遼東灣網采浮游植物粒級結構的脅迫響應
2015-08-30 03:00:35宋廣軍王年斌趙海勃遼寧省海洋水產科學研究院遼寧大連116023遼寧省海洋環境監測總站遼寧大連116023
中國環境科學 2015年9期
宋 倫,宋廣軍,王年斌,趙海勃,田 金,楊 爽,杜 靜(1.遼寧省海洋水產科學研究院,遼寧 大連 116023;2.遼寧省海洋環境監測總站,遼寧 大連 116023)
遼東灣網采浮游植物粒級結構的脅迫響應
宋 倫1,2*,宋廣軍1,2,王年斌1,2,趙海勃1,2,田 金1,2,楊 爽1,2,杜 靜1,2(1.遼寧省海洋水產科學研究院,遼寧 大連 116023;2.遼寧省海洋環境監測總站,遼寧 大連 116023)
基于2012~2014年春、夏、秋、冬季遼東灣網采浮游植物及環境因子的調查資料,對遼東灣南、北2個區域的浮游植物粒級結構分布規律及其環境脅迫響應進行了分析,并評估了浮游植物群落結構的穩定性.結果表明,遼東灣海域浮游植物春、夏、秋季豐度主要集中在小粒徑范圍,冬季主要集中在中粒徑范圍;生物量春、夏、秋季主要集中在中粒徑范圍,冬季主要集中在大粒徑范圍,南部區域浮游植物粒徑整體上大于北部區域.寡營養水域以及較低的水溫環境更適于大粒徑浮游植物生存.春、秋、冬季浮游植物群落結構比較穩定,但夏季偏離平衡點.南部區域群落結構整體上比北部區域穩定,網采中、大粒徑組的W統計量可以作為浮游植物群落結構穩定性的評估指標.
浮游植物;粒級結構;ABC曲線;生態對策;生態穩定性;遼東灣
浮游植物作為海洋初級生產力的主要貢獻者,參與物質循環和能量流動,在海洋生態系統中發揮著重要的生態作用.浮游植物粒級差異及演變規律是許多生物和非生物環境因素共同作用的結果[1],浮游植物對營養鹽吸收、污染物脅迫等存在一定程度的粒徑效應[2-4],比較不同條件下浮游植物的粒級分布對研究生態系統的結構和功能以及探討不同粒徑的進化機制都有非常重要的意義.目前針對浮游植物粒級結構的研究大多數都是采用葉綠素 a分級測定的方法來粗略表示不同粒徑生物量結構,但無法了解不同物種的生物量分布.顯微測微計法雖然耗時費力,但可進行種間定量描述和深入的作用機理研究.國內學者孫軍等[5]首次計算了 87種中國近海常見浮游植物的細胞體積、鮮重、碳含量、氮含量,為后來學者研究長江口[6]、南黃海[7]、大亞灣[1]浮游植物粒級結構及生物量譜奠定了基礎.
遼東灣為渤海半封閉式海灣,多年來超負荷承載陸源污染,富營養化加劇,赤潮現象頻繁發生[8-9].近些年,遼東灣浮游植物的研究多集中在種群結構及生態分布規律方面[10-15],在粒級結構對環境脅迫響應及穩定性評估方面鮮有研究.宋倫等[16]對遼東灣河口區和港口區渾濁帶海域的浮游生物粒徑結構進行了研究,發現浮游植物群落對無機氮含量高的渾濁帶海域響應可能是通過小粒徑生物進行反饋調節的.
本研究選取遼東灣南、北2個區域,采用顯微測微計法對浮游植物粒級結構進行了對比分析,研究了群落粒徑組成及季節變化規律,并首次利用ABC曲線法探討了2個區域群落的穩定性狀況及對環境脅迫變化響應規律,揭示了富營養海域浮游植物的粒徑分布、生物量更替原因及其環境響應機制,可為遼東灣海域赤潮監管和生態修復效果評價提供技術支持.
1 材料與方法
1.1調查區域與樣品采集
2012~2014年5月(春)、8月(夏)、10月(秋)、12月(冬,2012年未調查)分別在遼東灣海域進行了網采浮游植物及環境因子調查,于南(S區)、北(N區)2個區域分別設置10個調查站位,其中N區鄰近河口,受近岸陸源環境影響較大,S區則相對影響較小(圖1).
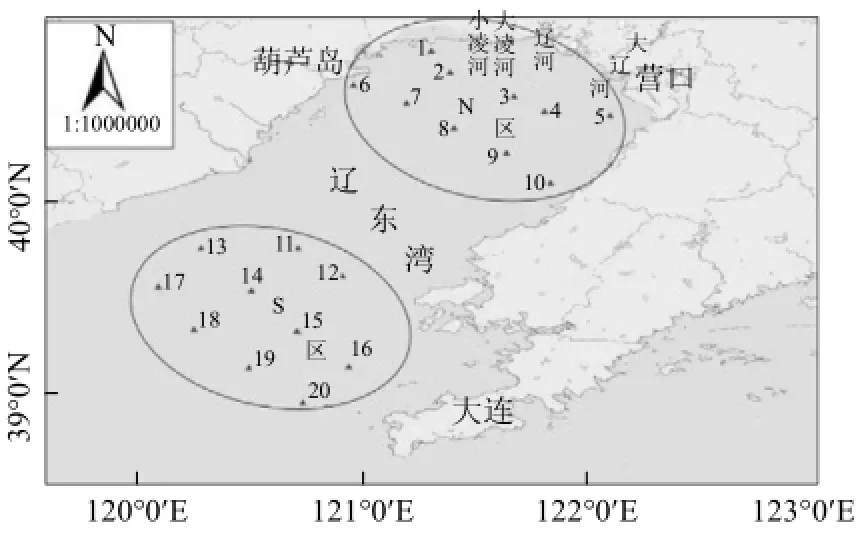
圖1 遼東灣采樣站位示意Fig.4 Sampling sites in the Liaodong Bay
浮游植物采用《海洋調查規范》[17]推薦的淺水Ⅲ型浮游生物網從底至表層垂直拖網;同期進行海水的水溫、鹽度、葉綠素a(Chl a.)、無機氮(DIN)[銨鹽(NH4+)、硝酸鹽(NO3-)、亞硝酸鹽(NO2-)之和]、活性磷酸鹽(DIP)、石油類(Oil)、pH值、溶解氧(DO)、COD、重金屬汞(Hg)、鉛(Pb)、銅(Cu)、鋅(Zn)、鎘(Cd)、砷(As)等指標的監測.調查和監測分析方法均參照《海洋調查規范》(GB/T 12763-2007)和《海洋監測規范》(GB17378-2007)[18]執行.
1.2生物粒級分析與數據處理
實驗室內對浮游植物樣品進行鏡檢、計數,并測量其幾何參數(對于單個游離細胞,只統計長軸大于 5μm的個體,對于鏈狀細胞,測量總長度后換算平均值,從粒徑及營養級角度考慮,本文沒有統計夜光藻),對于難以準確鑒定到種的個體,按未定種歸類到屬記錄,根據測定的形態參數、借助數學幾何運算公式和體長-含碳量的經驗公式分別對浮游植物個體體積和個體含碳量進行估算[5,19-20].浮游植物濕重生物量為細胞體積×1.1,浮游植物粒徑大小統一換算成等效球徑(ESD)來衡量.
采用相對重要性指數(IRI)確定群落中的優勢種:

式中:W為某種類的生物量百分比;N為某種類的豐度百分比;F為調查某種類出現的頻率[21].
采用 Shannon-Wiener指數(H′)[16]統計物種多樣性水平:
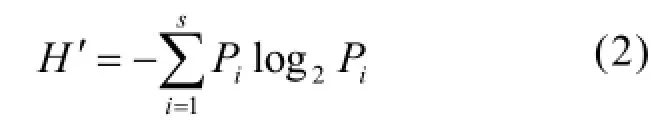
式中:Pi為第i種生物的豐度(或生物量)與樣本總豐度(或生物量)的比值;S為種類數.
利用營養指數法(E)并結合CN/CP(無機氮/無機磷)對研究海域的營養狀況進行分類[22]:
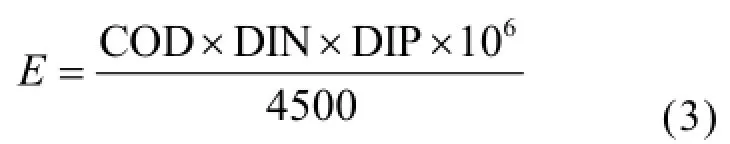
式中:E為富營養化判斷值(E),COD、DIN、DIP分別為COD(mg/L)、無機氮(mg/L)、無機磷(mg/L)的測定值.若E≥1,即為富營養化.
環境指數(測定值/標準值)中標準值參照《海水水質標準》[23](GB3097-1997)中的第二類海水水質標準計算.
通過ABC曲線法對群落的穩定性和脅迫變化狀況進行分析,用 W統計量(W-statistic)作為ABC曲線方法的一個統計量,其公式為:
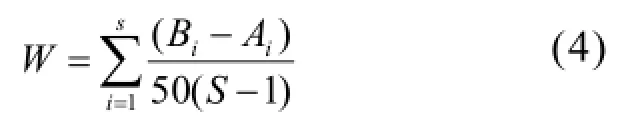
式中:Bi和Ai為ABC曲線中種類序號對應的生物量和豐度的累積百分比,S為出現物種數.當生物量優勢度曲線在豐度優勢度曲線之上時,W為正,反之W為負[24].
對上述公式計算、數據分析、方差檢驗、曲線繪制均通過 Microsoft Excel 2007和 SPSS 19.0、PRIMER 5.0軟件完成.
2 結果
2.1研究區水質環境特征
調查期間 N 區水溫(5.19±0.39)~(25.76± 0.21) ℃,鹽度(24.87±1.01)~(27.87±0.41);S區水溫(9.99±0.20)~(25.01±0.35)℃、鹽度(28.93±0.31)~(30.88±0.15)(圖2).N區DIN在春、秋、冬季全部超過第二類海水水質標準,S區水質均在第一類海水水質標準范圍內(圖3).
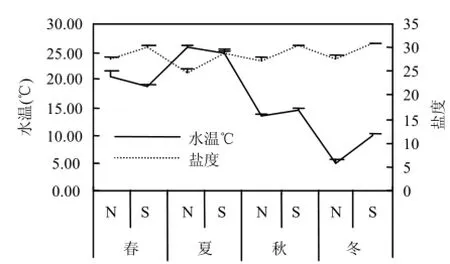
圖2 研究區溫、鹽季節變化Fig.4 Seasonal changes in temperature and salinity in the study area
t檢驗(成對雙樣本均值分析)結果表明,N、S區部分水環境指標在調查期間差異較為顯著(表 1),綜合考慮差值和相關系數的大小,DIN、COD、Cd、Hg、As含量N區較S區高,而S區鹽度全年高于N區,冬季水溫也高于N區.
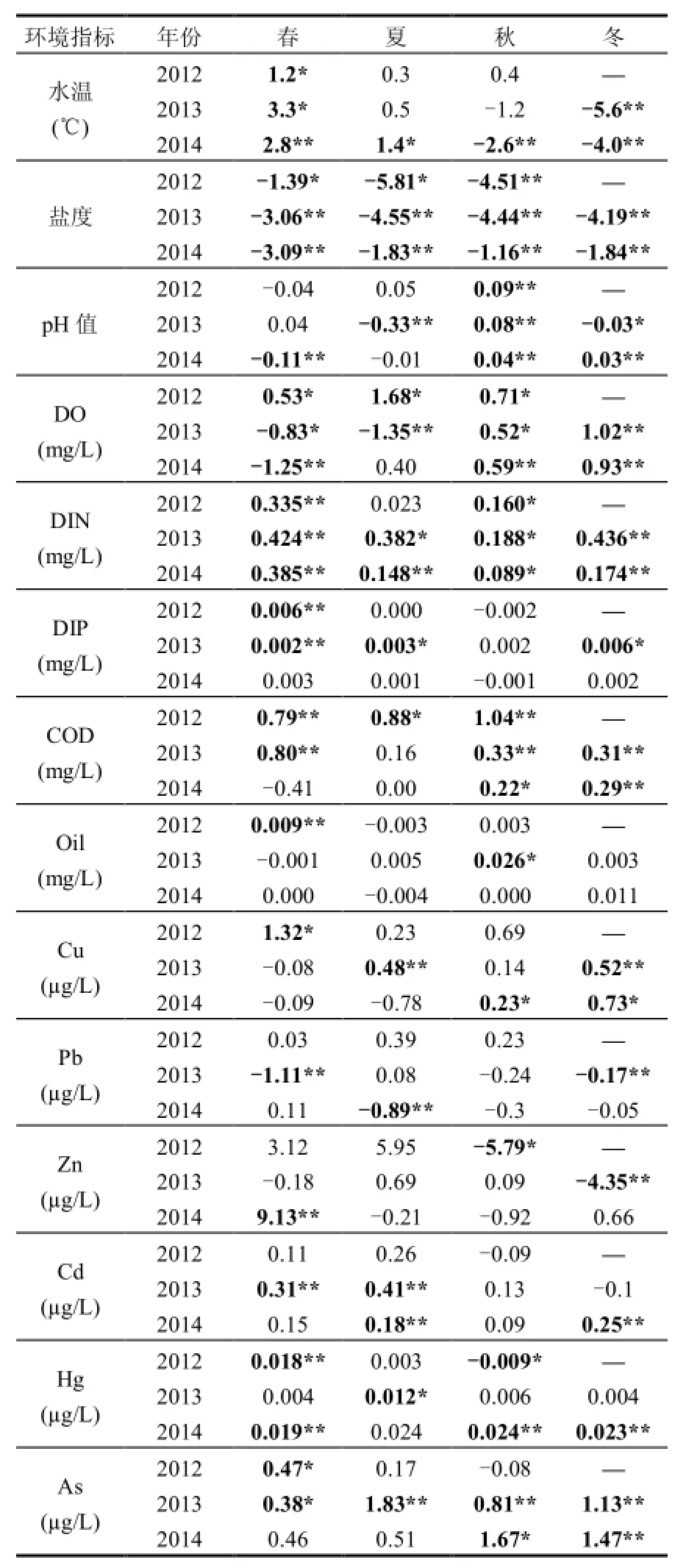
表1 研究區環境因子差值(N-S)Table 1 N-S difference of environmental factors in research area
海水營養狀況分析表明,研究N區屬于富營養化水平較高海域,S區基本屬于低營養水平(表2).這也符合本海域的自然屬性,N區北側鄰近遼河、大遼河及大、小凌河,陸域氮源輸入豐富.
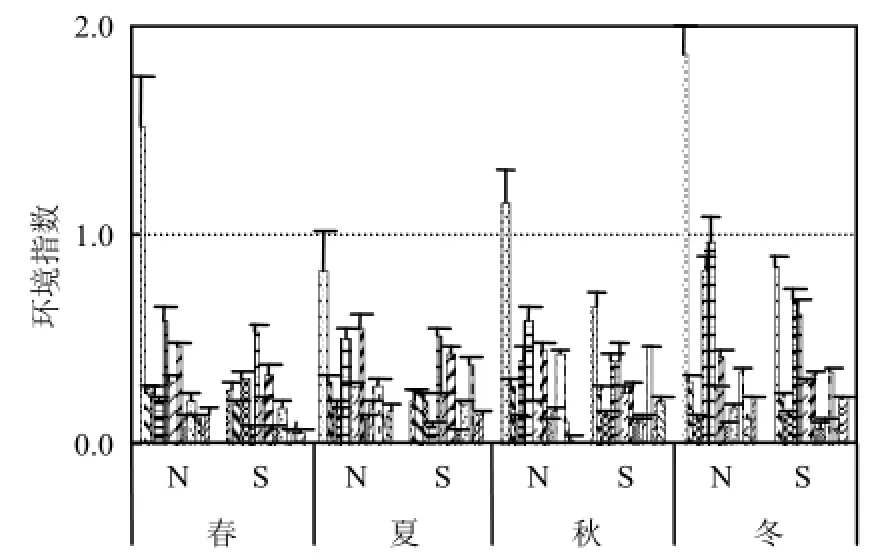
圖3 研究區環境因子季節變化Fig.4 Seasonal change of environmental factors in the study area
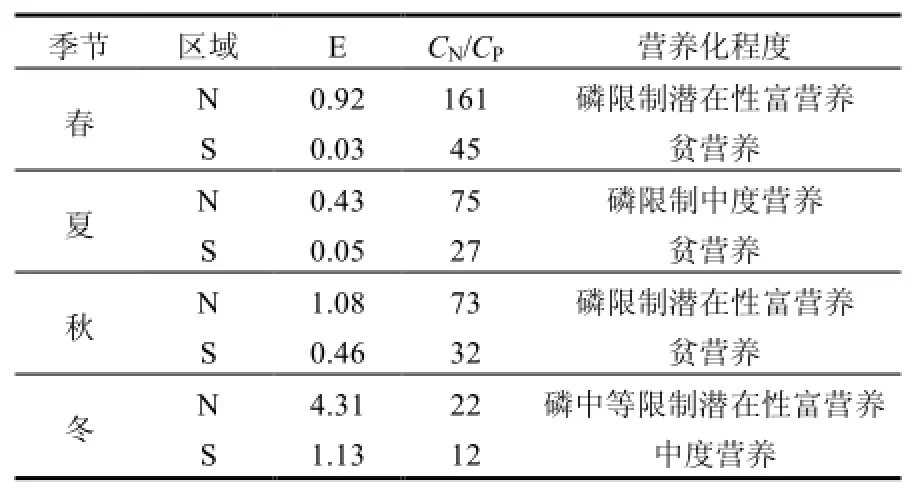
表2 研究區海水營養水平Table 1 Eutrophication level of marine in the study area
2.2優勢種及特有種組成
研究區(N、S) ESD<20 μm小粒徑組優勢種主要由中肋骨條藻(Skeletonema costatum)、具槽帕拉藻(Paralia sulcata)、角毛藻(Chaetoceros sp.)組成,20 μm<ESD<100 μm中粒徑組優勢種主要由格式圓篩藻(Coscinodiscus granii)、威氏圓篩藻(Coscinodiscus wailesii)組成,ESD>100 μm大粒徑組優勢種主要由星臍圓篩藻(Coscinodiscus asteromphalus)、虹彩圓篩藻(Coscinodiscus oculus-iridis)組成,N、S區優勢種差異不明顯(表3).調查還發現,長耳齒狀藻(Odontella aurita)、細弱圓篩藻在 N區出現頻率較高,而冰河擬星桿藻(Asterionellopsis glacialis)、新月菱形藻(Nitzschia closterium)、 離 心 列 海 鏈 藻 (Thalassiosira excentrica)、丹麥角毛藻(Chaetoceros danicus)、印度翼根管藻(Rhizosolenia alata f.indica)、蜂窩三角藻(Triceratium favus)、透明輻桿藻(Bacteriastrum hyalinum)、六幅輻裥藻(Actinoptychus senarius)、優美旭氏藻矮小變形(Schroderella delicatula)、掌狀冠蓋藻(Stephanopyxis palmeriana)、豪豬棘冠藻(Corethron hystrix)、巨圓篩藻(Coscinodiscus gigas)、 海 洋 原 多 甲 藻 (Protoperidinium oceanicum)、長角角藻(Ceratium macroceros)在S區出現頻率較高.
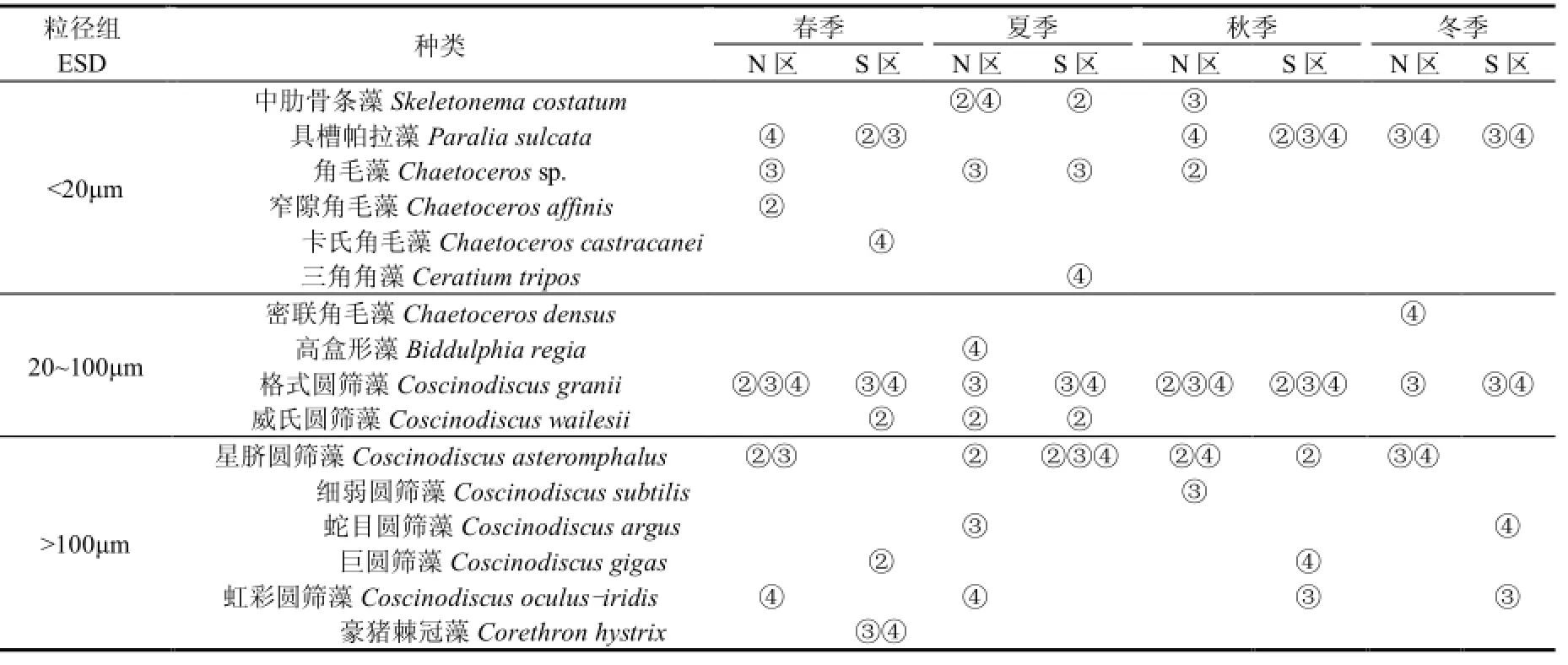
表3 研究區浮游植物各粒徑組第一優勢種Table 1 Phytoplankton size of the first dominant species in the study area
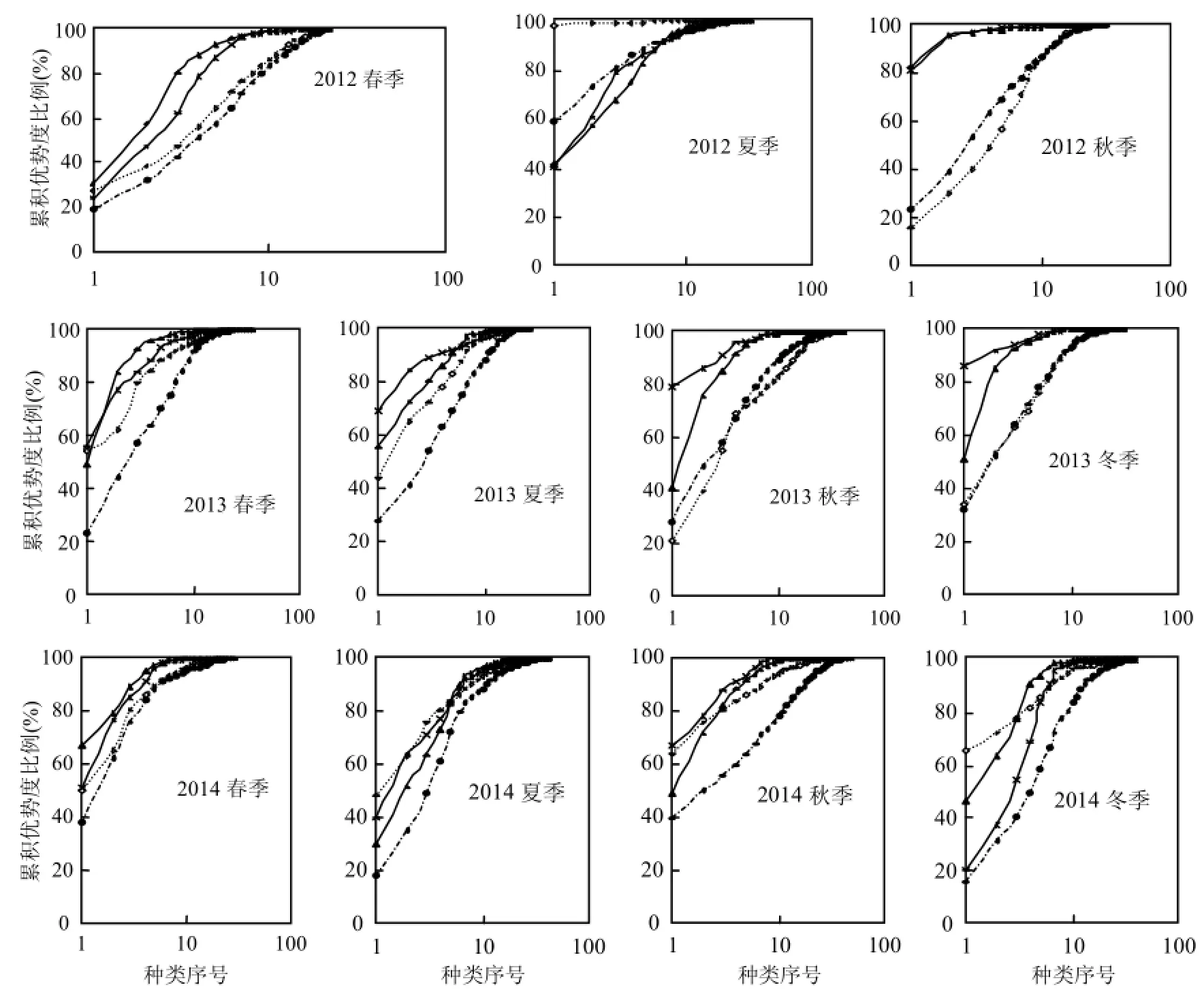
圖4 各季節浮游植物群落的ABC曲線以及W統計值Fig.4 ABC curves and W value of phytoplankton community in each season

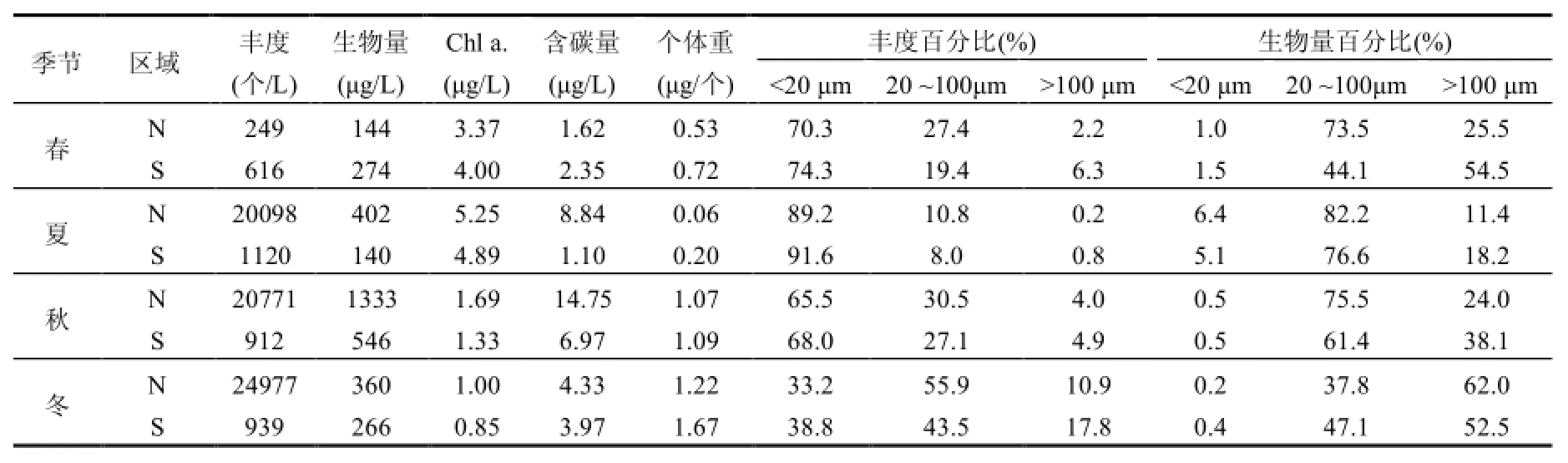
表4 研究區浮游植物粒徑組成Table 1 Phytoplankton size composition of the study area
2.3粒級結構變化特征
N、S區ESD全年分布在3~245μm,N區浮游植物總豐度、生物量、Chl a.、含碳量在夏、秋、冬季平均值均高于S區,而浮游植物平均個體重量全年S區均高于N區,表明S區整體上粒徑大于N區(表4).從各粒徑豐度、生物量組成可見,N、S區春、夏、秋季浮游植物豐度主要集中在 ESD<20μm小粒徑區域,冬季主要集中在20μm<ESD<100μm中粒徑區域;生物量春、夏、秋季主要集中在20μm<ESD<100μm中粒徑區域,冬季主要集中在 ESD>100μm大粒徑區域.S區ESD<20μm小粒徑組豐度全年高于 N區,而ESD>100μm大粒徑組生物量整體高于N區(除冬季外).豐度主要貢獻者為中肋骨條藻、具槽帕拉藻、薄壁幾內亞藻(Guinardia flaccida)、三角角藻、伏氏海線藻(Thalassionema frauenfeldii)、角毛藻等,而生物量主要貢獻者為粒徑較大的圓篩藻屬.
2.4ABC曲線特征
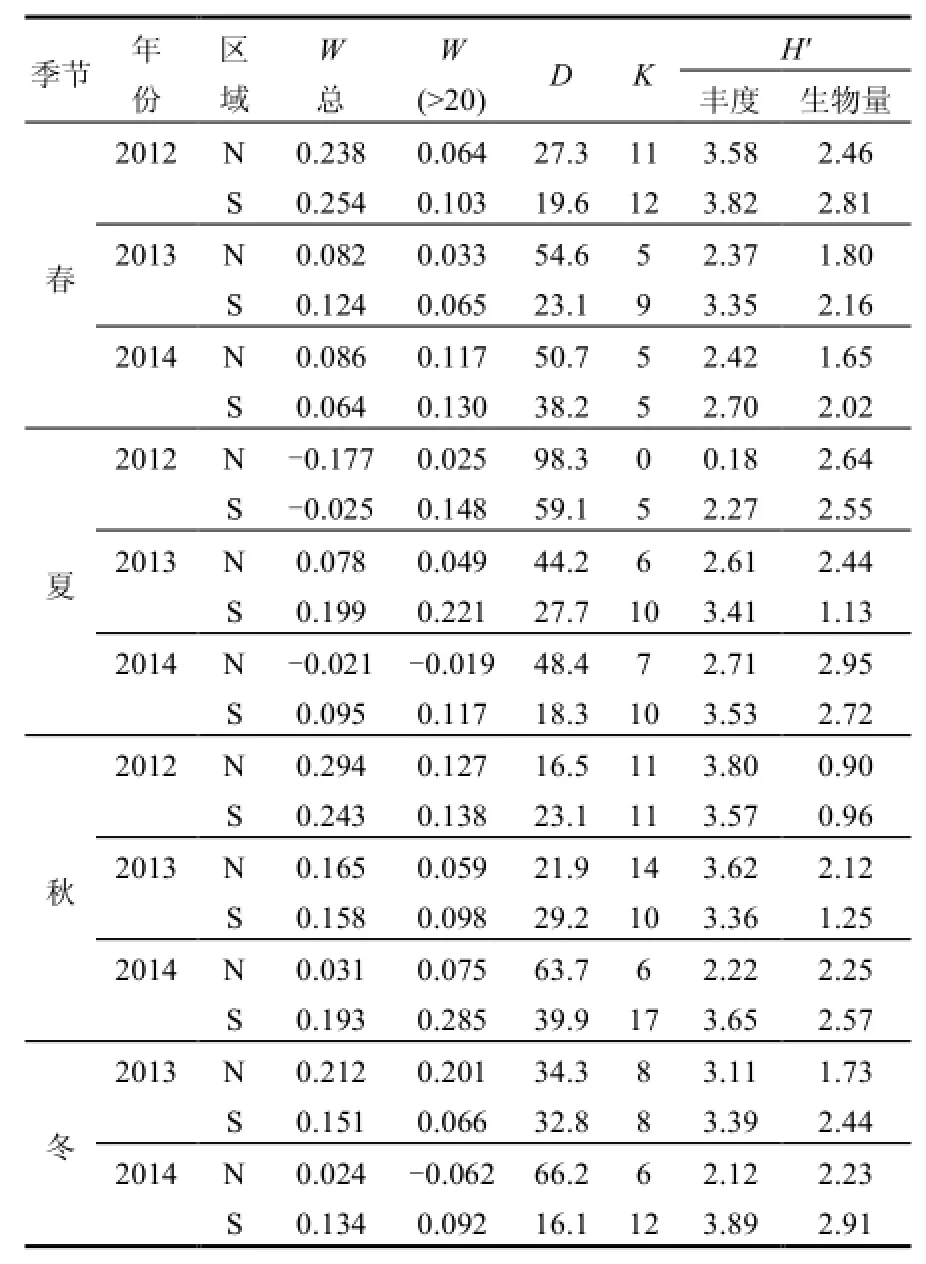
表5 ABC曲線及多樣性水平統計參數Table 1 The statistical parameter of ABC curves and diversity level
N、S區2012~2014年4個季節浮游植物群落的ABC曲線變化如圖4所示,總體而言,N、S區生物量優勢度曲線基本都在豐度優勢度曲線之上,僅2012年夏季N、S區豐度優勢度曲線居上,N區的2013年春季和2014年夏、秋、冬季2個優勢度曲線更替相交.說明N、S區春、秋、冬季浮游植物群落結構整體上比較穩定,僅夏季偏離平衡點.
從W總統計量來看(表5),數值基本為正,僅在夏季(2012年N、S區和2014年N區)出現負值.S區W統計量整體上高于N區,尤其ESD>20μm粒徑組(除2013年冬季外)均高于N區,說明S區群落結構整體上比N區穩定.
2.5多樣性水平
圖4中的豐度優勢度曲線即為k-優勢度曲線(k-dominance curve),用來檢驗種類豐度組成中包含的優勢種類的多少,可進行多樣性水平的分析與評價,曲線位置越低代表群落多樣性水平越高[25-26],而第一優勢度決定了k-優勢度曲線的起始位置.總體來看,N區浮游植物群落多樣性水平依次為秋>春>冬>夏,與多樣性指數(H′)統計的規律一致,而 S區多樣性水平依次為冬>春>秋>夏,與多樣性指數(H′)計算的結果基本一致(表5),總體而言,S區多樣性水平高于N區.
3 討論
本研究選取遼東灣南(S區)、北(N區)2個區域進行浮游植物群落及粒級結構分析,結果顯示不僅豐度與生物量上存在差異,其粒級結構和群落穩定性也存在明顯差異,說明環境因子的不同是其主要的影響因素.
3.1粒級結構的環境脅迫響應
N、S區中心相距僅150km,但鹽度、水溫、DIN、COD、Cd、Hg、As差異較為顯著,為避免浮游植物低鹽種類的影響,N區選取的10個站位鹽度均在20以上,S區選取的10個站位鹽度均在28以上,調查中N區也未檢出淡水種類.由于N區水深相對較淺,同期調查水溫在春、夏季略高,秋、冬季略低.重金屬 Cd、Hg、As含量在兩區之間雖有差異,但差值較小,所測閾值也均在第二類海水水質標準范圍內,對浮游植物的群落結構影響較小.而營養鹽水平是影響浮游植物粒度的主要因素,調查發現,N區富營養化水平較高,S區則為寡營養水平.
研究發現,N區浮游植物生物量在 20μm<ESD<100μm中粒徑區域較S區有明顯優勢,而在ESD>100μm大粒徑區域比例卻相對較低.由于是網采樣品,ESD<20μm粒徑組主要是由小粒徑的鏈狀或桿狀的骨條藻、帕拉藻、角毛藻、菱形藻等組成,導致低營養水平的S區ESD<20μm小粒徑組豐度組成全年高于N區,但在以圓篩藻等短圓柱形為主的ESD>100μm大粒徑組生物量整體高于N區.本研究于2014年夏、秋季對同期采集的水樣品進行了葉綠素 a分級測定(20、3、0.22μm的濾膜逐級過濾),結果表明,S區<20μm葉綠素a組成(夏88.4%、秋76.5%)均高于N區(夏49.2%、秋72.6%).這與Grover[27]研究結果相似,高營養鹽環境更能促進小粒徑球形細胞和大粒徑細長型細胞的繁殖.許多研究也表明,寡營養海域,微微型浮游植物(0.2~3μm)是初級生產力的主要貢獻者,營養豐富的水域,初級生產力主要由粒徑較大的小型浮游植物(20~200μm)貢獻[2].
另外,溫度也是影響浮游植物粒徑分布的重要因素,在溫度較低的南極普里茲灣,各粒級浮游植物對初級生產力的貢獻分別為小型浮游植物69%、微型浮游植物 21%、微微型浮游植物10%[28].本研究在冬季水溫較低環境下,浮游植物生物量主要由ESD>100 μm大粒徑組貢獻,也驗證了低水溫環境下更適于大粒徑浮游植物生存的結論.
由于本研究針對的浮游植物粒級研究是網采樣品,不可避免漏掉大部分微型及微微型浮游植物,可能無法體現各粒級浮游植物的實際分布特征,但本研究利用同種方法分析的2個區域,可以反映不同環境脅迫下網采浮游植物的粒級結構響應規律.
3.2群落結構的穩定性評估
ABC曲線法具有生態學中r-選擇和k-選擇策略的理論基礎,可比較分析不同時期,不同脅迫情況下浮游植物群落的響應[29-31].
春季,由于光照充足、溫度適宜和營養鹽的蓄積給浮游植物生長帶來了有利條件,使得中肋骨條藻、具槽帕拉藻、角毛藻等以 r-對策者為主的小個體細胞迅速繁殖,生物量曲線暫時位于豐度曲線之上;夏季,海水營養鹽大部分被浮游植物消耗,由于降雨量較小,營養鹽沒有及時從陸源得到補充(N、S區2012年和2014年無機氮檢測含量相對其他季節偏低),導致群落結構偏離平衡點,生物量曲線位于豐度曲線之下;秋季,由于營養鹽消耗大于補充,以r-對策者為主的小個體細胞逐漸被 k-對策者的圓篩藻屬等大個體細胞演替,群落逐漸恢復到平衡點;到了冬季,主要受水溫影響,群落主要由k-對策者的圓篩藻屬完成反饋調節.
浮游植物群落結構穩定性評估結果顯示,營養化程度較低的S區群落結構整體上比N區穩定,與生物多樣性評估結果基本一致.因此,可以嘗試利用ABC曲線法定量評估浮游植物群落結構穩定性狀況,而對于網采樣品,統計 ESD>20 μm粒徑組的W統計量評估群落結構穩定性相對更加合理和符合實際情況.
另外,研究區浮游植物類群主要以硅藻為主,甲藻為輔.N區硅/甲藻豐度(生物量)比例在夏、秋、冬季平均值均高于S區,而春季比例接近,說明甲藻更適于S區的生存環境.因為與硅藻相比,甲藻更耐受較低的營養鹽,但需要較高的溫度和較強的光照.
4 結論
4.1遼東灣海域網采浮游植物小粒徑優勢種主要由中肋骨條藻、具槽帕拉藻、角毛藻組成,中粒徑優勢種主要由格式圓篩藻、威氏圓篩藻組成,大粒徑優勢種主要由星臍圓篩藻、虹彩圓篩藻組成,由于種群分布不均,僅分布于南部區域的物種數量相對于只分布在北部區域的物種數量偏多.
4.2低營養水平的南部區域浮游植物粒徑整體上大于北部區域,雖小粒徑組豐度比例較高,但大粒徑組生物量比例卻高于北部區域,群落結構整體上比北部區域穩定.寡營養水域以及較低的水溫環境更適于k-對策者的大粒徑浮游植物生存.
4.3可以嘗試利用網采ESD>20μm粒徑組的W統計量作為浮游植物群落結構穩定性的定量評估指標之一.
[1] 馬艷娥,柯志新,黃良民,等.夏冬兩季大亞灣典型海域浮游植物粒級結構特征 [J]. 熱帶海洋學報, 2013,32(3):40-46.
[2] 楊茹君,王修林,韓秀榮,等.海洋浮游植物粒徑組成及其生物粒徑效應研究 [J]. 海洋科學, 2003,27(11):5-9.
[3] 周林濱,譚燁輝,黃良民,等.水生生物粒徑譜/生物量譜研究進展[J]. 生態學報, 2010,30(12):3319-3333.
[4] 李清雪,陶建華.應用浮游植物群落結構指數評價海域富營養化[J]. 中國環境科學, 1999,19(6):548-551.
[5] 孫 軍,劉東艷,錢樹本.浮游植物生物量研究I.浮游植物生物量細胞體積轉化法 [J]. 海洋學報, 1999,21(2):75-85.
[6] 左 濤,王 俊,金顯仕,等.春季長江口鄰近外海網采浮游生物的生物量譜 [J]. 生態學報, 2008,28(3):1174-1182.
[7] 左 濤,王 俊,唐啟升,等.秋季南黃海網采浮游生物的生物量譜 [J]. 海洋學報, 2008,30(5):71-80.
[8] 吳金浩,徐雪梅,楊 爽,等.2007年春、秋遼東灣北部營養鹽及影響因素 [J]. 海洋科學進展, 2012,30(4):477-486.
[9] 周遵春,馬志強,薛 克,等.對遼東灣夜光藻赤潮和叉狀角藻赤潮成因的初步研究 [J]. 水產科學, 2002,21(2):9-12.
[10] 欒 莎,宮相忠,雙秀芝,等.2009年春季遼東灣網采浮游植物群落結構 [J]. 海洋科學, 2012,36(5):57-64.
[11] 康元德,王 俊.渤海浮游植物的數量分布和季節變化 [J]. 海洋水產研究, 1991,12(12):31-54.
[12] 俞建鑾,李瑞香.渤 海,黃海浮游植物生態的研究 [J]. 黃渤海海洋, 1993,11(3):52-59.
[13] 王 俊,康元德.渤海浮游植物種群動態的研究 [J]. 海洋水產研究, 1998,19(1):51-59.
[14] 宋 倫,周遵春,王年斌,等.遼東灣浮游植物多樣性與海洋環境的關系 [J]. 海洋環境科學, 2007,26(4):365-368.
[15] 耿文華,陳繼淼,馮劍豐,等.遼河浮游植物群落及生物多樣性基準驗證 [J]. 中國環境科學, 2014,34(1):239-245.
[16] 宋 倫,王年斌,宋永剛,等.遼寧近岸渾濁海域網采浮游生物的粒徑結構特征 [J]. 應用生態學報, 2013,24(4):900-908.
[17] GB/T 12763-2007 海洋調查規范 [S].
[18] GB 17378-2007 海洋監測規范 [S].
[19] 孫 軍,劉東艷.中國海區常見浮游植物種名更改初步意見 [J].海洋與湖沼, 2002,33(3):271-286.
[20] Eppley RW, Reid F M, Strickland J D. The ecology of the plankton off La Jolla, Callifornia, in the period April through September 1967Par Ⅱ. Estimates of phytoplankton crop size, growth rate and primary production [J]. Bulletin of the Scripps Institution of Oceanography, 1970,17:33-42.
[21] Pinkas L, Oliphant M S, Iverson I L K. Food habits of albacore, bluefin tuna, and bonito in California waters [J]. California Department of Fish and Game Fish Bulletin, 1971,152:1-105.
[22] 郭衛東,章小明,楊逸萍,等.中國近岸海域潛在性富營養化程度的評價 [J]. 臺灣海峽, 1998,17(1):64-70.
[23] GB3097-1997 海水水質標準 [S].
[24] Warwick R M. A new method for detecting pollution effects on marine macrobenthic communities [J]. Marine Biology, 1986, 92(4):557-562.
[25] Ellis J R, Rogers S I, Freeman S M. Demersal assemblages in the Irish Sea, St George's Channel and Bristol Channel [J]. Estuarine, Coastal and Shelf Science, 2000,51(3):299-315.
[26] Platt H M, Shaw K M, Lambshead P J D. Nematode species abundance patterns and their use in the detection of environment perturbations [J]. Hydrobiologia, 1984,118(1):59-66.
[27] Grover JP. Influence of cell shape and size on algal competitive ability [J]. Journal of Phycology, 1989,25:402-405.
[28] 劉子琳,蔡昱明,陳中元,等.1998/1999年南極夏季普里茲灣及北部海區葉綠素 a和初級生產力的分布特征 [J]. 極地研究, 2002,14(1):12-21.
[29] Yemane D, Field J G, Leslie R W. Exploring the effects of fishing on fish assemblages using Abundance Biomass Comparison (ABC) curves [J]. ICES Journal of Marine Science, 2005,62(3):374-379.
[30] 曲方圓,于子山,隋吉星,等.豐度生物量比較法應用局限性 [J].海洋科學, 2009,33(6):118-121.
[31] 宋 倫,王年斌,楊國軍,等.鴨綠江口及鄰近海域生物群落的脅迫響應 [J]. 生態學報, 2013,33(9):2790-2802.
The stress response of net phytoplankton biomass size structure in Liaodong Bay.
SONG Lun1,2*, SONG Guang-jun1,2, WANG Nian-bin1,2, ZHAO Hai-bo1,2, TIAN Jin1,2, YANG Shuang1,2, DU Jing1,2(1.Liaoning Ocean and Fisheries Science Research Institute, Dalian 116023, China;2.Liaoning Ocean Environment Monitoring Station, Dalian 116023, China).
China Environmental Science, 2015,35(9):2764~2771
Based on net collection of phytoplankton samples in Liaodong Bay in all four seasons during 2012~2014, the distribution pattern of size structure of phytoplankton and its response to environmental stress were analyzed in the northern and southern areas of the bay. The stability of the phytoplankton community was estimated and was found to be dominated by small species in spring, summer and autumn, and by medium-sized species in winter. Phytoplankton biomass was controlled by medium-sized species in spring, summer and autumn, and by large species in winter. In general, phytoplankton sizes in the southern area were greater than those in the northern area. Large phytoplankton species were much more adaptive to oligotrophic water and a colder environment. The phytoplankton community in spring, autumn and winter was relatively stable; however, it deviated from equilibrium in summer. Phytoplankton community stability in the southern area was slightly better than in the northern area. The W statistical value of the group of large species can be used as an evaluation index of phytoplankton community stability.
phytoplankton;size structure;ABC curves;bionomic strategy;ecological stability;Liaodong Bay
X171.1;Q89;P735
A
1000-6923(2015)09-2764-08
2015-01-11
遼寧省自然科學基金資助項目(2014020182);遼寧省海洋與漁業科研項目(201415);國家自然科學基金資助項目(31400406)
*責任作者, 副研究員, songlun@lnshky.com
宋 倫(1980-),男,遼寧大連人,副研究員,碩士,主要從事海洋生物生態學研究.發表論文30余篇,專著2部.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
哲學評論(2021年2期)2021-08-22 01:53:34
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
