南極春季海冰物理對葉綠素控制作用的統計分析
2015-08-30 03:00:38李志軍李潤玲王自磐ChristianHaasGehardDieckmann大連理工大學海岸和近海工程國家重點實驗室遼寧大連60國家海洋局第二海洋研究所浙江杭州00DepartmentofEarthandSpaceScienceandEngineeringYorkUniversityTorontoMJPCanadaAlfredWegenerInstituteHelmholtzCentreforPolarandMarineResearc
中國環境科學 2015年9期
關鍵詞:海冰
李志軍,李潤玲,王自磐,Christian Haas,Gehard Dieckmann(.大連理工大學海岸和近海工程國家重點實驗室,遼寧 大連 60;.國家海洋局第二海洋研究所,浙江 杭州 00;.Department of Earth and Space Science and Engineering, York University, Toronto MJP, Canada;.Alfred Wegener Institute Helmholtz Centre for Polar and Marine Research, D-7570Bremerhaven, Germany)
南極春季海冰物理對葉綠素控制作用的統計分析
李志軍1*,李潤玲1,王自磐2,Christian Haas3,Gehard Dieckmann4(1.大連理工大學海岸和近海工程國家重點實驗室,遼寧 大連 116024;2.國家海洋局第二海洋研究所,浙江 杭州 310012;3.Department of Earth and Space Science and Engineering, York University, Toronto M3J1P3, Canada;4.Alfred Wegener Institute Helmholtz Centre for Polar and Marine Research, D-27570Bremerhaven, Germany)
利用2006年南極威德爾海西北海域冬末春初的冰芯晶體結構,和302組冰芯溫度、鹽度、密度、葉綠素-a、脫鎂葉綠素數據,以及冰底面葉綠素a垂直分布數據,進一步確定積雪/海冰厚度、冰內溫度、鹽度、密度對冰內和冰下葉綠素a的貢獻.利用環境制約與生態平衡原理,闡明積雪/海冰厚度通過阻礙光通量對冰底葉綠素的效應,以及鹵水體積通過冰溫度和鹽度體現對冰藻細胞活性的抑制作用.統計分析不僅確立了南極海冰冰內鹵水體積-內葉綠素a的邏輯斯蒂上限含量曲線,并體現出粒狀冰同柱狀冰鹵水排泄方式的差異及其對冰底春季葉綠素a快速增長的冰物理貢獻.冰藻細胞活性指標小于0.5的僅占4.3%,該數據為南極冰藻是維持南極冰區浮游植物旺發的主導因子提供了重要依據.
南極;海冰;晶體結構;鹵水體積;葉綠素a;模式
極地冰雪是全球氣候變化中最敏感的物質.應對氣候變化是一種措施;而適應氣候變化也是一種策略.為此中國學者已經從不同角度尋求冰雪的物理證據[1-3].其中冰生態系統中浮游植物葉綠素a含量不僅檢測快捷,而且能反映海洋浮游植物數量同初級生產力的關系[4].一般來講,冰內葉綠素 a是指示藻類細胞生物量的一個指標[5],與海冰形成時的水體內葉綠素的水平、光照條件、溫鹽相關.因此它表現出季節性變化.極地考察中,只有定點連續觀測的固定冰冰內葉綠素 a含量的時間系列才能用于研究季節性變化[6-8].現場走航考察中所調查的浮冰不一定處在最初凍結位置,其冰內葉綠素含量不僅具有季節性變化[9],而且還具有空間變化.海冰晶體之間的鹵水通道容納不同鹽分的鹵水或者氣泡,其中鹵水中的營養鹽支撐著生物的活性.因此也可以將鹵水體積作為冰內生物活動的評價指標之一,體現對生物活動的支持能力[10-11].2006年德國組織了覆蓋南極威德爾海65°30′~66°10′S,39°~61°W區域的海冰、生物、海洋等內容的科學考察,時間屬于南極冬末早春季節,是生物繁衍非常重要的季節.作者組成海冰物理和海冰生物聯合考察組,采用現場鉆取冰芯;分組測試海冰物理參數和海冰生物參數;然后再通過資料同化,獲得相同垂直剖面上的海冰物理和生物數據集.通過定量分析,從冰物理角度研究冰基本物理性質對冰葉綠素 a含量的抑制作用,以及它們之間的定量關系.文中首次將冰內孔隙視為冰藻活動的空間,將冰內鹵水體積視為冰藻營養鹽和溫度環境的媒體[10-11],將雪/冰厚度視為光線傳輸的屏障,旨在探索雪/冰厚度對冰-水界面附近冰藻細胞活性和繁榮的影響;研究鹵水體積同冰內葉綠素a含量之間的內在聯系;建立粒狀冰和柱狀冰內鹵水體積同葉綠素 a上限含量的相關關系.實現利用環境與承載力的概念[12]、環境變化特征驅動力的概念[13],把海冰生態同海冰物理性質有機地關聯起來的目的.
1 研究方法
1.1現場測試方法
2006年9月8日至10月18日,德國北極星號科學考察船在南極威德爾海執行冬季海冰物理和生態調查.作者在南極完成 28個冰站作業.在每一個冰站,通過挖雪坑測量積雪厚度、類型、分層溫度、鹽度、密度、含水量.通過取冰芯進行冰芯厚度、類型、分層溫度測量,并在低溫實驗室進行分層組構、鹽度、密度分析;其中對特殊冰芯的部分位置或整個冰芯做薄冰片,在正常光和偏振光下觀測冰內晶體和氣泡[14].另取一支冰芯測試冰內的葉綠素a含量[15].在28個冰站中成功取得27個冰站的冰芯,作為冰物理和冰生物的分析樣品,考察站位分布參見文獻[15].
1.2數據同化方法
冰內的鹵水體積和孔隙率的計算方法由Cox等[16]給出,它們的計算需要海冰的溫度、鹽度和密度,但僅適用于-2~-22.9℃的情況.對于冰溫高于-2℃的情況(占總數的 4%),利用Lepp?ranta等[17]的方法計算.上述 2種計算都是根據海冰標準相圖的固-液平衡曲線擬合所得,因此函數形式相同,但采用的溫度系數不同.它們的鹵水體積計算均需要冰溫度和冰鹽度,計算孔隙率需要增加冰的體密度.
冰芯溫度剖面是在現場按一定間距測量的,而其鹽度和密度是在低溫實驗室內按照冰的類型分割成不同厚度的小塊,分段測量.因此實測冰溫度、鹽度、密度對應的冰芯位置不同.要將數據統一到相同的位置上,必須利用統計方法建立起冰內溫度擬合剖面,再計算出對應冰層鹽度和密度分層位置的溫度.然后再利用相同位置的溫度、鹽度、密度,計算出鹵水體積和孔隙率的豎向剖面;最后將實測的葉綠素a含量進行線性插值,得到相同位置的冰內葉綠素a含量.
另外,冰內葉綠素a含量同結冰方式,即冰的晶體結構有關[18].本次考察中共發現5類冰晶體,但主要是柱狀冰和粒狀冰;其它類型有粒狀-柱狀混合冰、骨架和多邊粒狀冰.有關冰晶體結構、溫度、鹽度和密度垂直剖面和冰內葉綠素a含量的垂直剖面均有報道[14-15].
本航次中27支具有冰物理和生物數據的冰芯共計長度 35m,其中柱狀冰占 55.16%;粒狀冰占 28.73%;粒狀-柱狀混合冰占 14.44%;骨架占1.01%;多邊粒狀冰占 0.66%.在這些冰芯上共得到冰內溫度、鹽度、密度和葉綠素 a有效數據302組.多邊粒狀冰、粒狀-柱狀混合冰、骨架3種冰晶體所占比例太少,無法進行統計分析.冰內葉綠素a的垂直分布有3種類型,與有關報道文獻總結的類型要少[19].為了同前期研究比較,本文的定義同文獻[15]的一致,分為“底部葉綠素 a遞增型”、“表、底部葉綠素a增大型”和“中部葉綠素a增大型”.
當冰層春季升溫和冰內鹵水排泄時,近冰底面的冰晶間的鹵水通道首先拓寬,和冰下海水溝通.因此,從冰底面的葉綠素 a最大值到冰底面以上20cm以上出現最低值,這段區域視作鹵水排泄區;對應的最小葉綠素a被視為起始處葉綠素a.
2 結果與討論
2.1冰內和冰下水體葉綠素a分布垂直分布
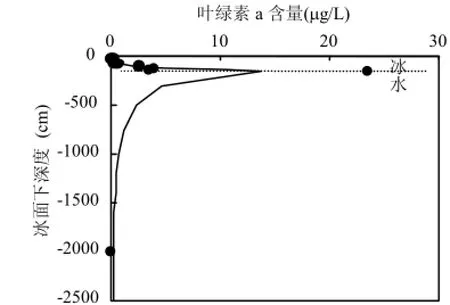
圖1 葉綠素a在冰層內部和水體中的垂直分布Fig.4 Vertical distribution of Chl-a in ice and in water
圖1是060921冰芯及其水內葉綠素a分布,葉綠素a在冰內的含量較低,在冰-水界面處出現峰值,但進入到海水,其含量繼續降低.冰下 20m處葉綠素a為0.124μg/L;冰下250m處葉綠素a 為0.006μg/L;冰下500m處葉綠素a為0.003μg/L.這些表明隨著水深的增加,水體內的光照減弱,葉綠素a含量隨水深在不斷降低[20-21].Wang等曾報道秋季威德爾海表層葉綠素 a平均值在 0.11~0.22μg/L,而本研究發現表層水內葉綠素 a含量的平均值范圍僅 0.01~0.05 μg/L,均說明在南極冬季,海冰極大地抑制了冰下海水中冰藻光合作用,導致海洋初級生產處于全年最低水平[22].與之相反,浮冰中葉綠素a含量很高,表明冬季冰內冰藻在維持南大洋初級生產力和生態系統正常運行中起到了關鍵作用.何劍鋒等曾發現,在春季固定冰中也有類似圖1中的冰底峰值,他們認為當海冰底部骨架層遇到合適的水文氣象條件,成為冰藻最佳生存環境,冰藻開始繁殖,水體內也出現水華,同時此處冰藻的繁榮導致葉綠素 a含量超出冰內1~3個量級[23].因此冰底是海冰生物量最高的部位,與該處的溫度、光輻照度、冰結構、冰藻種類和冰藻群落結構、鹽度、營養鹽、氨基酸以及主要金屬離子等有關;其中,與冰雪物理相關的最顯著影響因子是冰下水溫和光合作用.
2.2海冰內部溫度和鹽度對冰內葉綠素a的貢獻
冰在凍結過程中,附著在冰核水體中的藻類,在凍結過程中聚集[18].冰生長的方式不同,對應的晶體類型也不同,因此冰內初始葉綠素含量同冰晶體類型和內部組構也有關系.所以,對冰內葉綠素的定量分析既要考慮冰晶體類型,還要考慮冰內的鹵水體積.
以粒狀冰為主的冰內葉綠素 a的豎向剖面主要是中部增大型,而以柱狀冰為主的冰內葉綠素a的豎向剖面主要是底部增大型,并且粒狀冰內的初始值比柱狀冰內的略高.產生這二種類型的主要原因是柱狀冰封閉在晶體間的鹵水雖然比粒狀冰內的少,但是在冰溫升高時,柱狀冰內鹵水更容易在重力作用下沿著增寬的鹵水通道向冰底遷移[24],并富集在冰-水界面附近,從而給冰藻活躍提供了條件[22].
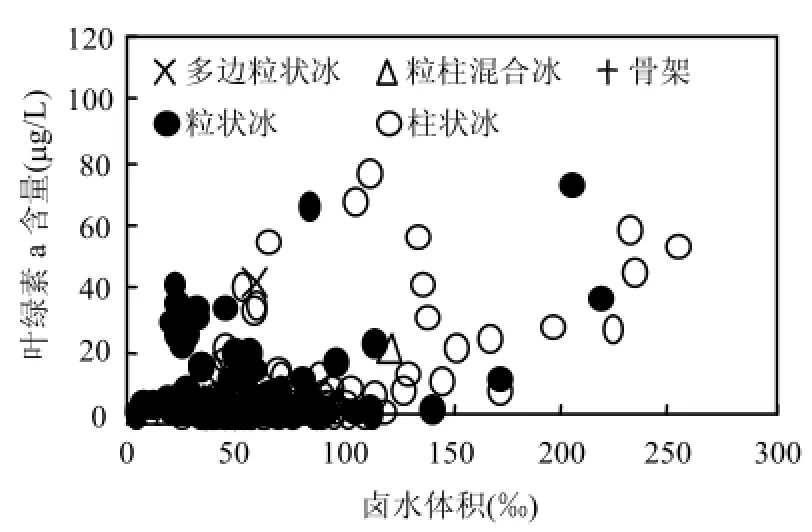
圖2 冰內葉綠素與同鹵水體積的散點關系Fig.4 Scatter relation between Chl-a with brine volume
由圖2可見,分析發現葉綠素a含量隨鹵水體積呈正比關系,且存在上限值,說明鹵水體積是冰內葉綠素a的控制因子之一,體現溫度和營養鹽的貢獻.
2.3冰內鹵水體積同冰藻細胞活性的關系
冰藻死亡時,葉綠素中的鎂離子被取代轉變成脫鎂葉綠素;因此葉綠素a含量在葉綠素a及脫鎂葉綠素之和中所占的比值(R=葉綠素 a/(葉綠素 a+脫鎂葉綠素))可作為反映冰藻細胞生理狀態的一個指標[15].R值越大(接近1.0),說明冰藻細胞活性越強,所含色素主要為葉綠素 a,冰藻種群生長旺盛,碳固定能力強,生產潛力高.當 R值降低到0.8,尤其小于0.5時,說明冰藻葉綠素a已被高度降解,細胞活性下降,潛在生產力低.在其最大鹵水體積 250‰以內,其接近最大值 1.0.表達式見圖 3.垂直虛線來自評估冰藻細胞活性能力的平均上線在R值為0.8時的鹵水體積交叉點;水平虛線來自最低潛在生產力臨界線對應的R=0.5.由圖 3可見,在南極惡劣環境中,無論是粒狀冰還是柱狀冰,當冰內的鹵水體積超過 8‰,它們內部的冰藻是活躍的;但當鹵水體積小于 8‰,冰藻細胞活性指標R值小于0.8.
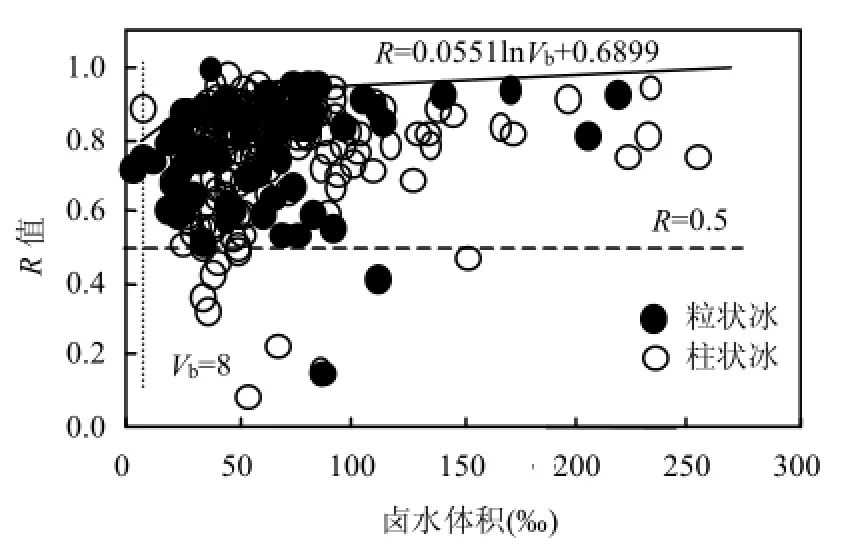
圖3 粒狀冰和柱狀冰冰內R值與鹵水體積的關系Fig.4 The relation between the R value and brine volume in granular ice and columnar ice
所有302個有效試樣所的冰鹽度95%范圍內為(2.100~9.000)‰、密度95%范圍內為(0.769~0.950)g/cm3,平均值分別為4.824‰和0.897g/cm3.根據圖 3,如果以鹵水體積 8‰為界限,當冰內鹽度和密度為平均值時,冰溫度可低至-38.0℃.在南極嚴寒的冬季,冰溫低于-38.0℃比較罕見,所以冰內供應冰藻細胞活性鹵水的溫度僅會略低于海水冰點(-1.8℃)[25],且鹵水體積較充足,因此南極冬季葉綠素a有達到上限值的條件,但事實上許多低于上限值.而在春季,冰溫升高,鹵水體積增加,特別是鹵水在重力作用下排泄.如果將海冰視為一個封閉體系,春季調查期間的冰內營養鹽只是在冰層的垂直分布發生變化,總營養鹽沒有增加,遷移并集中在冰層底部鹵水中的營養物質結合適合的鹵水溫度環境足可以支撐冰藻的繁榮,接近上限的所占比例數增加.
根據 Dai等[15]的分析,當年新生冰 R值為0.83,當年冰R值為0.78,二年冰及多年冰R值為0.80,說明當年新生冰中的冰藻細胞活性最高.何劍鋒等也指出,南極固定冰冰底R值在全年都高于 0.80,揭示出冰底冰藻一直具有較強的細胞活性[23].冰藻能在冰底大量繁殖(特別是在春季)的原因之一是南極冰藻在較低光強條件下的仍然保持旺盛的生命力[26],同時冰底具有較為衡定的溫度,冰結構疏松,又能從冰-水界面及時補充得到藻類繁殖所需的營養鹽,為冰藻提供了其它冰層不具備的良好生長環境,特別是冰骨架層內的鹵水營養鹽是限制冰藻細胞活性的環境因子[27].
如果以冰為載體,鹵水為生存資源,葉綠素 a為冰藻細胞活性能建立南極冰藻簡化生態系統成立的話,它就構成環境對生態的驅動[13],有一個生態平衡的極限,即冰內葉綠素a上限值所代表的冰藻細胞活性不應超過冰內鹵水體積的允許承載量.這同各種環境中的環境-生態的平衡關系類似[12].
設鹵水體積含量為νb,葉綠素a含量為P,鹵水體積控制下的最大葉綠素a為PA,則冰內鹵水體積能夠提供小于或等于PA對應所需的營養鹽等環境條件;同樣假設冰內鹵水不能提供的最小生態承載力為 PB,則 P(νb)隨νb的變化可用一個簡單的表現生物種群增長過程邏輯斯蒂(Logistic)增長模型表示,其形式為:
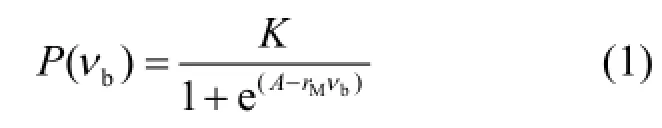
式中:K是在一定鹵水體積下冰藻達到“飽和”狀態時對應的葉綠素a含量;rM是在一定鹵水體積下葉綠素a最大增長速率;A是積分常數.
在邏輯斯蒂曲線中有一拐點,該拐點的葉綠素a含量為冰藻最適密度,即冰藻增長率最快時對應的葉綠素a含量.
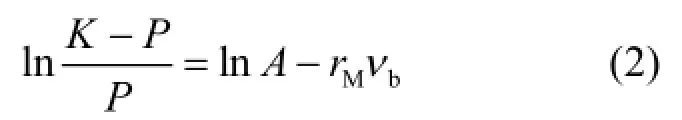
統計中,以ln((K-P)/P)和νb為坐標作圖,擬合得到一條直線, lnA為該直線的截距,rM是該直線的斜率.
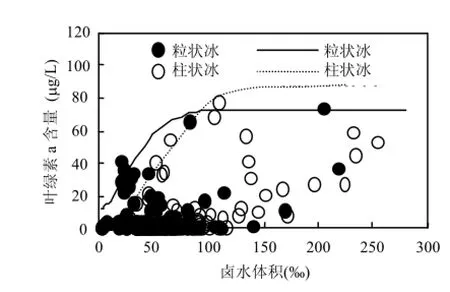
圖4 粒狀冰和柱狀冰冰內葉綠素a隨鹵水體積變化的上限含量曲線Fig.4 The upper limit curves of Chl-a with brine volume in granular ice and columnar ice
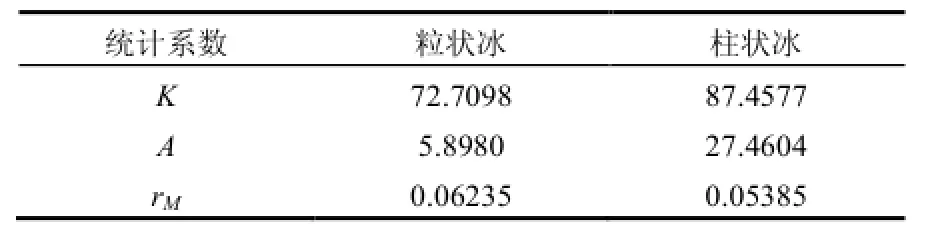
表1 粒狀和柱狀冰葉綠素a上限含量隨鹵水體積變化曲線的統計系數Table 1 The statistical coefficients in the upper limit curves of Chl-a with brine volume in granular ice and columnar ice
統計曲線表明,用鹵水體積來分析冰藻葉綠素a上限是合理的,存在低于上限含量的數據分散是南極惡劣環境下的正常現象(圖4).表1系數說明,粒狀冰和柱狀冰的K值處于同一水平,但柱狀冰的略高.說明在春季冰溫升高,冰內鹵水體積增加時,以柱狀冰為主的冰晶體內的冰藻細胞活性大,對應的葉綠素a含量高.粒狀冰的A值較低,說明在隆冬季節粒狀冰內所包裹的鹵水體積較高,維持較高的冰藻細胞活性.rM反映出粒狀冰內的冰藻早于柱狀冰內的繁榮,但只發生在冰藻開始繁榮期,過后進入鹵水體積增加階段,以柱狀冰為主的冰層在冰-水界面處體現強勢的繁榮(K值增高).
2.4積雪和海冰厚度對冰底葉綠素a的控制作用
積雪和冰層厚度越大,冰下的光線越弱[20],給冰藻提供光合作用的環境條件也越差.南極海冰表面均存在積雪,即使厚度只有厘米量級,太陽輻射也會被大幅反射.光線在雪中的衰減率要遠遠大于在冰中的衰減率[28],因此積雪的存在導致傳遞到冰內和冰下的太陽輻射強度大大減弱.
考慮到浮冰形成位置不一定是調查位置,以及冰內初始葉綠素a含量存在差異,本文不直接分析冰底部葉綠素含量 a,而是首先假定冰內鹵水排泄同冰底部葉綠素a繁榮有一定聯系,繼而將冰底部葉綠素 a同冰內鹵水排泄處起始葉綠素 a含量的比值作為一個客觀參數.這樣既能兼顧冰內鹵水排泄時的葉綠素a對冰底葉綠素a春季繁榮的貢獻,又能消除浮冰形成時包裹到冰內鹵水中初始葉綠素a含量的差異.利用觀測的積雪厚度、冰厚度與該比值建立相關關系,見圖5.
并通過曲面擬合得到關系式:
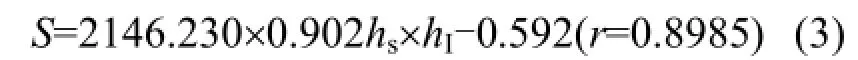
式中:S為冰底部葉綠素 a同冰內平均葉綠素 a或者冰表層葉綠素a的比值; hS為雪厚, cm; hI為冰厚, cm.
從圖5中可以發現,積雪厚度的影響大于冰厚度的影響,同 Maykut等報道的積雪和冰厚度對光強的衰減作用有相似之處(圖6)[20].圖6證實積雪和海冰厚度對光強存在直接聯系.由于光強與冰藻細胞活性存在直接聯系,因此積雪和海冰厚度同葉綠素含量呈反比關系.這種關系揭示了積雪和海冰厚度對冰下藻類的光合作用起到抑制作用,影響到藻類的生長,導致葉綠素a含量減少[29].目前冰光學中的廣泛成果[19,30-31]還沒有同冰藻直接建立聯系,圖5和圖6可將冰下光強同冰藻的生長聯系起來,但發展到同具體冰藻種類和冰藻群落的定量描述尚需努力.難點之一是當年新生冰區的冰藻春季水華期間,冰下輻照度通常小于冰表入射光的 1%[32-33],且冰表入射光在穿透冰層的過程中其有效光強和光譜成分都將受到冰內鹵水含量或冰藻顏色差異帶來的影響,因此即使在相同積雪或者冰厚度條件下仍存在冰內輻射衰減的區別[20-21].
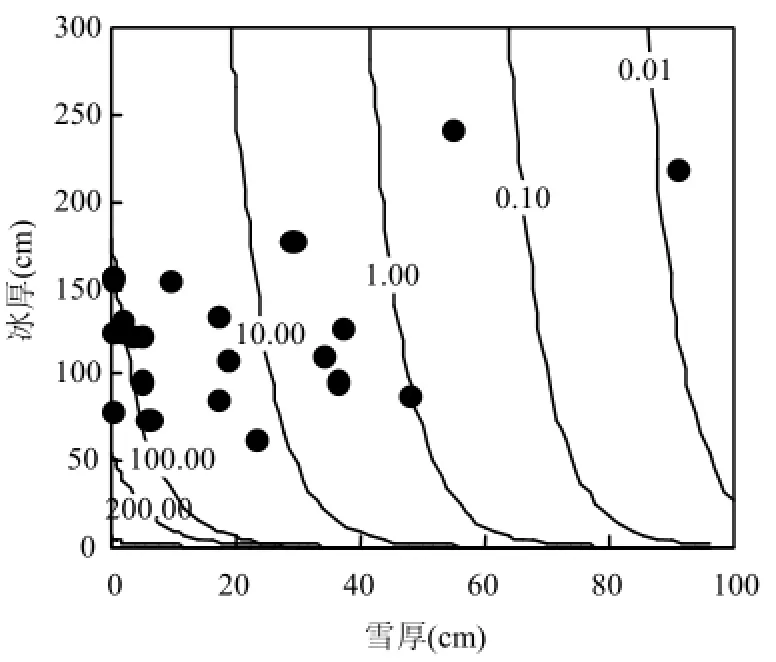
圖5 冰層底部和冰內鹵水排泄處葉綠素與比值同積雪和冰厚度的統計關系Fig.4 The statistical relation among the ratio of Chl-a under ice bottom and at the start point of brine drain section with snow thickness and ice thickness
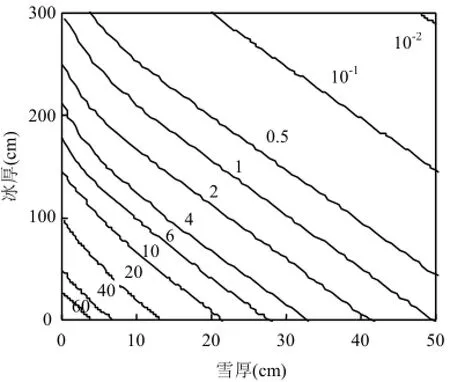
圖6 冰下總入射輻照度與冰厚和雪厚的關系[20]Fig.4 Dependence of total downwelling irradiance beneath the ice on ice thickness and snow depth[20]等高線代表雪表面總光強的百分數
3 結論
3.1春季南極威德爾海海冰內部葉綠素a的分布基本上是冰底面偏高,然后同冰-水界面的冰藻水花匹配.柱狀冰的鹵水通道比粒狀冰的更容易使鹵水向冰-水界面遷移,從而體現出柱狀冰底面的春季葉綠素a增幅大于粒狀冰,但粒狀冰形成時更容易包裹鹵水,因此粒狀冰內的葉綠素a水平比柱狀冰內偏高.水體內的葉綠素a隨著冰下水深的增加,迅速減小.
3.2冰內葉綠素a的水平和冰的晶體類型有一定關系,冰內鹵水是葉綠素a的控制因子.冰藻細胞活性指標同冰晶體類型和冰內鹵水體積存在定量關系.分析表明南極冬季冰內鹵水體積減少,但冰藻細胞活性指標維持在 0.5以上,反映出冬季冰藻的存在是海冰生物群落活動的基礎;而且在春季較高冰溫時,冰內鹵水體積增加,冰藻細胞活性水平趨于穩定.在此基礎上,利用邏輯斯蒂模型,得到南極冰內鹵水體積能夠支撐葉綠素a的上限含量曲線,表明南極海冰鹵水體積的最大承載力.而低于曲線的分散數據是南極惡略環境下生物脆弱性的證據.
3.3積雪和海冰厚度對冰底葉綠素a繁榮起著關鍵作用.它們的貢獻體現到積雪和海冰對太陽光的吸收,從而影響到冰藻的光合作用.其中積雪厚度對冰藻光合作用的效果大于海冰厚度.三者之間存在定量關系.
[1] 王星東,李新武,梁 雷.南極冰蓋凍融的時空分析 [J]. 中國環境科學, 2014,34(5):1303-1309.
[2] 武炳義,黃榮輝,高登義.北大西洋接壤的北極海冰和年際氣候變化 [J]. 科學通報, 2000,45(15):1993-1998.
[3] 艾松濤,王澤民,譚 智,等.北極 Pedersenbreen冰川變化(1936~1990~2009年) [J]. 科學通報, 2013,58(15):1430-1437.
[4] Olsen M S, Callaghan T V, Reist J D, et al. The changing Arctic cryosphere and likely consequences: An overview [J]. AMBIO A Journal of the Human Environment, 2011,40:111-118.
[5] Tedesco L, Vichi M, Thomas D N. Process studies on the ecological coupling between sea ice algae and phytoplankton [J]. Ecological Modelling, 2012,226:120-138.
[6] 陳興群,G迪克曼.南極威德爾海陸緣固冰區葉綠素a及硅藻的分布 [J]. 海洋學報, 1989,11(4):501-509.
[7] 何劍鋒,陳 波,黃鳳鵬.南極中山站近岸海冰生態學研究,2.1992年冰下水柱浮游植物生物量的季節變化及其與環境因子的關系 [J]. 南極研究(中文版), 1996,8(2):23-34.
[8] Meiners K M, Norman L, Granskog M A, et al. Physicoecobiogeo chemistry of East Antarctic pack ice during the winter-spring transition [J]. Deep-Sea Research II, 2011,58:1172-1181.
[9] Arrigo K R, van Dijken G L. Annual changes in sea-ice, chlorophyll a, and primary production in the Ross Sea, Antarctica [J]. Deep-Sea Research II, 2004,51:117-138.
[10] Mundy C J, Barber D G, Michel C. Variability of snow and ice thermal, physical and optical properties pertinent to sea ice algae biomass during spring [J]. Journal of Marine Systems, 2005,58:107-120.
[11] Hendry K R, Rickaby R E M, de Hoog J C M, et al. The cadmium-phosphate relationship in brine: biological versus physical control over micronutrients in sea ice environments [J]. Antarctic Sciences, 2010,22(1):11-18.
[12] 楊 靜,張仁鐸,翁士創,等.海岸帶環境承載力評價方法研究[J]. 中國環境科學, 2013,33(S1):178-185.
[13] 周 沙,黃躍飛,王光謙.黑河流域中游地區生態環境變化特征及驅動力 [J]. 中國環境科學, 2014,34(3):766-773.
[14] Li Z J, Nicolaus M, Toyota T, et al. Analysis on the crystals of sea ice cores derived from Weddell Sea, Antarctica [J]. Chinese Journal of Polar Science, 2010,21(1):1-10.
[15] Dai F F, Wang Z P, Yan X J, et al. Physical structure and vertical distribution of chlorophyll a in winter sea ice from the northwestern Weddell Sea, Antarctica [J]. Acta Oceanologica Sinica, 2010,29(3):97-105.
[16] Cox G F N, Weeks W F. Equations for determining the gas and brine volumes in sea-ice samples [J]. Journal of Glaciology, 1983, 29(102):306-316.
[17] Lepp?ranta M, Manninen T. The brine and gas content of sea ice with attention to low salinities and high temperatures [R]. Finnish Institute of Marine Research, 1988.
[18] Ackley S E. Ice scavenging and nucleation: Two mechanisms for incorporation of algae into newly forming sea ice [J]. EOS Transactions of the American Geophysical Union, 1982,63(3):47.
[19] 何劍鋒,王桂忠,李少菁,等.南極海冰區冰藻類群及興衰過程[J]. 極地研究, 2003,15(2):102-114.
[20] Maykut G A and Grenfell T C. The spectral distribution of light beneath first-year sea ice in the Arctic Ocean [J]. Limnology and Oceanography, 1975,20(4):554-563.
[21] Palmisano A C, Beeler Soohoo J, Sullivan C W. Effects of four environmental variables on photosynthesis-irradiance relationships in Antarctic sea-ice microalgae [J]. Marine Biology, 1987,94(2):299-306.
[22] 王自磐, Dieckmann G, Gradinger R.南極威德爾海新生期海冰生態結構I.葉綠素a與營養鹽 [J]. 極地研究, 1997,9(1):9-17.
[23] 何劍鋒,陳 波.南極中山站近岸海冰生態學研究, I. 1992年冰藻生物量的垂直分布及季節變化 [J]. 南極研究(中文版), 1995,7(4):53-64.
[24] Eicken H. Salinity profiles of Antarctic sea ice: field data and model results [J]. Journal of Geophysical Research, 1992,97(C10):15545-15557.
[25] Suzuki Y, Kudoh S, Takahashi M. Photosynthetic and respiratory characteristics of an Arctic ice algal community living in low light and low temperature conditions [J]. Journal of Marine Systems, 1997,11(1/2):111-121.
[26] Palmisano A C and Sullivan C W. Physiological Response of Micro-Algae in the Ice-Platelet Layer to Low-Light Conditions [M]. Antarctic Nutrient Cycles and Food Webs, 1985,84-88.
[27] Robinson D H, Arrigo K R, Brookhaven Z K, et al. Photophysiological evidence of nutrient limitation of platelet ice algae in McMurdo Sound, Antarctic [J]. Journal of Phycology, 1998,34:788-797.
[28] Garrison D L, Sullivan C W and Ackley S F. Sea ice microbial communities in Antarctica [J]. Bio. Sciences, 1986,36(4):243-250.
[29] Campbell K, Mundy C J, Barber D G, et al. Characterizing the sea ice algae chlorophyll a-snow depth relationship over Arctic spring melt using transmitted irradiance [J]. Journal of Marine Systems, 2014, http://dx.doi.org/10.1016/j.jmarsys.2014.01.008.
[30] Eicken H. The role of sea ice in structuring Antarctic ecosystems [J]. Polar Biology, 1992,12(1):3-13.
[31] Nicolaus M, Hudson S R, Gerland S, et al. A modern concept for autonomous and continuous measurements of spectral albedo and transmittance of sea ice [J]. Cold Regions Science and Technology, 2010,62:14-28.
[32] Sullivan C W, Palmisano A C. Sea ice microbial communities in McMurdo Sound, Antarctica [J]. Antarct. J. U. S., 198116:126-127.
[33] Palmisano A C, Sullivan C W. Sea ice microbial communities (SIMCO), I. Distribution, abundance and primary production of ice micro algae in McMurdo Sound in 1980 [J]. Polar Biology, 1983,2(3):171-177.
Statistical analysis of sea ice physical conditions effect on chlorophyll during the austral spring in Antarctica.
LI Zhi-jun1*, LI Run-ling1, WANG Zi-pan2, HAAS Christian3, Dieckmann Gerhard4(1.State Key Laboratory of Coastal and Offshore Engineering, Dalian University of Technology, Dalian 116024, China;2.Second Institute of Oceanography, State Oceanic Administration, Hangzhou 310012, China;3.Department of Earth and Space Science and Engineering, York University, Toronto M3J1P3, Canada;4.Alfred Wegener Institute Helmholtz Centre for Polar and Marine Research, D-27570Bremerhaven, Germany).
China Environmental Science, 2015,35(9):2813~2819
The sea ice core texture, and 302 groups of ice core temperature, salinity, density, chlorophyll a (Chl-a) as well as phaeophytin, Chl-a under ice from Northern West of Weddell Sea, Antarctica during the period between winter and spring of 2006 were statistically analyzed. The contributions of snow/ice thickness, ice temperature, ice salinity and ice density to the Chl-a content in ice and Chl-a blooms under ice were determined. The snow/ice thickness effect on the Chl-a blooms under the ice due to light absorption was determined. The relationship between the activity of ice algae cells and brine volume (function of ice temperature and ice salinity) in ice was investigated following the principle of environment control on the ecological balance. The statistical analysis not only revealed the relationship between the upper limits of Chl-a and brine volume in Logistic model, but also showed that granular and columnar ice crystals affect the brine drainage. The 4.3% of the index of active algal cells less than 0.5showed that the physical properties of sea ice not only affect the Chl-a blooms near ice bottom and at the ice-water interface in the austral Spring, but also are a key control element modulating the marine phytoplankton bloom in the ice covered waters around Antarctica.
Antarctic;sea ice;ice crystals;brine volume;chlorophyll a;modes
X55;P728.2;Q178.1
A
1000-6923(2015)09-2813-07
2015-01-05
國家自然科學基金(51221961,41376186);國家海洋局極地考察辦公室對外合作項目(IC201209);國家重點實驗室自主項目(SL14-5-3)
*責任作者, 教授, lizhijun@dlut.edu.cn
李志軍(1960-),男,河北崇禮縣人,教授,博士,主要從事極地和渤海冰物理和力學性質研究.發表論文300余篇.
猜你喜歡
海洋通報(2021年5期)2021-12-21 06:19:42
海洋通報(2021年3期)2021-08-14 02:20:38
成都信息工程大學學報(2021年2期)2021-07-22 07:23:32
文萃報·周五版(2020年1期)2020-02-14 10:23:59
極地研究(2018年2期)2018-06-27 09:09:34
電子技術與軟件工程(2016年24期)2017-02-23 19:13:41
海洋世界(2016年4期)2016-06-15 01:51:28
中國學術期刊文摘(2016年8期)2016-02-13 13:04:44
河北遙感(2014年4期)2014-07-10 13:54:59
自然資源遙感(2014年3期)2014-02-27 11:56:43
