根皮素對肝癌細胞HepG-2增殖和凋亡的影響
2015-07-07 15:16:05王會劉政吳漢東
中國生化藥物雜志 2015年7期
王會,劉政,吳漢東
(遼寧醫學院 食品科學與工程學院,遼寧 錦州 121001)
?
根皮素對肝癌細胞HepG-2增殖和凋亡的影響
王會,劉政,吳漢東
(遼寧醫學院 食品科學與工程學院,遼寧 錦州 121001)
目的 以人肝癌細胞HepG-2為研究對象,研究根皮素對肝癌細胞增殖和凋亡的影響。方法 應用相差顯微鏡、電子顯微鏡檢測凋亡細胞形態學變化,通過細胞毒性試驗檢測細胞增殖情況,應用流式細胞儀檢測細胞凋亡率、細胞周期、線粒體膜電位、鈣離子穩態的變化。結果 細胞呈典型的凋亡形態學變化,根皮素對HepG-2細胞有抑制增殖作用和誘導凋亡作用且呈濃度和時間依賴性,細胞周期阻滯于G1期,細胞線粒體膜電位降低,細胞內鈣離子濃度增大。結論 根皮素通過影響細胞周期,降低線粒體膜電位,改變細胞內鈣離子平衡來誘導肝癌細胞HepG-2凋亡。
根皮素;肝癌細胞;細胞凋亡;增殖
肝癌是發生在肝細胞或膽管細胞的世界第五大惡性腫瘤,具有診斷難,治愈難,發展快和預后差等特點。肝癌與其他惡性腫瘤相比具有較強的耐藥性,因此尋找高效、低毒、靶點明確的抗肝癌藥物迫在眉睫。
根皮素是一種存在于蘋果、梨等水果和多種蔬菜根莖或根皮中的類黃酮物質,它具有抗菌、抗氧化、抗糖尿病、抗腫瘤及雌激素樣作用等活性。根皮素能激活蜂窩蛋白激酶,對細胞無序增殖有抑制作用,可用于皮膚癌及其他腫瘤的治療[1-2]。根皮素對體外生長的人急性淋巴母細胞白血病細胞Molt-4 和體內膀朧癌細胞Fisher具有抑制增殖作用。根皮素主要通過抑制糖的跨膜運輸,阻礙能源的供應來誘導 B16 小鼠腫瘤 4A5 細胞凋亡[3],根皮素還可以嵌入到B16 腫瘤細胞DNA分子中干擾 DNA 的復制,影響蛋白激酶C (protein kinase C, PKC) PKC 的活性,從而促進細胞凋亡[4]。Park[5]等研究發現根皮素可誘導腸癌細胞凋亡7。但目前關于根皮素誘導肝癌細胞凋亡及其相關機理的詳細研究還非常少。本實驗以人肝癌細胞 HepG-2 為研究對象,探討了根皮素對其增殖和凋亡的影響。
1 材料與方法
1.1 材料 人肝癌HepG-2細胞株,由協和醫科大學提供;根皮素(純度, 99.99%;批號,P7912-100MG)、二甲基亞砜(DMSO)、甲基噻唑藍(MTT)、碘化丙啶(PI)、均購于Sigma公司;DMEM培養基和胎牛血清購于GIBCO公司;Annexin V-FITC Apoptosis Detection試劑盒購于BD公司。細胞凋亡-Hoechst染色試劑盒購于碧云天公司。
流式細胞儀(BECKMAN FC500);激光共聚焦顯微鏡(Nikon TE2000-E);倒置相差顯微鏡(Olympus IX-71)。酶標儀(THERMO FISHER),透射電子顯微(JEM-123O)
1.2 細胞培養 用含10% 標準胎牛血清的DMEM培養基,于37 ℃、5% CO2及飽和濕度的培養箱中培養細胞,當培養液顏色變黃時更換培養液,當細胞達到70%~80% 融合度時細胞傳代。
1.3 方法
1.3.1 藥物配制及細胞處理 根皮素用DMSO溶解,再用基礎培養基稀釋成濃度為8 mg/mL的母液,分裝,置-20 ℃ 冰箱避光保存備用。在實驗中DMSO的終濃度為0.05%。取對數生長期細胞,換用含有40、80和160 mg/L根皮素的培養液培養至12、24、36、48 h。分別記為低、中、高濃度處理組。設對照組(用DMSO處理,終濃度為0.05%)。
1.3.2 細胞形態學觀察 細胞懸液接種于六孔板,濃度為4.0×105個/孔,待細胞貼壁融合進入對數生長期,用低、中、高濃度處理組根皮素處理,用培養液培養至24 h,并設對照組(用DMSO處理,終濃度為0.05%)。應用倒置相差顯微鏡觀察并拍照。
1.3.3 Hoechst 33258染色觀察凋亡細胞核 將蓋玻片置于培養皿內,種入細胞。待細胞進入對數用低、中、高濃度處理組處理,設置對照組(用DMSO處理,終濃度為0.05%)。固定細胞,加入Hoechst 33258染色液,去染色液,滴抗熒光淬滅封片液于載玻片上,蓋蓋玻片,在激光共聚焦顯微鏡下觀察并拍照。
1.3.4 細胞毒性分析 將180 μL細胞懸液接種于96孔培養板。待細胞進入對數生長期,低、中、高濃度處理組處理,設置對照組,每組設 8個重復。培養到設定時間,然后每孔加入 MTT 20 μL(5 mg/mL, PBS溶解),4 h 后終止孵育,離心去上清,每孔加入 200 μL DMSO,混勻,于酶標儀在 490 nm 波長處檢測吸光度 OD值。試驗重復3次。
1.3.5 透射電鏡觀察凋亡細胞亞顯微結構 常規方法收集細胞,用2.4% 戊二醛固定,0.15 M 磷酸緩沖液漂沖,1.2% 鋨酸固定,0.12M 磷酸緩沖液漂洗,用20%、30%、60%、70%、80% 和100% 梯度的丙酮系列脫水。用環氧樹脂SPURR包埋聚合,切片,醋酸雙氧鈾和檸檬酸鉛復染。透射電子顯微鏡觀察,拍攝。
1.3.6 Annexin V-FITC/PI雙染檢測細胞凋亡率 常規方法收集細胞,調整細胞濃度為5.0×105細胞/樣品,樣品加入100 μL緩沖液和5 μL AnnexinV-FITC及5 μL PI,搖勻。避光孵育10~15 min,流式細胞儀檢測。
1.3.7 細胞周期檢測 收集對照組和處理組細胞,調整細胞濃度為(1~5)×105個/樣品,于4 ℃,70% 無水乙醇中孵育12 h。離心去乙醇,PBS漂洗,加入0.5 mLPI染液,4 ℃ 避光孵育20 min。流式細胞儀上機檢測。
1.3.8 線粒體膜電位檢測 收集對照組和處理組細胞,1.0×106個/樣品,加入0.5 mL JC-1染色液,在CO2培養箱避光孵育15 min,用預熱的PBS洗滌,于 0.5 mL PBS懸浮,流式細胞儀上機檢測。
1.3.9 細胞內鈣離子濃度的測定 收集對照組和處理組細胞,調整細胞濃度至(1~2)×106個/mL。加入Fluo-3/Am至終濃度為6~10 μM,細胞于5% CO2,37 ℃,避光孵育20~40 min,期間輕輕振蕩幾次,設置陰性對照(不加Fluo-3/Am)。用無鈣離子PBS離心洗滌2次,然后 PBS重懸至0.5 mL,流式細胞儀上機檢測。

2 結果
2.1 細胞形態學觀察 對照組細胞排列緊密,融合成片,胞漿飽滿,呈橢圓形,邊緣清晰,細胞折光度好,增殖旺盛。經根皮素處理24 h后,低濃度處理組細胞萎縮,出現空泡;中濃度處理組細胞增殖速度顯著減慢,細胞形態逐漸變的模糊不清,凋亡細胞與鄰近細胞脫離;高濃度處理組細胞間連接幾乎消失,細胞裂解成小的碎片,細胞膜皺縮,胞體縮小,細胞膜表面凹凸不平,細胞形態發生重大變化,出現細胞壞死(見圖1)。
2.2 Hoechst33258染色觀察凋亡細胞核 對照組細胞核呈卵圓形或圓形,染色質均勻分布于細胞核,著色均勻。處理組細胞染色質凝聚呈分散點狀,著色不均勻。隨根皮素濃度的增加點狀著色現象明顯(見圖2)。
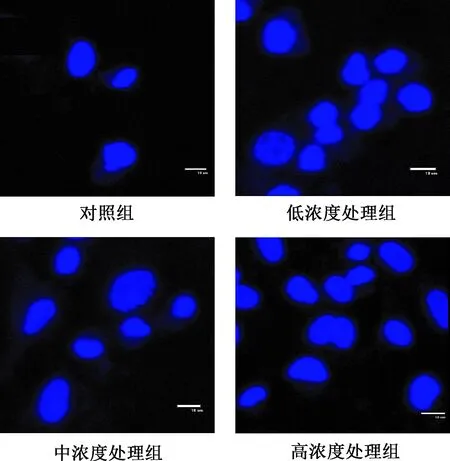
圖2 Hoechst33258 染色,根皮素處理HepG-2細胞24 h (×100)Fig.2 Hoechst33258 staining, HepG-2 cells after 24 h treatment by phloretin (×100)
2.3 細胞毒性分析 細胞毒性結果見表1,OD490值反映的是活細胞數量。在根皮素處理HepG-2細胞12、24、36、48 h后,隨著根皮素濃度的增大其OD490值隨之減小,間接反映活細胞數量減小,呈良好的量效和時效關系。
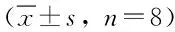
表1 根皮素對HepG-2細胞生長的影響Tab.1 The inhibition effect of phloretin on HepG-2 cell ±s, n=8)
*P<0.05,**P<0.01,與對照組比較, compared with control group
2.4 透射電鏡觀察凋亡細胞亞顯微結構 對照組細胞胞質均勻,核膜結構完好,呈雙層膜結構。核仁清晰可見,染色質分布均勻。高濃度處理組細胞,雙層核膜膜結構遭到損傷、裂解,染色質濃縮、聚集(見圖4)。
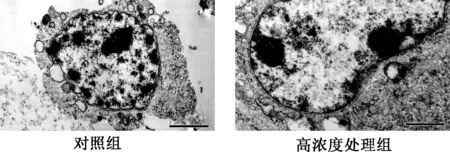
圖4 電子顯微鏡觀察根皮素處理HepG-2細胞24 h亞細胞結構(×100)Fig.4 Subcellular observation of HepG-2 cells after 24h treatment by phloretin using transmission electron microscopy(×100)
2.5 細胞凋亡率的測定 Annexin V-FITC/PI雙標記法能夠定量分析細胞凋亡,并可定量分析早期凋亡細胞、晚期凋亡細胞、活細胞和壞死細胞。實驗結果如圖5(12、36、48 h凋亡率),表2結果表明,隨著根皮素處理時間的延長和濃度的增加,誘導凋亡效果越顯著,呈現濃度依賴性和時間依賴性。

圖5 根皮素作用HepG-2細胞凋亡率Fig.5 The apoptosis rate of HepG-2 after treatment by phloretin.
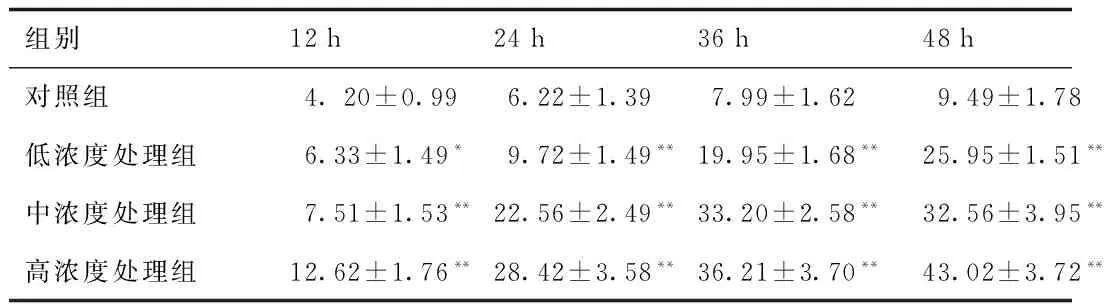
組別 12h24h36h48h對照組 4.20±0.996.22±1.397.99±1.629.49±1.78低濃度處理組6.33±1.49*9.72±1.49**19.95±1.68**25.95±1.51**中濃度處理組7.51±1.53**22.56±2.49**33.20±2.58**32.56±3.95**高濃度處理組12.62±1.76**28.42±3.58**36.21±3.70**43.02±3.72**
*P<0.05,**P<0.01,與對照組比較, compared with control group
2.6 細胞周期的檢測 采用PI標記結合流式細胞術檢測細胞周期的變化,結果如圖6。隨著根皮素濃度的增大,G1期細胞數百分比增大,S期細胞百分比下降,G2期細胞百分比下降,表明細胞經根皮素處理后增殖停滯在 G1期,DNA合成受到抑制,這種關系隨著根皮素劑量的增大而愈加明顯,呈現劑量依賴性。
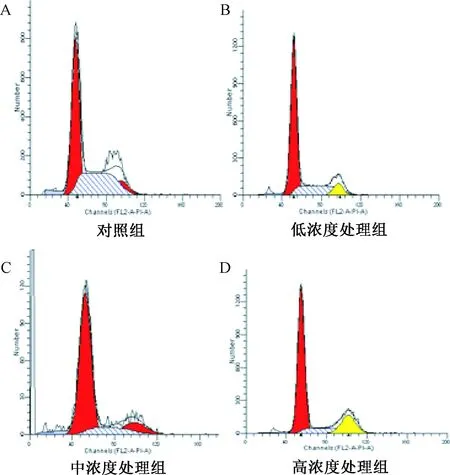
圖6 根皮素處理HepG-2細胞24 h細胞周期Fig.6 The cell cycle of HepG-2 cells after 24h treatment by phloretin
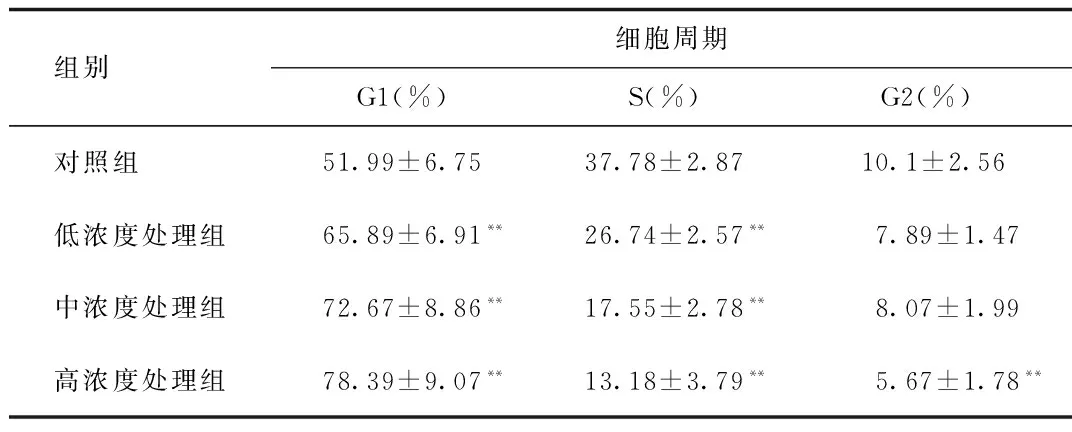
組別 細胞周期G1(%)S(%)G2(%)對照組 51.99±6.7537.78±2.8710.1±2.56低濃度處理組65.89±6.91**26.74±2.57**7.89±1.47中濃度處理組72.67±8.86**17.55±2.78**8.07±1.99高濃度處理組78.39±9.07**13.18±3.79**5.67±1.78**
*P<0.05,**P<0.01,與對照組比較, compared with control group
2.7 細胞線粒體膜電位的檢測 本實驗采用JC-1標記流式細胞儀檢測線粒體膜電位變化,發現經根皮素作用后的HepG-2細胞的線粒體膜電位下降,對照組為(10.05±1.03)mV,在濃度處理組為(5.30±0.72)mV,中濃度處理組為(3.77±8.92)mV,高濃度處理組為(2.04±0.78)mV,各實驗組分別與對照組比較差異極顯著(P<0.01,見圖7)。

圖7 根皮素作用HepG-2細胞24 h,線粒體膜電位變化情況,**P<0.01,與對照組相比Fig.7 The mitochondrial membrane potential of HepG-2 cells after 24 h treatment by phloretin,**P<0.01, compared with control group
2.8 細胞內鈣離子濃度的測定 圖 8反映了根皮素素作用HepG-2細胞24 h后Ca2+的濃度。對照組為(2.11±0.72)mmol/L,低濃度處理組為(3.79±0.75)mmol/L,中濃度處理組為(11.36±0.67)mmol/L,高濃度處理組為(20.98±0.76)mmol/L。隨根皮素濃度增大 Ca2+濃度呈現增大現象,處理組 Ca2+濃度較對照有明顯增多(P<0.01)。可見 Ca2+的濃度與根皮素濃度成正比。
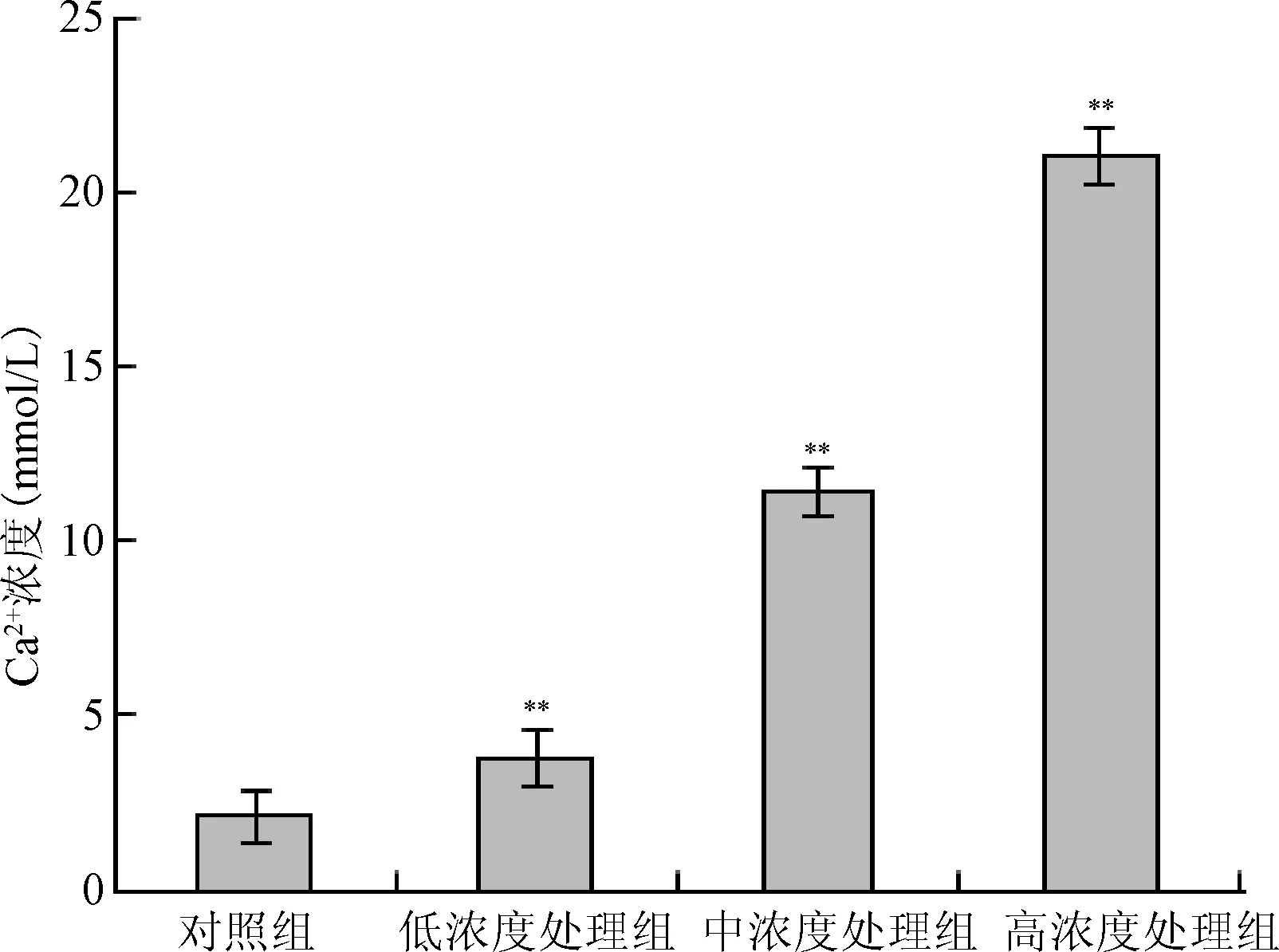
圖8 根皮素作用HepG-2細胞24 h Ca2+濃度變化,**P<0.01,與對照組相比Fig.8 The concentrations of intracellular calcium of HepG-2 cells after 24 h treatment by phloretin**P<0.01, compared with control group
3 討論
腫瘤的發生和發展是一個基因調控的復雜過程,是細胞增殖失控和凋亡受阻的最終結果。隨著人們對凋亡機理的研究,人們認識到促進腫瘤細胞凋亡是治療腫瘤的有效途徑之一。
根皮素是一種類黃酮類物質, 是一種具有臨床應用潛能的治療腫瘤的天然化合物, 根皮素可抑制腫瘤細胞增殖, 誘導腫瘤細胞凋亡[6]。根皮素可通過干擾Ⅱ型葡萄糖轉運蛋白來促進肝癌細胞凋亡[7]。根皮素能夠以線粒體途徑誘導 BEL-7402 細胞凋亡,且呈現濃度依賴性;根皮素能干預肝癌細胞粘附、運動及侵襲能力,從而抑制腫瘤細胞轉移[8-9]。由于根皮素結構中的 4 個羥基能夠抑制細胞中糖的跨膜轉運,阻斷能量運輸,因而具有誘導癌細胞凋亡的作用。
腫瘤是一種細胞周期失控性疾病,細胞無限增殖,導致細胞惡性轉化,最終形成腫瘤[10]。研究認為,細胞周期進程受到阻礙會誘導細胞凋亡,細胞周期的調節可以影響細胞分裂增殖又能影響細胞凋亡,許多凋亡誘導因素在影響細胞周期進程的同時也影響凋亡機制。所以對細胞周期的檢測是研究細胞凋亡的重要指標之一,通過干擾細胞周期進程來誘導凋亡也作為抗腫瘤研究的一個新思路。根皮素分子結構中具有 4 個羥基,與葡萄糖結構相似,與葡萄糖運輸形成非競爭性抑制,抑制能量來源,促使細胞發生凋亡。根皮素為芳香環結構可插入到 DNA雙螺旋分子中,從而阻止 DNA 合成,影響細胞周期的分布,細胞發生凋亡。
通過檢測根皮素作用的細胞的周期的分布情況,發現經根皮素作用的細胞發生G1期阻滯,導致G2期細胞減少,造成進入M期的細胞數量減少,無法分裂,使增殖受阻。隨著根皮素濃度的增加,G1期細胞增加,S期細胞減少。隨根皮素濃度增加,160 mg/L的根皮素誘導細胞大量凋亡,而造成 G2期細胞減少。推測根皮素通過調控細胞周期的進程進而發揮促凋亡作用,這為以后開展根皮素誘導肝癌細胞凋亡機理的研究提供了重要實驗依據。
線粒體與細胞凋亡過程中許多重要事件密切相關[11],發揮著重要的調控作用,線粒體膜電位下降,線粒體膜通透性轉運孔開放, 膜通透性增加,使線粒體內的細胞色素C,凋亡誘導因子 AIF 等進入胞漿,啟動凋亡程序[12]。根皮素是一種解偶聯試劑,可以抑制線粒體的氧化磷酸化作用[13],ATP合成受阻,能量缺乏,促使細胞凋亡。本實驗采用 JC-1標記結合流式細胞儀檢測線粒體膜電位變化,發現經根皮素作用后的HepG-2細胞的線粒體膜電位降低。線粒體膜電位降低,導致膜通透性增強,從而使細胞色素C 從線粒體內流入細胞質,細胞色素C 與信號接頭分子Apaf-1 (apoptotic protease activating factor-1) 結合,激活Caspase-9,活化 Caspase-3,啟動 Caspase 的級聯反應,引起細胞凋亡。推測根皮素誘導 HepG-2凋亡與細胞凋亡線粒體途徑有關。
在正常細胞中Ca2+主要與蛋白質結合封存在內質網和線粒體中,游離的 Ca2+很少,只是在收到外界刺激時才被釋放出來充當信號分子。幾乎所有細胞的反應,從收縮、胞吐到基因表達、細胞凋亡都被細胞質內局部或全部游離 Ca2+濃度的變化控制。有研究表明游離 Ca2+在細胞內的過度積累可以直接導致細胞凋亡的發生[14]。當根皮素誘導 HepG-2 細胞凋亡時,細胞質內游離鈣離子濃度升高,Ca2+穩態發生變化,表明適當濃度的根皮素有較好的誘導 HepG-2 細胞凋亡的效果。結合線粒體膜電位降低的結果推測與內質網上鈣離子的釋放有關,但其具體的機制有待進一步的研究。
根皮素是通過抑制細胞 DNA 合成,降低線粒體膜和干擾鈣離子穩態來誘導 HepG-2 細胞凋亡的。根皮素誘導肝癌細胞凋亡及其機制有待進一步研究。
[1] 譚飔,周志欽.根皮苷研究進展[J].食品與發酵工業,2013,39(8): 182-186.
[2] Setala Kai ME.Composition and method for rational treatment of cancer[P].USA: 4555806, 1986-01-21.
[3] Kobori M, I Washita k, Shinmoto H, et al. Phloretin-induced apoptosis in B16 melanoma 4A5 and HL60 human leukemia cells.[J]. Biosci Biotechnol Bioehem, 1999, 63(4):719-725.
[4] Kobori M, Shinmoto H, Tsushida T, et al. Phloretin-induced apoptosis in B16 melanoma 4A5 celll by inhibition of glucose transmembrane transport [J]. Cancer Lett, 1997,119(2):207-212.
[5] Park SY, Kim EJ, Shin HK, et al. In-duction of apoptosis in HT-29 human colon cancer cells by phloretin [J]. J Med Food, 2007,10(4):581-586.
[6] Zhang HT, Feng ZL, Wu J, et al.Sodium butyrate-induced death-associated protein kinase expression promote Ra ji cell morphological change and apoptosis by reducing FAK protein levels.Acta.Pharmacol.Sin, 2007, 28: 1783-1790.
[7] Wu C H, Ho YS, Tsai CY, et al.In vitro and in vivo study of phloretininduced apoptosis in human l iver cancer cells involving inhibition of type II glucose transporter[J].Int J Cancer, 2009, 124(9): 2210-2219.
[8] 羅輝,汪亞君,陳杰.根皮素誘導肝癌 BEL-7402 細胞凋亡[J].南方醫科大學學報,2008,28(7): 1249-1251.
[9] 羅輝,汪亞君,陳 杰.根皮素抑制肝癌細胞惡性表型的研究[J].中藥材,2008,31(7): 1019-1021.
[10] Frantz D J.Hughes B G,Nelson D R.Cell cycle arrest and differential gene expression in HT-29 cel1s exposed to an aqueous garlic wxtract[J].Nutr Cancer,2000,38(2):255-264.
[11] Bouchier HL, Lartigue L, Newmeyer D.Mitochondria: pharmacological manipulation of cell death[J].J Clin Invest,2005,115: 2640-2647.
[12] Amstrong JS.Mitochondrial membrane permeabilization: the sine qua non for cell death.Bioessays, 2006, 28: 253-260.
[13] Jonge PD, Wieringa T, Van Putten JP.Phloretin- an uncoupler and an inhibitor of mitochondrial oxidative phosphorylation[J].Biochim Biophys Acta, 1983, 722(1): 219-225.
[14] Jiang S, Chow SC, Nicotera P, Orrenius S.Intracellular Ca2+signals activate apoptosis in thymocytes: Studies using the Ca2+- ATPase inhibitor thapsigargin[J].Exp Cell Res,1994, 212: 84-92.
(編校:譚玲)
Effects of proliferation and apoptosis of phloretin on the hepatoma carcinoma cells HepG-2
WANG Hui,LIU Zheng,WU Han-dong
(College of Food Science and Engineering, Liaoning Medical University, Jinzhou 121001, China)
ObjectiveTo investigate proliferative and apoptotic effects of phloretin on hepatoma carcinoma cells, hepatoma carcinoma cells HepG-2 was used as research materials.MethodsThis research observed morphological alterations using phase contrast microscopy and electron microscopy, cell proliferation were detected by MTT assay, and using flow cytometry detected apoptotic rates, cell cycle progression, mitochondrial trans-membrane potential and intracellular calcium homeostasis.ResultsApoptotic cells appeared morphological alterations.Phloretin exerted a inhibitory the proliferation of HepG-2 cell line, and induced its apoptosis in a dosage and duration dependent manner.Cell cycle was arrested at G1 phase, mitochondrial trans-membrane potential dropped, intracellular free Ca2+increased.ConclusionPhloretin can induce apoptosis of HepG-2 via arresting cell cycle progression, reducing mitochondrial trans-membrane potential and disturbing intracellular calcium homeostasis.
phloretin; human hepatoma cells; apoptosis; proliferation
王會,男,碩士,講師,研究方向:細胞凋亡,E-mail: yxlwhx077@163.com。
R735.7
A
1005-1678(2015)07-0039-05
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
海峽科技與產業(2016年3期)2016-05-17 04:32:12
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
