慧),LOU A R(婁 安 如),GAO Y J(高 益 軍),etal.Life history characteristics and spatial distribution of theBetula platyphyllapopulation in the Dongling Mountain region in Beijing[J].JournalofPlantEcology(Chinese Version)(植物生態(tài)學(xué)報),2007,31(2):272-282(in Chinese).
[2]HE F,LEGENDRE P,LAFRANKIE J V.Distribution patterns of tree species in a Malaysian tropical rain forest[J].JournalofVegetationScience,1997,8(1):105-114.
[3]DRUCKENBROD D L,SHUGART H H,DAVIES I.Spatial pattern and process in forest stands within the Virginia piedmont[J].Journal ofVegetationScience,2005,16(1):37-48.
[4]NATHAN R.Long-distance dispersal of plants[J].Science,2006,313(5 788):786-788.
[5]LI L(李 立),CHEN J H(陳建華),REN H B(任海保),etal.Spatial patterns ofCastanopsiseyreiandSchimasuperbain mid-subtropical broadleaved evergreen forest in Gutianshan National Reserve,China[J].ChineseJournalofPlantEcology(植物生態(tài)學(xué)報),2010,34(3):241-252(in Chinese).
[6]CONDIT R,ASHTON P S,BAKER P,etal.Spatial patterns in the distribution of tropical tree species[J].Science,2000,288(5 470):1 414-1 418.
[7]MIAO N(繆 寧),LIU SH R(劉世榮),SHI Z M(史作民),etal.Spatial patterns of dominant tree species in sub-alpineBetulaabiesforest in West Sichuan of China[J].ChineseJournalofAppliedEcology(應(yīng)用生態(tài)學(xué)報),2009,20(6):1 263-1 270(in Chinese).
[8]WEI Q SH(尉秋實),WANG J H(王繼和),LI CH L(李昌龍),etal.A preliminary study on the distribution patterns and characeristics ofAmmopiptanthusmongolicuspopulations in different desert environments[J].JournalofPlantEcology(Chinese Version)(植物生態(tài)學(xué)報),2006,29(4):591-598(in Chinese).
[9]LIU W D(劉萬德),SU J R(蘇建榮),LI SH F(李帥鋒),etal.Species-area relationship at different succession stages of monsoon evergreen broad-leaved forest in south subtropical area of Yunnan Province[J].ChineseJournalofAppliedEcology(應(yīng)用生態(tài)學(xué)報),2011,22(2):317-322(in Chinese).
[10]LI SH F(李帥鋒),LIU W D(劉萬德),SU J R(蘇建榮),etal.Niches and interspecific associations of dominant tree populations at different restoration stages of monsoonal broad-leaved evergreen frorest[J].ChineseJournalofEcology(生態(tài)學(xué)雜志),2011,30(3):508-515(in Chinese).
[11]LI SH F(李帥鋒),LIU W D(劉萬德),SU J R(蘇建榮),etal.Woody seedling regeneration in secondary succession of monsoon broad-leaved evergreen forest in Puer,Yunnan,Southwest China[J].ActaEcologicaSinica(生態(tài)學(xué)報),2012,32(18):5 653-5 662(in Chinese).
[12]LI SH F(李帥鋒),SU J R(蘇建榮),LIU W D(劉萬德),etal.Deversity and distribution of vascular epiphytes in different restoration stages of monsoonal broad-leaved evergreen forest communities[J].ForestResearch(林業(yè)科學(xué)研究),2011,24(2):151-158(in Chinese).
[13]LIU W D(劉萬德),SU J R(蘇建榮),ZHANG ZH J(張志鈞),etal.Effect of rstoration strategy and time on community characteristic of monsoon evergreen broad-leaved forest[J].ForestResearch(林業(yè)科學(xué)研究),2011,24(1):1-7(in Chinese).
[14]LIU W D(劉萬德),SU J R(蘇建榮),LI SH F(李帥鋒),etal.Stoichiometry study of C,N and P inplant and soil at different successional stages of monsoon broad-leaved evergreen forest in Pu’er,Yunnan Province[J].ActaEcologicaSinica(生態(tài)學(xué)報),2010,30(23):6 581-6 590(in Chinese).
[15]SU J R(蘇建榮),LIU W D(劉萬德),ZHANG ZH J(張志鈞),etal.Sprouting characteristic in restoration ecosystems of monsoon evergreen broad-leaved forest in south-central of Yunnan Province[J].ActaEcologicaSinica(生態(tài)學(xué)報),2012,32(3):805-814(in Chinese).
[16]LIU W P(劉文平),CAO H L(曹洪麟),LIU W(劉 衛(wèi)),etal.Study on diversity of monsoon evergreen broad leaved forest in different kinds of habitat in Dinghushan[J].JournalofAnhuiAgriculturalSciences(安徽農(nóng)業(yè)科學(xué)),2011,39(26):16 159-16 163(in Chinese).
[17]ZHOU G Y(周國逸),LUO Y(羅 艷),OUYANG X J(歐陽學(xué)軍),etal.Analysis on the concentration of some sediment elements in their transport process through monsoon evergreen broad-leaved forest ecosystem in Dinghushan,Guangdong Province,China[J].Acta EcologicaSinica(生態(tài)學(xué)報),2003,23(7):1 408-1 414(in Chinese).
[18]ZHOU X Y(周小勇),HUANG ZH L(黃忠良),OUYANG X J(歐陽學(xué)軍),etal.Succession of the originalCastanopsischinensis-Cryptocaryachinensis-Schimasuperbacommunity of monsoon evergreen broad-leaved forest in Dinghushan nature reserve[J].ActaEcologicaSinica(生態(tài)學(xué)報),2005,25(1):37-44(in Chinese).
[19]SONG J(宋 娟),LI R H(李榮華),ZHU SH D(朱師丹),etal.Leaf functional traits of ferns from different habitats in monsoon evergreen broad-leaved forest in Dinghushan Mountain[J].JournalofTropicalandSubtropicalBotany(熱帶亞熱帶植物學(xué)報),2013,21(6):489-495(in Chinese).
[20]ZHANG N(張 娜),QIAO Y N(喬玉娜),LIU X ZH(劉興詔),etal.Nutrient characteristics in incident rainfall,throughfall,and stemflow in monsoon evergreen broad-leaved forest at Dinghushan[J].JournalofTropicalandSubtropicalBotany(熱帶亞熱帶植物學(xué)報),2010,18(5):502-510(in Chinese).
[21]ZHANG CH(張 池),HUANG ZH L(黃忠良),SHI J H(史軍輝),etal.Dynamics and causes of woody plant death in the monsoon evergreen broad-leaved forest in Dinghushan nature reserve[J].ActaEcologicaSinica(生態(tài)學(xué)報),2006,26(8):2 457-2 462(in Chinese).
[22]WEI X Z(魏新增),HUANG H D(黃漢東),JIANG M X(江明喜),etal.Quantitative characteristics and spatial distribution patterns of Euptelea pleiospermum populations in riparian zones of the Shennongjia area,central China[J].JournalofPlantEcology(Chinese Version)(植物生態(tài)學(xué)報),2008,32(4):825-837(in Chinese).
[23]ZHANG W H(張文輝),WANG Y P(王延平),KANG Y X(康永祥),etal.Spatial distribution pattern ofLarixchinensispopulation in Taibai Mt.[J].ChineseJournalofAppliedEcology(應(yīng)用生態(tài)學(xué)報),2005,16(2):207-212(in Chinese).
[24]FAN F R(范繁榮),PAN B ZH 潘標(biāo)志),MA X Q(馬祥慶),etal.A study on structure and spatial distribution pattern ofArtocarpus hypargyreuspopulation[J].ForestResearch(林業(yè)科學(xué)研究),2008,21(2):176-181(in Chinese).
[25]LIU W D(劉萬德),SU J R(蘇建榮),WANG M J(王夢君),etal.Spatial distribution pattern ofTaxusyunnanensispopulation in northwest Yunnan Province[J].JournalofNorthwestForestryUniversity(西北林學(xué)院學(xué)報),2012,27(6):131-138(in Chinese).
[26]HAO CH Y(郝朝運),ZHANG X P(張小平),LI W L(李文良),etal.Spatial distribution ofMonimopetalumchinensepopulations in different forest types[J].ActaEcologicaSinica(生態(tài)學(xué)報),2008,28(6):2 900-2 908(in Chinese).
[27]LAN G Y(蘭國玉),HU Y H(胡躍華),CAO M(曹 敏),etal.Establishment of Xishuangbanna tropical forest dynamics plot:species compositions and spatial distribution pattern[J].JournalofPlantEcology(Chinese Version)(植物生態(tài)學(xué)報),2008,32(2):287-298(in Chinese).
[28]SHEN L(沈 林),YANG H(楊 華),KANG X G(亢新剛),etal.Effects of selective cutting intensity on spatial distribution pattern of natural spruce-fir forests[J].JournalofCentralSouthUniversityofForestry&Technology(中南林業(yè)科技大學(xué)學(xué)報),2013,(1):68-74(in Chinese).
[29]ZHAO L Q(趙麗瓊),HUANG H G(黃華國),LIANG D SH(梁大雙),etal.Spatial distribution pattern ofPiceacrassifoliapopulation in Dayekou,Gansu Province[J].JournalofBeijingForestryUniversity(北京林業(yè)大學(xué)學(xué)報),2010,32(4):59-64(in Chinese).
[30]ZHAO CH M(趙常明),CHEN Q H(陳慶恒),QIAO Y K(喬永康),etal.Structure and spatial pattern of a naturalAbiesfaxonianapopulation on the eastern edge of Qinghai-Tibetan plateau[J].JournalofPlantEcology(Chinese Version)(植物生態(tài) 學(xué)報),2004,28(3):341-350(in Chinese).
[31]NIU L L(牛麗麗),YU X X(余新曉),LIU SH Y(劉淑燕),etal.Life history characteristics and spatial distribution ofPinustabulaeformispopulation in Songshan Nature Reserve of Beijing[J].JournalofBeijingForestryUniversity(北京林業(yè)大學(xué)學(xué)報),2008,30(s2):17-21(in Chinese).
[32]VIEIRA D L M,SCARIOT A.Principles of natural regeneration of tropical dry forests for restoration[J].RestorationEcology,2006,14(1):11-20.
[33]WANG B Y(王本洋),YU SH X(余世孝).Multi-scale analyses of population distribution patterns[J].ActaPhytoecologicaSinica(植物生態(tài)學(xué)報),2005,29(2):235-241(in Chinese).
[34]LI M H(李明輝),HE F H(何風(fēng)華),LIU Y(劉 云),etal.Spatial distribution pattern of tree individuals in the schrek spruce forest,northwest China[J].ActaEcologicaSinica(生態(tài)學(xué)報),2005,25(5):1 000-1 006(in Chinese).
[35]XIE Z Q(謝宗強),CHEN W L(陳偉烈),LIU ZH Y(劉正宇),etal.Spatial distribution pattern ofCathayaargyrophyllapopulation[J].BulletinofBotany(植物學(xué)報),1999,41(1):95-101(in Chinese).
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學(xué)生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學(xué)苑創(chuàng)造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(shù)(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
