大鱗副泥鰍部分線粒體基因的序列分析研究
2015-08-22 09:37:40韓曉磊朱綠定王富凱韓曜平朱慶華
常熟理工學院學報 2015年2期
韓曉磊,王 丹,徐 濤,朱綠定,王富凱,韓曜平,朱慶華
(1.常熟理工學院 生物與食品工程學院,江蘇 常熟 215500;2.常州市武進區前黃鎮農技農機站,江蘇 常州 213100;3.常州市秋余水產養殖專業合作社,江蘇 常州 213100)
大鱗副泥鰍部分線粒體基因的序列分析研究
韓曉磊1,王丹2,徐濤1,朱綠定3,王富凱1,韓曜平1,朱慶華3
(1.常熟理工學院 生物與食品工程學院,江蘇 常熟 215500;2.常州市武進區前黃鎮農技農機站,江蘇 常州 213100;3.常州市秋余水產養殖專業合作社,江蘇 常州 213100)
通過PCR和DNA測序技術,獲得了大鱗副泥鰍(Paramisgurnus dabryanus)線粒體基因組的部分序列,所得4段序列長度共計11552 bp,包括5個蛋白質編碼基因(ND1、ND2、ND4、ND4L、ND6和Cytb),14個tRNA基因.本研究獲得序列的堿基組成為A 28.9%、C 27.7%、G 15.7%、T 27.7%,可看出(A+T)>(C+G);測出的蛋白編碼基因的堿基都有一個明顯的現象,就是G的含量很明顯的少于其他堿基,而C的含量都比較高,并且(A+T)>(C+G),這樣的結果表現出明顯的反G偏倚和堿基組成偏好性.將大鱗副泥鰍Cytb序列與GenBank其他6種魚類的Cytb序列進行%對,表明大鱗副泥鰍與洛東江高麗鰍和俄羅斯泥鰍親緣關系最近.用NJ法構建7種魚類的進化關系樹,結果表明:大鱗副泥鰍與洛東江高麗鰍和俄羅斯泥鰍最先聚類,再與其他鰍科魚類相聚,最后與鯉科魚類斑馬魚相聚,結果與傳統分類一致.
大鱗副泥鰍;線粒體DNA;NADH還原酶復合體;Cytb
大鱗副泥鰍(Paramisgurnus dabryanus)隸屬于鯉形目,鰍科,花鰍亞科,副泥鰍屬,主要分布于四川、江蘇、浙江、臺灣,遼寧、黑龍江等地,具有很高的食用價值,俗稱“水中人參”[1-3].大鱗副泥鰍現已成為我國出口韓國的重要水產品之一,且在國內市場的需求量也在不斷攀升.作為一種優良的養殖品種,它是中國近年養殖規模發展較快的名優魚類,許多省份已開始規模化養殖生產[4-7].動物mtDNA是共價閉合的雙鏈DNA分子,核酸序列和組成比較保守,基因的排列順序比較穩定而且緊密,無重組和單拷貝.由于mtDNA含有豐富的進化信息,具有長度短、含量豐富、母性遺傳和進化速度快等特點,已成為研究動物起源進化以及群體遺傳分化的理想對象[8-13].本實驗采用PCR技術,獲得大鱗副泥鰍部分線粒體DNA的序列,為大鱗副泥鰍的系統分類提供分子水平的證據,并對其種屬水平的分類研究進行了初步探討,同時也為今后大鱗副泥鰍的資源保護、開發和養殖提供相應的理論依據.
1 材料與方法
1.1實驗樣品
大鱗副泥鰍,取自常州市武進區水產推廣站大鱗副泥鰍良種場,運輸到實驗室后進行活體宰殺,迅速采集肌肉組織后放于-80℃冰箱保存備用.
1.2實驗方法
1.2.1基因組DNA提取
基因組DNA提取參照《分子克隆試驗指南》[14].用Eppendorf公司生產的Bio Photometer核酸檢測儀檢測DNA溶液濃度與純度,并將濃度稀釋至30 ng/μL,置-20℃保存備用.
1.2.2線粒體DNA基因片段的擴增
從GenBank下載了與大鱗副泥鰍親緣關系較近的3種鰍科魚類(Cobitis striata,Misgurnus anguillicaudatus,Misgurnus nikolskyi)線粒體基因組全序列并采用DNAMAN5.0進行比對,尋找保守區域,設計了7對PCR引物(表1),引物合成由上海生工完成.用PE9600型PCR儀進行PCR擴增反應.PCR反應的總體積為50 μL,其中包括5 μL的10×Reaction Buffer,1 μL dNTP mix(10 mmol/L),引物F和R各1 μL(25 μmol/L),0.4 μL Taq DNA聚合酶(5 U/μL),1 μL總DNA(約100 ng),加滅菌雙蒸水至終體積.PCR反應程序如下:94℃預變性5 min,94℃變性30 s,55℃退火30 s,72℃延伸45 s,運行35個循環,然后再延伸10 min.最后將PCR擴增產物在2%瓊脂糖凝膠電泳(內有溴化乙錠)中電泳,以DL2000 marker為分子量標記,電泳結束后在凝膠成像系統中拍照.
1.2.3DNA序列測定

表1 大鱗副泥鰍線粒體擴增引物設計
將PCR產物割膠純化(純化試劑盒購自中科開瑞生物技術公司),然后送往上海英駿生物工程公司進行序列測定,測序在ABI3730測序儀上進行.
1.2.4數據分析
所獲得的序列經校對后,采用ClustalX1.83軟件進行比對,用MEGA5.1軟件進行分析.根據Kimura雙參數模型計算遺傳距離,用NJ法構建系統發生樹,采用bootstrap檢驗,重復次數為1000次.
2 結果與分析
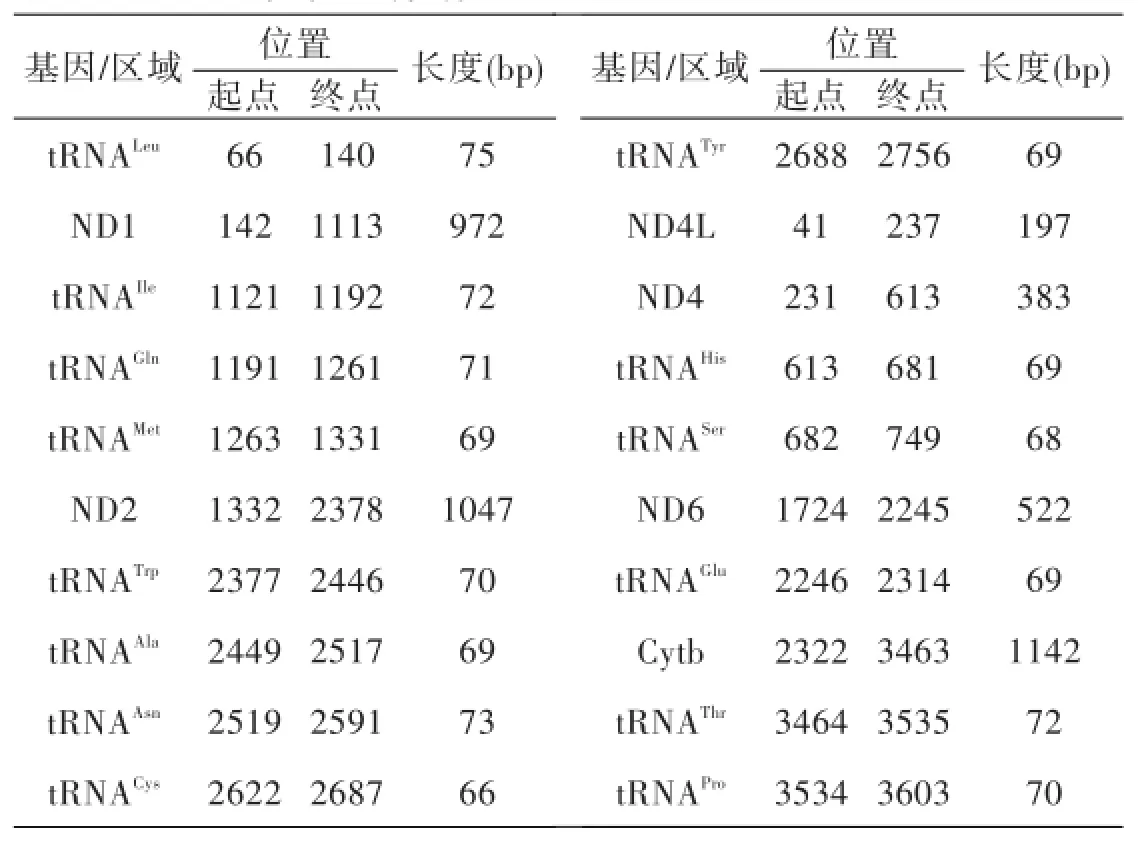
表2 大鱗副泥鰍部分線粒體基因
2.1大鱗副泥鰍部分線粒體DNA序列
本實驗共測定拼接完成4段大鱗副泥鰍的線粒體基因,拼接后長度分別為2790 bp、2861 bp、1859 bp和4042 bp,其中包括6個蛋白質編碼基因(ND1、ND2、ND4、ND4L、ND6和Cytb,14個tRNA基因,也注明了各個基因的起始位點和終止位點(表2).
2.2蛋白質編碼基因的序列分析
魚類線粒體基因組共編碼13個蛋白質,包括:細胞色素b(Cytb),2個ATP酶的亞單位(ATPase8和ATPase6),3個細胞色素C氧化酶的亞單位(COI,Ⅱ,Ⅲ),7個NADH還原酶復合體的亞單位(ND1,2,3,4,4L,5和6).我們獲得了部分蛋白質編碼基因(ND1、ND2、ND4、ND4L、ND6和Cy?tb),分析了其堿基組成及密碼子的使用情況.
2.2.1ND1基因
大鱗副泥鰍ND1基因的序列長度為972 bp(表2).堿基T、C、A、G分別是30.5%、27.6%、27.5%、14.5%;密碼子第三位上堿基T、C、A、G分別是29.6%、23.8%、41.7%、4.9%(表3).比較ND1基因總的堿基組成和第三位點的堿基組成,發現A的含量都比較高.比較ND1基因三個位點的堿基組成,發現第一位點C含量稍高;第二位點T含量最高,為40.4%,存在反G偏移,含量為11.4%;第三位點A含量最高,為41.7%,存在反G偏移,含量為4.9%.
將大鱗副泥鰍ND1基因核苷酸序列翻譯為氨基酸序列時,在所有密碼子中,UUG,CCG,UAG,CAG,AAG,UGU,UGC,CGG,AGU,AGA,AGG都沒有使用到.使用頻率最高的是CUA密碼子(30.0),其次是AUU 和GCC密碼子(都是18.0);GGG,UGG,GAU,UAA,GCG,ACG,CUG密碼子使用情況均為1.0.由于篇幅有限,此處及下文的密碼子使用情況表格均未列出.
2.2.2ND2基因
大鱗副泥鰍ND2基因的序列長度為1047 bp(表2).堿基T、C、A、G分別為25.7%、30.0%、30.5%、13.8%;密碼子第三位上堿基T、C、A、G含量分別為38.4%、34.7%、15.8%、11.2%(表3).比較ND2基因總的堿基組成和第一位點和第三位點的堿基組成,發現C的含量都比較高,第一位點C含量很高,為30.0%,存在反G偏移,只有8.0%;第三位點T含量最高,為38.4%.C為34.7%,G為11.2%,存在反G偏移.

表3 大鱗副泥鰍ND1基因密碼子各位置的堿基組成及含量(%)
將大鱗副泥鰍ND2基因核苷酸序列翻譯為氨基酸序列時,在所有密碼子中,CCG,UCG,GCG,UAG,GAA,GAG,CGA,CGG,GGA都沒有使用到.使用頻率最高的是CCU密碼子(28.0),其次是AAC密碼子(25.0);GUU,GUA,GCA,GAU,CGU密碼子使用情況均為1.0.
2.2.3ND4基因
大鱗副泥鰍ND4基因的序列長度為383 bp(表2).堿基T、C、A、G含量分別為26.9%、31.9%、27.2%、14.1%;密碼%第三位上堿基T、C、A、G含量分別為27.6%、30.7%、29.9%、11.8%(表3).比較ND4基因總的堿基組成和第一、二、三位點的堿基組成,發現C的含量都比較高,第一位點C含量很高,為35.9%,存在反G偏移,含量為18.0%;第二位點A含量最高,C為28.8%,存在反G偏移,含量為12.5%;第三位點C含量最高,為30.7%,G為11.8%,存在反G偏移.
將大鱗副泥鰍ND4基因核苷酸序列翻譯為氨基酸序列時,在所有密碼子中,CCG,CGG,AUU,AGU,AGC,AUG,ACG,AGG,GGC,GUA,GUG,GCG,GAG,GGG,UCG,UGA,CGU都沒有使用到.使用頻率最高的是CAA,CCC密碼子(6.0),其次是CUA,CCA密碼子(5.0);CUC,AUA,AAA,AAG,GCA,GAA,CGC,CGA,GGU密碼子使用情況均為1.0.
2.2.4ND4L基因
大鱗副泥鰍ND4L基因的序列長度為197 bp(表2).堿基T、C、A、G含量分別為29.9%、31.5%、19.8%、18.8%;密碼子第三位上堿基T、C、A、G含量為46.2%、29.2%、9.2%、15.4%(表3).比較ND4L基因總的堿基組成和第三位點的堿基組成,發現C的含量都比較高.比較ND4L基因三個位點的堿基組成,發現第一位點C含量很高,為30.3%,存在反G偏移,含量為10.6%;第三位點T含量最高,為46.2%,C為29.2%,存在反G偏移,含量為15.4%.
將大鱗副泥鰍ND4L基因核苷酸序列翻譯為氨基酸序列時,在所有密碼%中,UUU,UCC,UUA,UUG,UCG,UAG,CAU,CGU,CCC,CUA,CAA,CGA,CGG,AGU,AUA,AAA,AUG,ACG,AAG,GUU,GGU,GUC,GCC,GAC,GUA,GCA,GAA,GGA,GUG,GCG,GAG,GGG都沒有使用到.使用頻率最高的是CCU密碼子(8.0),其次是AUU密碼子(5.0);UUC,CUU,CUC,UCA,CCA,CCG,ACC,ACA,CAG,AAC,GAU,UGU,UGG,AGA密碼子使用情況均為1.0.
2.2.5ND6基因
大鱗副泥鰍ND6基因的序列長度為522 bp(表2).各堿基T、C、A、G含量分別為18.2%、30.8%、38.1%、12.8%;密碼子第三位上堿基T、C、A、G含量為23.0%、28.7%、40.2%、8.0%(表3).比較ND6基因總的堿基組成和第二、三位點的堿基組成,發現C的含量都比較高.第二位C含量最高,為40.8%,存在反G偏移,含量為12.1%;第三位A含量最高,為40.2%,C的含量為28.7%,存在反G偏移,含量為4.9%.
將大鱗副泥鰍ND6基因核苷酸序列翻譯為氨基酸序列時,在所有密碼子中,UUU,UGU,UGC,UGA,CGU,CUG,CCG,CAG,ACG,GUA,GUG,GG%都沒有使用到.使用頻率最高的是AAA密碼子(13.0),其次是AUU和CCC密碼子(都是11.0);UCU,UUC,UUA,UUG,UGG,CUU,CAU,CUC,CGG,AGU,AUC,AUG,AGG,GAU,GUC,GGC,GGA,GCG密碼子使用情況均為1.0.
2.2.6Cytb基因
大鱗副泥鰍Cytb基因的序列長度為1142 bp(表2).各堿基T、C、A、G含量分別為30.6%、27.0、27.3、15.1%(表3).比較Cytb基因總的堿基組成和第一、二位點的堿基組成,發現C的含量都比較高.比較Cytb基因三個位點的堿基組成,發現第一位點C含量很高,為25.7%,存在反G偏移,含量為13.4%;第二位點C含量明顯高于G,為29.4%,存在反G偏移,含量為7.6%.
將大鱗副泥鰍Cytb基因核苷酸%列翻譯為氨基酸序列時,在所有密碼子中,GUC,GUA,GCG,CGU,AGG,GGC都沒有使用到.使用頻率最高的是UUC密碼子(19.0),其次是AUU密碼子(5.0);CGC,CGA,CGG,AGU,AGC,GGA密碼子使用情況均為1.0.
2.3基于Cytb的大鱗副泥鰍系統分類分析
2.3.1大鱗副泥鰍Cytb與其他物種的同源性比較
利用DNAMAN5.0軟件,將大鱗副泥鰍Cytb的DNA序列與泥鰍,丘氏益秀朝鮮鰍,洛東江高麗鰍,俄羅斯泥鰍,后鰭花鰍和斑馬魚6個物種分別進行比對,得到大鱗副泥鰍與其他物種的同源性系數.從表10中可知,大鱗副泥鰍與其他幾種鰍科魚類的堿基同源性均在為85%~88%之間,以洛東江高麗鰍同源性最高,丘氏益秀朝鮮鰍同源性最低.
2.3.2不同物種間的遺傳距離
利用MEGA5.1軟件計算出各個物種間的遺傳距離,結果見表11(左下角為遺傳距離,右上角為同源性系數).由表可知,大鱗副泥鰍與洛東江高麗鰍之間的遺傳距離最小,為0.147,其次是俄羅斯泥鰍,為0.159;與斑馬魚的遺傳距離最大,為0.303. 2.%.3基于大鱗副泥鰍Cytb序列的系統進化樹
利用ClustalX1.83和MEGA5.1軟件,將大鱗副泥鰍Cytb序列與其他6個物種進行比對.再用MEGA5.1軟件中的NJ法構建分子進化樹(圖1).以斑馬魚為外類群,其余6個物種為內群,從所構建的系統樹可以看出,內群的6個物種分為2個支系,第1支系由丘氏益秀朝鮮鰍、后鰭花鰍和泥鰍組成;第2支系由大鱗副泥鰍、俄羅斯泥鰍和洛東江高麗鰍組成,這六種魚類都是鰍科,但分屬于不同的屬.大鱗副泥鰍與洛東江高麗鰍的親緣關系最近,與丘氏益秀朝鮮鰍親緣關系最遠.
3 討論
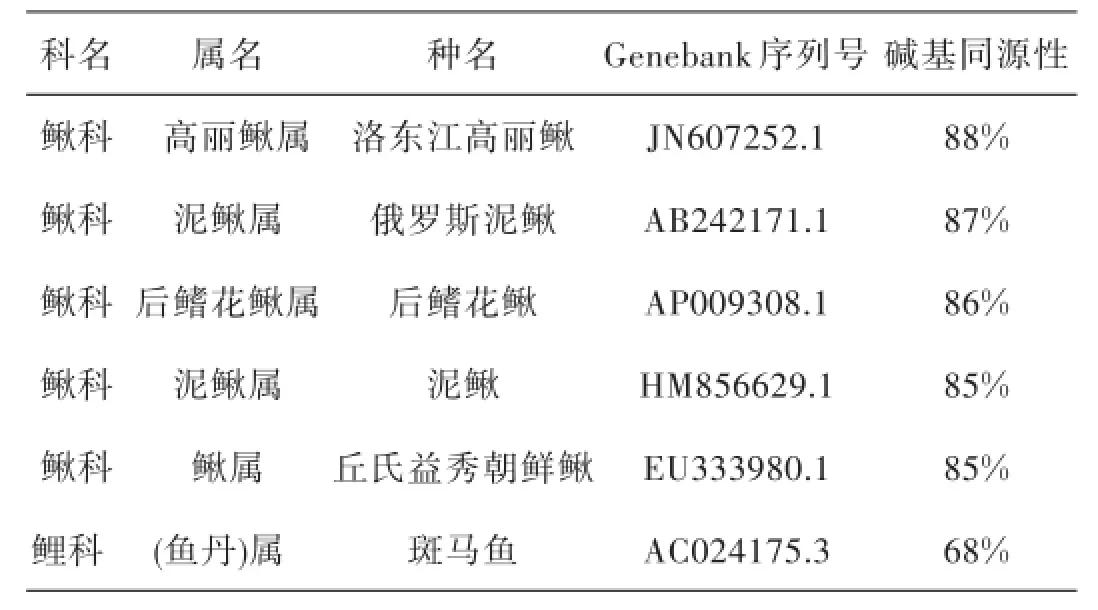
表10 大鱗副泥鰍與其他物種Cytb同源性比較
3.1MtDNA編碼基因的序列特征分析
NADH還原酶復合體亞單位ND1、ND2、ND4、ND4L和ND6以及Cytb蛋白質編碼基因中,都很明顯存在反G偏移.ND1的第一、三位,ND2第一、三位,ND4第一、二、三位,ND4L第一、三位,ND6第二、三位,分析這幾個基因的堿基組成發現,C的含量比較高,G的含量相對較低.分析這個基因三個位點的堿基組成發現,第一、二、三位存在的反G偏移概率較高,第四位堿基反G概率較低,說明第一、二、三位點對核苷酸有使用偏好.在脊椎動物中,第三位的反G偏移主要是因為該位的堿基在進化上選擇壓力比其他兩位小,表現出明顯的反G偏倚和堿基組成偏好性.
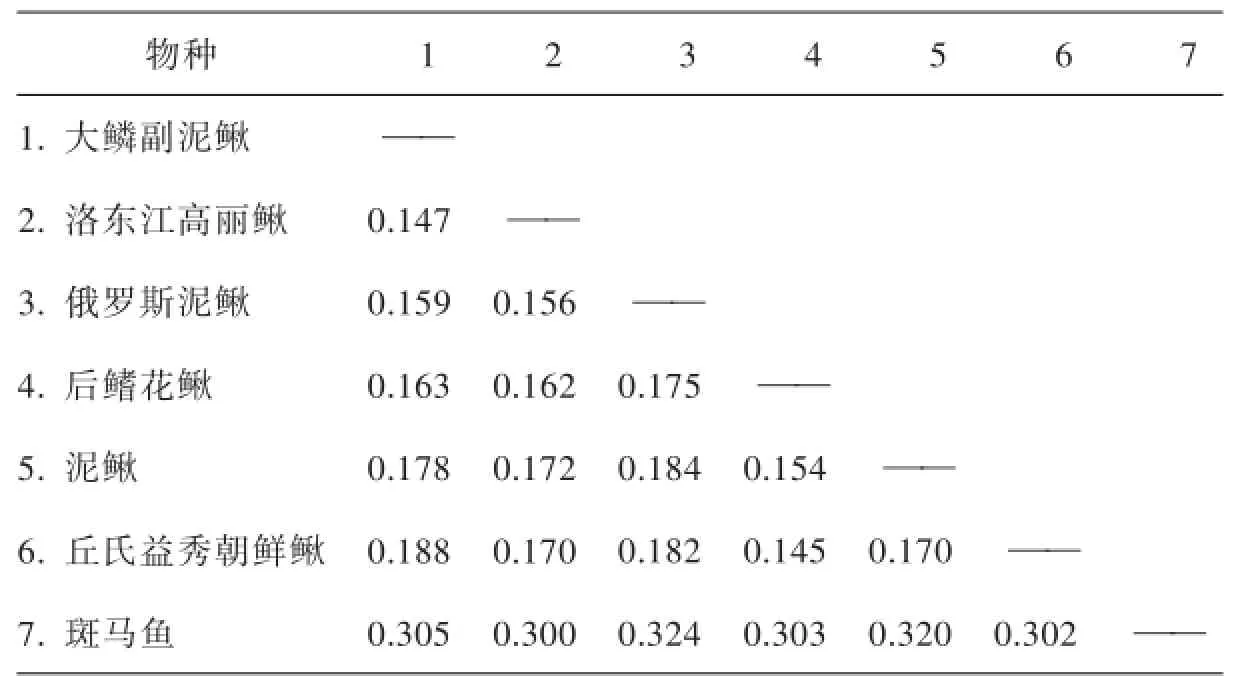
表11 幾種鰍科魚類遺傳距離比較
3.2大鱗副泥鰍Cytb在系統分類中的適用性
從表11的各個物種間的遺傳距離可知,大鱗副泥鰍與洛東江高麗鰍、俄羅斯泥鰍、后鰭花鰍泥鰍、泥鰍、丘氏益秀朝鮮鰍的遺傳距離較接近(分別是0.147、0.159、0.163、0.178、0.188),與斑馬魚遺傳距離較遠(0.305).另外表10中計算出的大鱗副泥鰍與洛東江高麗鰍、俄羅斯泥鰍、后鰭花鰍泥鰍、丘氏益秀朝鮮鰍的堿基同源性分別為88%、87%、86%、85%、85%,顯示大鱗副泥鰍與洛東江高麗鰍、俄羅斯泥鰍、后鰭花鰍泥鰍、丘氏益秀朝鮮鰍的關系較近,與斑馬魚的堿基同源性為68%,關系較遠.認為這是基于兩種算法有所差別所致,且相差很小.
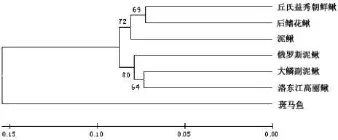
圖1 由鄰接法構建的系統樹(基于Cytb序列)
進化樹結果顯示:以斑馬魚為外群,其余6個物種為內群,主要分為2個支系,第1支系由大鱗副泥鰍與洛東江高麗鰍首先聚為一群,再與俄羅斯泥鰍聚合,BCL值均為80%;第2支系由后鰭花鰍泥鰍、丘氏益秀朝鮮鰍、泥鰍組成,首先由后鰭花鰍泥鰍、丘氏益秀朝鮮鰍組成一群,再與泥鰍聚合,BCL值為72%.
隨著研究的深入,以mtDNA中完整的基因序列或多個基因序列協同作用而獲得的遺傳信息來探討魚類的系統發育,將是今后研究的發展方向,它已在多種魚類的系統發育中得到應用[15-16].本研究基于CYTB的幾種魚系統分類與傳統形態學分類結果相符[1-2],可見Cytb基因與mtDNA中16SrRNA、12SrRNA和D-loop區等是魚類系統進化中有效的分子標記手段.
[1]孟慶聞,蘇錦祥,繆學祖.魚類分類學[M].北京:中國農業出版社,1995.
[2]倪勇,伍漢霖.江蘇魚類志[M].北京:中國農業出版社,2006.
[3]吳焱,時月,凌去非,等.蘇州地區泥鰍與大鱗副泥鰍肌肉微量元素含量比較[J].水產養殖,2011,32(10):%1-43.
[4]Ham L,Xu Y.Design of generalized E L Gamal type distal signature schemes based on discrete logarithm[J].Electron Lett,1994,30 (24):2025-2026.
[5]田功太,陳有光,段登選,等.大鱗副泥鰍人工繁育技術研究[J].河北漁業,2010(6):34-36.
[6]趙林斌.泥鰍苗種規模化繁育與高效生態養殖技術[J].中國水產,2007(5):40-42.
[7]張剛.洪澤湖大鱗副泥鰍苗種規模化繁育技術[J].中國水產,2004(3):48-50.
[8]Curole J P,Kocher T D.Mitogenomics:digging deeper with complete mitochondrial genomes[J].Tree,1999,14:394-398.
[9]Wallace D C.Mitochandrial DNA variation in human evolution,degenerative disease,and aging[J].Am J Hum Genet,1995,57: 201-223.
[10]Zhang Ya-Ping,Oliver A Ryder.Mitochondfial DNA sequence evolution in the Arctoidea[J].Proceedings of the National Academy of Sciences,1993,90(20):9557-9561.
[11]Russo CA,Takezaki N,Nei M.Efficiencies of different genes and different tree-building methods in recovering a known verte?brate phylogeny[J].Molecular biology and evolution,1996,13(3):525-536.
[12]Pollock D D,Eisen J A,Doggett N A,et al.A case for evolutionary genomics and the comprehensive examination of sequence bio?diversity[J].Molecular biology and evolution,2000,17(12):1776-1788.
[13]Roques S,Fox C J,Villasana M I,et al.The complete mitochondrial genome of the whiting,Merlangius merlangus and the had?dock,Melanogrammus aeglefinus:a detailed genomic comparison among closely related species of the Gadidae family[J].Gene, 2006,383:12-23.
[14]Sambrook J,Fritsch E F,Maniatic T.分子克隆試驗指南[M].2版.金東雁,黎孟楓,譯.北京:科學出版社,1996.
[15]肖武漢,張亞平.魚類線粒體DNA的遺傳與進化[J].水生生物學報,2000,24(4):384-391.
[16]郭新紅,劉少軍,劉巧,等.魚類線粒體DNA研究新進展[J].遺傳學報,2004,31(9):983-1000.
Sequence Analysis of Several Mitochondrial Genes in Paramisgurnus dabryanus
HAN Xiao-lei1,WANG Dan2,XU Tao1,ZHU Lv-ding3,WANG Fu-kai1,HAN Yao-ping1,ZHU Qing-hua3
(1.School of Biology and Food Engineering,Changshu Institute of Technology,Changshu 215500,China;2.Qianhuang Town Agricultural Machinery Station of Wujin District,Changzhou 213100,China;3.Qiuyu Aquiculture Professional Cooperative,Changzhou 213100,China)
This research obtained a total of 11556bp fragment of the Paramisgurnus dabryanus mitochondrial gene by using PCR method and analyzed the base composition and codon usage of protein genes in this frag?ment.The sequence contains 5 protein-encoding genes(ND1,ND2,ND4,ND4L and ND6)and one tRNA gene 14tRNA gene.The results showed that the average contents of A,T,C and G bases were 28.9%、27.7%、15.7% and 27.7%respectively,indicating a clear preference for base composition.The codon usage,base composition of ND1,ND2,ND4,ND4L,ND6 and Cytb genes and amino acid composition of the encoded proteins were ana?lyzed in this study,providing a basis for the study of genetic variation on P.dabryanus.
Paramisgurnus dabryanus;Mitochondrial DNA;NADH dehydrogenase;Cytb
Q959.468
A
1008-2794(2015)02-0110-06
2014-12-09
常州市科技支撐計劃(農業)項目“泥鰍種質資源保存與提純復壯技術研究”(CE20132014)
通訊聯系人:韓曉磊,實驗師,碩士,研究方向:淡水水生生物學,E-mail:hanxiaolei0724@163.com.
