
酵母穿梭質粒pXZ200的構建
2015-09-10 21:06:00王珊珊徐海洋王世明
江蘇農業科學 2015年8期
王珊珊+徐海洋+王世明


摘要: 利用釀酒酵母高效的同源重組能力,通過PCR擴增技術,將復制子pBBR1片段、KanR(卡納霉素抗性)片段、酵母復制子2 μ片段這3個兩端含有30 bp同源臂的片段電轉入釀酒酵母,通過同源重組獲得宿主更具廣譜性的革蘭氏陰性細菌-啤酒酵母穿梭質粒pXZ200。經檢測,pXZ200能夠在惡臭假單胞桿菌KT2440和大腸桿菌中穩定復制。
關鍵詞: 釀酒酵母;同源重組;穿梭質粒;PCR技術;pXZ200
中圖分類號: Q936 文獻標志碼: A
文章編號:1002-1302(2015)08-0031-03
分子生物學中,利用DNA重組技術使宿主細胞獲得某種特殊能力是非常重要而且被廣泛利用的方法。然而,這種方法在體外需要經多步的酶切和酶連,很大程度上依賴于載體上限制性核酸內切酶的酶切位點,步驟繁瑣,所連接片段一般限于幾個kbp大小,很少超過10 kbp,但同源重組的新方法克服了這種限制。
釀酒酵母細胞內有著較其他生物更高效的同源重組修復雙鏈DNA斷裂機制,是研究同源重組機制和應用同源重組技術的主要模式生物 [1-2]。在釀酒酵母系統中,15 bp的同源序列就可以介導同源重組的發生,獲得重組克隆 [3];當兩端的同源序列為30 bp時,有效同源重組的頻率可接近80% [4-5]。PCR介導重組技術是指通過人工合成寡核苷酸單鏈作為PCR引物,用PCR擴增方法得到重組片段的兩端帶有重組所需同源臂外源的DNA片段 [6-7],是目前應用最為廣泛的一種重組技術 [4,8-10],這種方法與傳統酶切酶連的基因克隆方法相比,具有很多優點:省去了多步的酶切酶連,操作更為簡單快捷;不依賴于限制性酶切位點的存在,不會引入多余的堿基;通過合成特定的同源序列,目的基因可以精確地定位在靶位點上;解決了大片段的基因克隆問題 [11-12]。體外將合成的DNA片段利用釀酒酵母的重組機制,在體內完成多個DNA片段的一步整合已被廣泛運用。1970—2008年,DNA合成片段已經從75 bp增加到583 kbp [13]。1970年,Khorana等成功合成丙氨酸轉運RNA的完整基因,這是第一次實現化學合成可遺傳序列 [14];2008年,Venter研究組合成了由101個DNA片段構建、長度為583 kbp的DNA [15],該DNA通過體外合成25個具有同源臂的DNA片段,然后利用酵母重組機制合成基因組 [16];2009年,Shao等在酵母中一步裝配完成含有8個基因的生化途徑,含有這個生化途徑的重組菌能夠利用木糖產生玉米黃質 [17]。
帶有完整2μ質粒序列的質粒pXZ198是一個既能在酵母中復制表達、又能在大腸桿菌中復制表達的穿梭質粒,在研究中被廣泛應用。但是,pXZ198質粒只含有大腸桿菌的復制子Coli E1,而不能在其他原核細胞中復制表達,而 pBBR1MCS5 卻是能夠在多種革蘭氏陰性菌中穩定復制表達的質粒 [18]。本試驗利用釀酒酵母的體內高效重組系統,將廣譜復制子pBBR1、卡那霉素抗性基因KanR、酵母復制子2μ序列重組成一新的質粒pXZ200,經驗證,該質粒可以在大腸桿菌及惡臭假單胞桿菌KT2440中穩定復制。
1 材料與方法
1.1 材料
1.1.1 主要儀器 恒溫培養箱、PCR儀、搖床、低溫離心機、冰箱、制冰機、無菌操作臺、滅菌鍋、電轉杯、電轉儀、多功能酶標儀等。
1.1.2 主要試劑 DraⅠ限制性內切酶、DNA回收試劑盒、1 mol/L 山梨醇、氨芐抗生素、慶大霉素、卡那霉素、博來霉素等。
1.1.3 主要菌種和質粒 釀酒酵母S288C、大腸桿菌DH5α、惡臭假單胞桿菌KT2440、含有完整2 μ序列及博來霉素抗性基因片段pXZ 的pXZ198質粒、含有KanR片段的pET29a質粒、含有在多種原核細胞中能復制的廣譜復制子pBBR1的pBBR1MCS5質粒。
1.1.4 培養基 YPD培養基:2%蛋白胨、1%酵母粉、2%葡萄糖,pH值為7.0,115 ℃滅菌20 min。LB培養基:1%蛋白胨、0.5%酵母粉、1%氯化鈉,pH值為7.0,121 ℃滅菌 15 min。固體培養基是在液體培養基的基礎上加入1.5%瓊脂。
1.2 方法
1.2.1 打靶片段的準備 在-80 ℃接種pXZ198 DH5α至含有100 μg/mL氨芐抗生素、接種pET29a DH5α至含有 50 μg/mL Kan、接種pBBR1MCS5 DH5α至含有30 μg/mL慶大霉素的LB試管中,37 ℃ 200 r/min過夜培養;分別取約 3 mL 菌液,提取質粒 [19]pXZ198、pET29a、pBBR1MCS5,用PCR擴增方法獲得兩端含有30 bp同源臂的打靶片段,打靶片段PCR對應的引物和模板見表1;每個片段擴增約 120 μL,按照康寧生命科學的試劑盒回收各個片段;回收的各個片段進行0.75%瓊脂糖電泳并測各片段的濃度。
1.2.2 感受態細胞的制備 參照文獻[17]的方法制作酵母電轉感受態、KT2440電轉感受態以及采用大腸桿菌DH5α一步法轉化感受態細胞 [20]。
1.2.3 電轉重組及初步篩選 取50 μL酵母感受態細胞置于預冷的0.2 cm的電轉杯中,加入打靶片段pXZ、pBBR1、KanR各5 μL,混勻;擦干電轉杯上的水,放入電轉化儀的樣品槽中,以1.5 kV進行電轉化;電轉結束,迅速將1 mL已滅菌的YPD培養基加入到電轉杯中;在無菌操作臺上,將電轉杯中液體轉移到1.5 mL已滅菌的離心管中,30 ℃ 120 r/min復蘇1 h;取100 μL涂布于含有200 μg/mL博來霉素的YPD平板上,共涂10個平板;平板放置于30 ℃恒溫培養箱中培養24~36 h。
1.2.4 酵母重組子的驗證及pXZ200在酵母中的穩定性 將平板上長出的單菌落隨機挑選出3個,并編號為1、2、3,接種至含200 μg/mL博來霉素的YPD試管中,30 ℃ 200 r/min培養24 h,以Kan-R、pBBR1-F為引物對重組子進行PCR鑒定。經PCR鑒定正確的重組子轉接至含有200 μg/mL博來霉素的50 mL YPD培養基中,30 ℃ 200 r/min培養24 h以提取質粒。由于酵母菌是真核生物,細胞的破碎難于大腸桿菌。試驗采用液氮研磨的方法進行細胞破碎,用酚氯仿反復抽提進行質粒的提取。質粒以pXZR-F、pXZR-R為引物,送至上海桑尼生物科技有限公司測序,測序序列與pBBR1、KanR片段的序列進行比對;測序正確的重組子轉接至YPD試管中,30 ℃ 200 r/min過夜培養,反復轉接1周并提質粒;對所提質粒進行DraⅠ酶切,酶切體系為10 μL:pXZ200 3 μL、DraⅠ和10×M Bf各1 μL、無菌ddH2O 5 μL;0.75%瓊脂糖電泳檢測質粒的正確性。
1.2.5 構建質粒在宿主細菌中的復制 將2 μL質粒轉入 [21]50 μL DH5α一步法感受態細胞中;轉化后加入600 μL的LB復蘇1 h;取100 μL涂布于含有50 μg/mL卡那霉素的LB平板上,平板放置于37 ℃恒溫培養箱中過夜培養;挑取平板上長出的單菌落,接種到含有50 μg/mL卡那霉素的LB試管中,37 ℃ 200 r/min過夜培養,提取質粒;質粒正確的菌轉接至LB試管中,37 ℃ 200 r/min過夜培養,反復轉接1周并提取質粒,對所提的質粒進行DraⅠ酶切;0.75%瓊脂糖電泳檢測質粒的正確性。以同樣方法在假單胞桿菌KT2440中檢測pXZ200的穩定性。
2 結果與分析
2.1 pXZ200質粒的構建
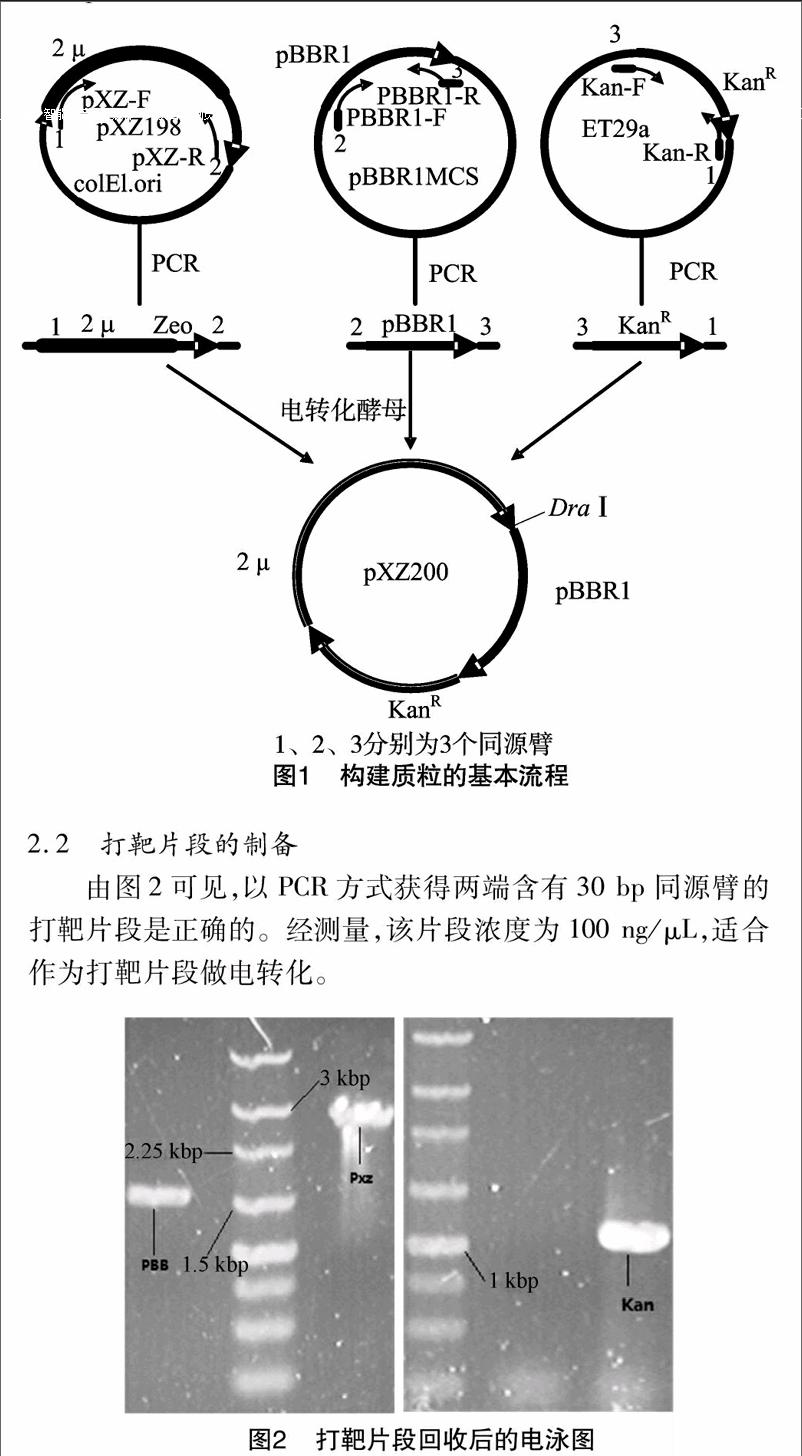
質粒構建的基本流程見圖1。
2.2 打靶片段的制備
由圖2可見,以PCR方式獲得兩端含有30 bp同源臂的打靶片段是正確的。經測量,該片段濃度為100 ng/μL,適合作為打靶片段做電轉化。
2.3 對重組子的驗證及 在酵母中的穩定性。
結果表明,10個含200 μg/mL博來霉素的YPD平板上都長出單菌落,每個平板平均長出20個單菌落;對菌落進行驗證統計,重組效率約為70%;對其中3個重組子進行PCR驗證,由圖3可見,KanR和pBBR1 2個片段的長度大小約為2.5 kbp,1號和2號單菌落是正確的。將2號菌提出的質粒以pXZR-F、pXZR-R為引物測序,經與pBBR1及KanR片段的序列進行比對,符合度達99%,證明該質粒構建正確。
2.4 構建質粒 在假單胞桿菌和大腸桿菌中的表達及穩定性
從酵母菌提取的質粒 轉入大腸桿菌和假單胞桿菌KT2440中,提質粒(3 mL菌液)驗證都正確(圖4)。將含有質粒的大腸桿菌、假單胞桿菌KT2440及驗證正確的酵母重組子分別連續轉接1周,所提取的質粒經DraⅠ酶切。由圖5可見,從大腸桿菌及假單胞桿菌KT2440中提取的質粒和從酵母中提取的質粒相比,大小一致且轉接1周后質粒的量沒有變化,酶切后質粒大小為5.3 kbp。說明 能夠在酵母菌、大腸桿菌DH5α和假單胞桿菌KT2440中穩定復制。
3 結論
傳統的質粒構建需要找到一個合適的載體,將目的片段和載體片段在相同的酶切后進行酶連,再轉入到宿主菌中鑒定,如果構建的質粒需要插入多個片段,則工作量很大。本試驗采用的方法比較簡單,只需要在PCR獲取目的片段時加入同源臂,然后將所有目的片段混勻,電轉到酵母感受態細胞中再進行篩選驗證。與傳統方法相比,這種方法對多片段質粒的構建簡單且易于操作。試驗所構建的新質粒 能夠在酵母菌及大腸桿菌、假單胞桿菌中穩定復制。
參考文獻:
[1] Szostak J W,Orr-Weaver T L,Rothstein R J,et al. The double-strand-break repair model for recombination[J]. Cell,1983,33 (1):25-35.[HJ1.7mm]
[2][JP2]Bhatia P K,Wang Z G,Friedberg E C. DNA repair and transcription[J]. Current Opinion Genetics Development,1996,6(2):146-150.
[3]Judd S R,Petes T D. Physical lengths of meiotic and mitotic gene conversion tracts in Saccharomyces cerevisiae[J]. Genetics,1988,118(3):401-410.
[4]Nickoloff J A,Hoekstra M F. Double-strand break and recombinational repair in Saccharomyces cerevisiae[M]//Nickoloff J A,Hoekstra M F. DNA damage and repair,vol. 1:DNA repair in prokaryotes and lower eukaryotes. Totowa,NJ:Humana Press Inc,1998:335-362.
[5]Willis K K,Klein H L. Intrachromosomal recombination in Saccharomyces cerevisiae:reciprocal exchange in an inverted repeat and associated gene conversion[J]. Genetics,1987,117(4):633-643.
[6][JP3]Kramer K M,Brock J A,Bloom K,et al. Two different types of double- strand breaks in Saccharomyces cerevisiae are repaired by similar RAD52-independent,nonhomologous recombination events[J]. Molecular and Cellular Biology,1994,14(2):1293-1301.
[7]Orr-Weaver T L,Szostak J W. Yeast recombination:the association between double-strand gap repair and crossing-over[J]. Proceedings of the National Academy of Sciences of the United States of America,1983,80(14):4417-4421.
[8][JP2]Schwacha A,Kleckner N. Identification of double Holliday junctions as [JP3]intermediates in meiotic recombination[J]. Cell,1995,83(5):783-791.
[9]Nasmyth K A.Molecular genetics of yeast mating type[J]. Annual Review Genetics 1982,16:439-500.
[10] Thaler D S,Stahl F W. DNA double-chain breaks in recombination of phage lambda and of yeast[J]. Annual Review of Genetics,1988,22:169-197.
[11]Nassif N,Penney J,Pal S,et al. Efficient copying of nonhomologous sequences from ectopic sites via P-element-induced gap repair[J]. Molecular and Cellular Biology,1994,14(3):1613-1625.
[12]Mcgill C,Shafer B,Strathern J. Coconversion of flanking sequences with homothallic switching[J]. Cell,1989,57(3):459-467.
[13]Mueller S,Coleman J R,Wimmer E. Putting synthesis into biology:a viral view of genetic engineering through de novo gene and genome synthesis[J]. Chemistry & Biology,2009,16(3):337-347.
[14] Gibson D G. Synthesis of DNA fragments in yeast by one-step assembly of overlapping oligonucleotides[J]. Nucleic Acids Research,2009,37(20):6984-6990.
[15]Gibson D G,Benders G A,Andrews-Pfannkoch C,et al. Complete chemical synthesis,assembly,and cloning of a mycoplasma genitalium genome[J]. Science,2008,319(5867):1215-1220.
[16]Gibson D G,Benders G A,Axelrod K C,et al. One-step assembly in yeast of 25 overlapping DNA fragments to form a complete synthetic mycoplasma genitalium genome[J]. Proc Natl Acad Sci USA,2008,l05(51):20404-20409.
[17]Shao Z Y,Zhao H,Zhao H M. DNA assembler,an in vivo genetic method for rapid construction of biochemical pathways[J]. Nucleic Acids Research,2009,37(2):e16.
[18]Kovach M E,Elzer P H,Steven Hill D,et al. Four new derivatives of the broad-host-range cloning vector pBBR1MCS,carrying different antibiotic-resistance cassettes[J]. Gene,1995,166 (1):175-176.
[19]任大勇,李 昌,秦艷青,等. 小提質粒快速排除假陽性克隆的新方法[J]. 吉林農業大學學報,2011,33(2):185-188.
[20]涂知明,陳明潔,何光源,等. 三種大腸桿菌高效感受態的制備及轉化[J].
