NaN3處理馬齒莧種子的最適劑量與葉綠素熒光輔助篩選的方法
2015-10-20 00:23:38王瑞聰等
江蘇農業科學 2015年9期
關鍵詞:生長
王瑞聰等
摘要:以不同劑量NaN3處理馬齒莧種子,從種子萌發、植株生長、光合性能的角度確定最適NaN3處理劑量,并分析以葉綠素熒光動力學方法輔助篩選的可行性。結果表明,1.0 mmol/L NaN3處理8 h或0.5 mmol/L NaN3處理12 h均可顯著提高種子萌發率和萌發質量,而更高濃度、更長時間的處理則使萌發率和萌發質量逐漸下降;根據長勢初篩后的各處理組種子萌發苗,其株高均高于對照或與對照相當,單葉面積(2.0 mmol/L NaN3 8 h處理組除外)則低于對照;各處理組葉片PSⅡ的潛在和實際光化學效率普遍高于對照(其中2.0 mmol/L NaN3 8 h處理組最高),但除2.0 mmol/L NaN3 8 h處理組外,各組對高光強的抵御能力,特別是快速保護能力均有所下降。2.0 mmol/L NaN3 8 h處理組中馬齒莧種子獲得植株的株型最高、單葉面積最大、潛在和實際光化學活性最強、抵御強光脅迫能力最強。可見,以2.0 mmol/L NaN3處理8 h是馬齒莧種子的最適處理條件,而采用葉綠素熒光動力學分析技術可實現對當代植株光合能力、高光強抗性的無損傷分析,極有利于提高篩選效率。
關鍵詞:馬齒莧;NaN3;種子萌發;生長;葉綠素熒光
中圖分類號:S335;Q945.34文獻標志碼: A文章編號:1002-1302(2015)09-0224-05
馬齒莧(Portulaca oleracea)別稱馬齒菜、長生菜、五行草等,為馬齒莧科(Portulacease)馬齒莧屬(Portulaca)的一年生肉質草本植物。馬齒莧的蛋白質、總黃酮含量高,氨基酸種類齊全,富含ω-3不飽和脂肪酸、鉀元素,并含有去甲腎上腺素、褪黑激素等多種保健營養成分,具有改善血液循環、提高人體免疫力、防治心血管疾病、抑制微生物生長等多重功效[1-3]。此外,馬齒莧對高溫、干旱、高濕、高鹽、重金屬污染等逆境的抵抗能力強大,且是優良的生態修復植物[4]。馬齒莧集保健、食用、藥用、生態價值于一身,具有進一步研究開發的意義。
我國馬齒莧主要以野生狀態存在,其株形匍匐、分枝多、抗白粉病能力弱、主要生長于盛夏高溫季節等原因限制了馬齒莧的開發應用及推廣。為改善這一現狀,須篩選并培育出株形高大、生長旺盛、適應范圍廣的馬齒莧新種質。誘變育種是獲得新種質最常見的方法,而由于化學誘變操作簡便、對設備依賴程度低等原因,使其在該領域占有重要地位[5]。化學誘變多為隱性突變,遺傳變異須在誘變后的子一代甚至子二代中才能被檢出[6-7],使得誘變育種的最大工作量集中于篩選,而尋找簡便有效的篩選方法始終是誘變育種的主題和難題。傳統研究中多數選擇半致死劑量(LD50)作為誘導最適劑量[8],或以子一代、子二代的葉色變化代表誘變率來篩選最適劑量,并直接對目標性狀進行篩選。近年來得以發展的RAPD、SRAP多態性等分子生物學方法,對于從遺傳學角度輔助確立最適劑量,以及確定是否發生了可穩定的突變發揮著越來越大的作用。但由于致死不與誘變直接相關,葉色異常、DNA指紋圖譜等不與目標優良性狀直接相關,且RAPD、SRAP等方法步驟繁瑣、耗時耗材,致使誘變劑量的確定及后續突變株的篩選成為誘變育種中最關鍵、最耗時的步驟。
近年來得以發展的葉綠素熒光動力學技術,可快速、高效、無損傷地檢測植物的實際和潛在光合能力、抗逆境脅迫能力等,從而在光合、生態、品種篩選上發揮著越來越大的作用[9-10]。誘變育種中對目標性狀篩選前,先利用葉綠素熒光動力學技術將正常及逆境條件下光合能力增強的植株篩選出,將便于長勢旺盛的突變株及目標株系的篩選,極大減少目標性狀篩選的工作量。而關于葉綠素熒光動力學技術輔助篩選突變株的報道尚較少。
NaN3具有高效、低毒、價廉、使用安全等優點,是一種常用的化學點突變誘變劑[11-12]。本研究在不同處理濃度、處理時間下以NaN3處理馬齒莧種子,通過綜合比較種子萌發速度、萌發質量、誘變當代植株的形態和長勢、葉綠素熒光動力學參數等,得到最佳的NaN3處理濃度和處理時間,并初步篩選植株高、長勢旺盛、抗逆能力強的潛在優良植株,為葉綠素熒光動力學技術在誘變育種中的進一步應用提供參考。
1材料與方法
馬齒莧種子于2012年9月下旬采自南京師范大學生命科學學院植物園。試驗分為2個部分,即分析NaN3處理對馬齒莧種子萌發的影響;比較經NaN3處理后各組幼苗生長、光合性能的變化。
1.1種子萌發
選擇飽滿、整齊的種子并均分為10組,每組設3個重復。各組分別用0.0、0.5、1.0、1.5、2.0 mmol/L NaN3溶液(pH值為4)浸種8、12 h,并用清水沖洗30 min,其中8 h處理組須再用清水浸種4 h以達到12 h的最佳浸種時間[11]。參照楊子儀等的培養皿濾紙法[13],于CONVIRON A-1000型人工智能生物物候培養箱[14 h/10 h(L/D),32 ℃/28 ℃,照度300 μmol/(m2·s),相對濕度(RH) 65%]中萌發種子并記錄萌發種子數,萌發結束時每組隨機選取20棵幼苗測定其胚根與下胚軸的長度,并計算發芽率、發芽指數、活力指數。
1.2幼苗培養
將萌發的芽苗移入Jiffy泥炭育苗塊中,置于CONVIRON A-1000型人工智能生物物候培養箱[14 h/10 h(L/D),32 ℃/28 ℃,照度300 μmol/(m2·s),RH 65%]中培養。10 d后篩選長勢優良的幼苗20株并繼續培養,待幼苗長至約15 cm時對各植株進行葉綠素熒光相關指標的測定;45 d時測定各植株的生長相關指標,并在生長后期種子成熟時收集種子。
1.3葉綠素熒光參數的測定
利用FC-1000-H型便攜式葉綠素熒光成像系統進行葉綠素熒光檢測。該系統提供的測量光為620 nm紅光,強度為500 μmol/(m2·s),照度為500 μmol/(m2·s),持續時間為2 min;飽和脈沖強度為1 800 μmol/(m2·s),持續時間為0.8 s。
首先選取馬齒莧植株上的第5張完全展開葉,暗適應 20 min 后測量最小熒光Fo和暗適應下的最大熒光Fm;隨后打開光化學誘導熒光動力學程序,并間隔30 s打開飽和脈沖測量光適應下的最大熒光Fm′;光化學光持續10 min后將其關閉,并在關閉后第60、100、1 000 s分別打開飽和脈沖測量最大熒光Fm″。
熒光參數的定義采用van Kootan等的方法[14],并由儀器自帶軟件完成計算。光化學光關閉后100、1 000 s時的非光化學淬滅,與光化學光打開時非光化學淬滅的差值分別為qE、qT,1 000 s 時的非光化學淬滅為qI[15]。
1.4生長指標的檢測
馬齒莧移栽45 d后用直尺測量其株高,并采用ADC-AM300 型多用途葉面積儀測量第5張完全展開葉的面積。
1.5數據處理
采用SPSS 19軟件分析試驗數據,采用GraphPad Prism 6軟件制作圖表。試驗結果以“平均值±標準差”表示,表中不同小寫字母表示在單因素方差分析時不同處理間差異顯著(P<0.05)。
2結果與分析
2.1NaN3處理對種子萌發的影響
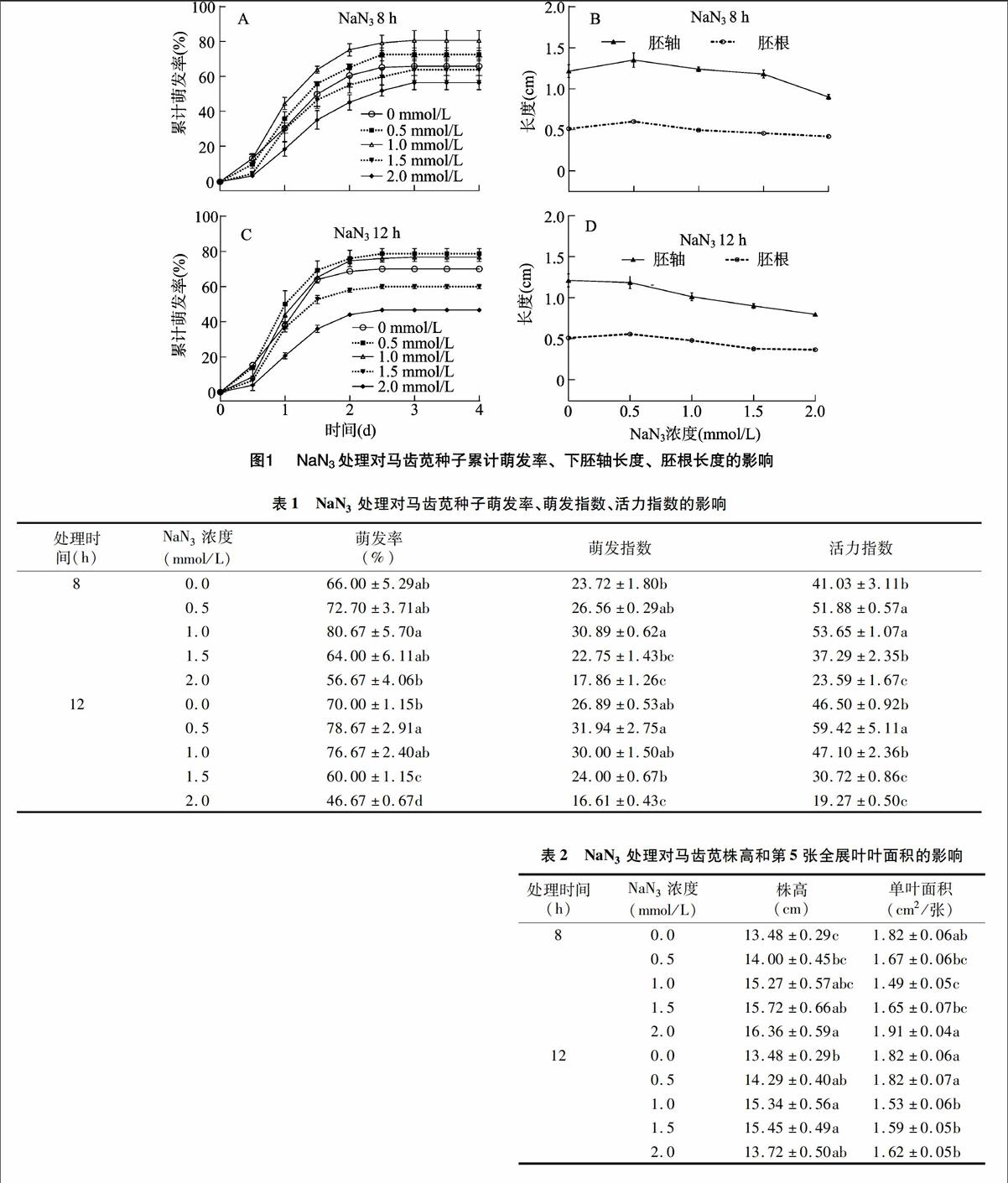
以不同濃度NaN3溶液浸種,分別處理馬齒莧種子8、12 h,再在正常條件下萌發。由圖1-A、圖1-C可知,NaN3處理8、12 h對種子初始萌動時間的影響不大,但隨萌發時間的延長,濃度≤1.0 mmol/L的NaN3處理可顯著提高種子的累計萌發率;1.0 mmol/L NaN3 8 h處理組的累計萌發率最高,高達80.67%±5.7%;0.5、1.0 mmol/L NaN3 12 h處理組對種子萌發的促進效果相當;2.0 mmol/L NaN3處理8 h與1.5~2.0 mmol/L NaN3處理12 h則顯著降低了種子的累計萌發率和最終萌發率(表1)。
NaN3處理不僅影響種子的萌發速度和萌發率,對芽苗質量也有一定影響(表1)。0.5 mmol/L NaN3處理8 h時,芽苗下胚軸長度、胚根長度均顯著高于對照,但隨著NaN3處理濃度的增加,當處理時間延長至12 h,芽苗的胚根長度、胚軸長度逐漸下降(圖1-B、圖1-D)。雖然馬齒莧種子的萌發指數、活力指數對NaN3處理更為敏感,但低濃度、短時間的NaN3處理能提高馬齒莧種子的萌發速度和質量,反之則表現為抑制。
2.2NaN3處理對馬齒莧生長的影響
為檢測NaN3處理對馬齒莧幼苗的生長是否有影響,經NaN3處理的種子萌發后將其移栽到育苗塊于正常條件下培養,并間苗留下優勢株,45 d后比較馬齒莧植株株高、第5張全展葉的葉面積(表2)。除0.5、2.0 mmol/L 12 h處理組外,各組的馬齒莧株高均顯著高于對照,但各組間差距較小,其中 2.0 mmol/L NaN3 8 h處理組的平均株高最高;0.5~1.5 mmol/L NaN3 8 h各處理組以及1.0~2.0 mmol/L NaN3 12 h各處理組的單葉面積均小于或接近于對照,其中2.0 mmol/L NaN3 8 h處理組顯著高于其他組。可見,NaN3處理馬齒莧種子可顯著提高馬齒莧株高,但多數情況下會減小單葉面積。在本試驗的處理濃度及處理時間范圍內,2.0 mmol/L NaN3處理8 h時,馬齒莧植株的單葉面積穩中有升,且株高增幅最大(較對照提高21.36%),表明 2.0 mmol/L NaN3處理馬齒莧種子8 h可顯著提高植株長勢。
2.3NaN3處理對馬齒莧植株光合性能的影響
在葉綠素熒光參數指標中,Fv/Fm值代表PSⅡ的最大光化學效率,絕大多數正常生長植物的Fv/Fm值恒定在0.80~0.85范圍內,而Fv/Fo值則代表實際檢測條件下PSⅡ的光化學活性[16]。由表3可知,對照組馬齒莧葉片的Fv/Fm值高達
表2NaN3處理對馬齒莧株高和第5張全展葉葉面積的影響
0.850,表明馬齒莧葉片的潛在光合性能很高,而各NaN3處理組植株葉片的Fv/Fm值均顯著高于對照,其中2.0 mmol/L NaN3 8 h處理組的Fv/Fm值高達0.865;各NaN3處理組的Fv/Fo值均顯著高于對照,同樣以2.0 mmol/L NaN3 8 h 處理組最高,高達6.38。可見,以NaN3處理馬齒莧種子不僅能提高植株葉片的潛在光化學性能,還可顯著提高植株的實際光化學活性,從而提高植株的實際光合能力。就馬齒莧而言,以表3NaN3處理對馬齒莧葉片葉綠素熒光參數的影響
2.0 mmol/L NaN3處理種子8 h對于植物潛在、實際光合性能的提高最有利。
植物葉綠素吸收的光能不僅用于光化學反應,還有部分能量用于非光化學耗散(NPQ)。非光化學耗散所消耗的能量主要包括qE、qT、qI,分別表示植物在高光強不同階段的光保護能力[14,17-18]。
qE通過提高內囊體內側的pH值,并誘導激發態葉綠素分子中過量的光能以熱的形式耗散,從而對激發態葉綠素淬滅,減少強光誘變下自由基的產生,以減少細胞損傷[18-19]。qE使葉片在幾秒至幾分鐘內快速對強光誘變作出調節,減輕植物受到的光誘變傷害。qE/NPQ的值越大,表明植物對強光環境的適應能力越強。在飽和脈沖刺激下,0.5、2.0 mmol/L NaN3 8 h處理組的qE/NPQ值均與對照相當,2.0 mmol/L NaN3 8 h處理組顯著高于0.5 mmol/L NaN38 h處理組,而其余各組的qE/NPQ值均顯著低于對照。可見在正常生長情況下,NaN3處理馬齒莧種子可促進植株潛在光合性能、實際光化學效率的提高,但在強光脅迫下,多數處理組植株對光脅迫的快速保護能力下降,而2.0 mmol/L NaN3 8 h處理組植株對光脅迫的快速保護能力穩中有升。
qT在三者中所占比例最小,代表激發能由PSⅡ向PSⅠ分配的情況。強光刺激下qT升高表明在幾分鐘內促進能量由PSⅡ向PSⅠ分配,從而避免PSⅡ的光能吸收過量而引起光損傷[20]。在本試驗的NaN3處理濃度、處理時間范圍內,只有0.5、2.0 mmol/L NaN3 8 h處理組的qT/NPQ值與對照差異不顯著,其余各處理組均顯著高于對照。可見,雖然各處理組對光脅迫的快速保護能力下降,但仍可通過將激發能由PSⅡ向PSⅠ轉移,從而在幾分鐘內提高植株對光脅迫的保護能力,此保護能力相對較小。0.5、2.0 mmol/L NaN3 8 h處理組的植株能量由PSⅡ向PSⅠ的分配比例與對照相當。
在強光下,反應中心D1蛋白受損傷的速度超過其修復速度,qI代表捕光色素蛋白復合體吸收的光能未被有效利用而引起的能量耗散,這部分能量主要轉化為熱能或形成活性氧[18]。雖然qI是光損傷和光保護混合作用的結果,但qI/NPQ的值越大往往代表植物所受的光抑制越強[18,21]。由表3可知,2.0 mmol/L NaN3 8 h處理組的 qI/NPQ 值與對照相當,而其余各組均顯著高于對照,且各12 h處理組的 qI/NPQ 值顯著高于各8 h處理組。可見,除2.0 mmol/L NaN3 8 h處理組外,其余各處理組植株在強光下的損傷均顯著高于對照組,且植物在強光下的保護能力隨NaN3處理時間的延長而減弱。
3結論與討論
NaN3作為一種高效的誘變劑,被廣泛應用于水稻、小麥、西瓜的種子處理以及組織培養中的外植體處理,以促進誘變的發生。NaN3誘變時,突變多為隱性突變,且具有無定向性等特點,導致誘變處理后多數植株長勢減弱,目標性狀在子一代或子二代后才能顯現。若以種子萌發勢、當代或子一代植株葉片畸形率等來確定最適劑量,則可能導致大量后代植株長勢偏弱,這與目標性狀的篩選相違背;因此,本研究綜合種子萌發、植株長勢、光合能力來確定NaN3的最適處理劑量,并采用葉綠素熒光動力學技術輔助篩選抗性較強的植株,以加速篩選進程。
絕大多數研究結果表明,NaN3處理可延緩種子萌發并降低種子萌發率[6,11]。而本研究中,1.0 mmol/L NaN3處理8 h或 0.5 mmol/L NaN3處理12 h均可顯著提高種子的萌發率及萌發質量;更高處理濃度、更長處理時間則會使馬齒莧種子的萌發率、萌發質量逐漸下降。馬齒莧種子對NaN3處理的抗性大于其他種子,因此不應采用常規半致死率來確定馬齒莧種子的最適NaN3誘變劑量。
將萌發種子在育苗塊上培養,間苗篩選留下長勢旺盛的植株,并比較各組植物的生長及光合性能。結果顯示,初篩后各處理組植株的株高均高于對照或與對照相當,且正常生長條件下植株的潛在(Fv/Fm值)、實際光化學活性(Fv/Fo值)均強于對照或與對照相當[16],但各處理組增大幅度不一。雖然活力指數對NaN3處理最敏感,但株型最高、光合性能最強的2.0 mmol/L NaN3 8 h處理組在種子萌發期的活力指數為 57.49%。可見,為使后代植株中出現更多長勢旺盛、光合能力強的植株,應綜合萌發質量、長勢、光合能力等指標來確定NaN3的最適處理劑量。
正常條件下生長旺盛的植株不一定具有高抗性,而農業生產中生長旺盛、抗性強的植株更具有應用價值。若在當代先篩選長勢旺盛、光合能力強的植株,并在子一代篩選抗性強的植株,則會遺漏當代植株中長勢不強但抗性較強的植株,同時增加子一代植株的篩選工作量。若能篩選當代植株中長勢旺盛且抗性強的植株,對減少遺漏、加快篩選進程、減少工作量等極為有利。
葉綠素熒光動力學技術可通過改變外源光強,無損傷檢測植株在正常光強下的光合性能,及其在強光下非光化學淬滅各組分比例(qE/NPQ、qT/NPQ、qI/NPQ)的變化,并分析植株在正常條件下的光合潛能和高光強抵御能力[21]。該方法已在優良品種的抗性篩選上發揮了一定作用。本研究以此方法分析正常生長條件下馬齒莧植株的抗逆能力,并綜合植株的其他性狀選擇NaN3最適處理劑量和目標性狀的備選株系。結果表明,NaN3處理可提高植株在正常條件下的光合性能,但也會導致多數植物對高光強逆境的抵御能力減弱。NaN3處理降低了除2.0 mmol/L NaN3 8 h處理組外其余各組植物對光脅迫的快速保護能力(qE/NPQ),并促進其將PSⅡ吸收的多余能量向PSⅠ轉移的能力(qT/NPQ),而由于此能力有限,最終導致這些植株在強光下的損傷高于對照 (qI/NPQ)。2.0 mmol/L NaN3 8 h處理組的植株不僅PSⅡ的潛在(Fv/Fm)、實際光化學性能(Fv/Fo)提升幅度最大,且該組植物在強光脅迫下可將非光化學淬滅中的能量更多地應用于快速光保護(qE/NPQ),而用于熱耗散和產生自由基的能量比例相應減少(qT/NPQ),最終導致植物對高光強的抵御能力增強。試驗證實了正常條件下長勢旺盛的植株,其抗逆性不一定強;利用葉綠素熒光動力學技術可無損傷篩選同時具有光合旺盛、抗逆性強特性的植株。
綜合不同濃度NaN3處理下的種子萌發質量、初篩后植株的長勢、植株光合能力、抗高光強能力可知,以2.0 mmol/L NaN3處理8 h是馬齒莧種子的最適處理條件;采用葉綠素熒光動力學分析技術分析植株的潛在和實際光合性能、抗高光強能力,極大地提高篩選效率。后續研究將對收集的種子在子一代中進一步篩選,以篩選出生長旺盛、抗性強、抗白粉病、營養豐富的馬齒莧種質。
參考文獻:
[1]Simopoulos A P,Tan D X,Manchester L C,et al. Purslane:a plant source of omega-3 fatty acids and melatonin[J]. Journal of Pineal Research,2005,39(3):331-332.
[2]Feng P C,Haynes L J,Magnus K E. High concentration of (—)-noradrenaline in Portulaca oleracea L.[J]. Nature,1961,191:1108.
[3]Sauer L A,Dauchy R T,Blask D E. Polyunsaturated fatty acids,melatonin,and cancer prevention[J]. Biochemical Pharmacology,2001,61(12):1455-1462.
[4]Tiwari K K,Dwivedi S,Mishra S,et al. Phytoremediation efficiency of Portulaca tuberosa rox and Portulaca oleracea L. naturally growing in an industrial effluent irrigated area in Vadodra,Gujrat,India[J]. Environmental Monitoring and Assessment,2008,147(1/2/3):15-22.
[5]安學麗,蔡一林,王久光,等. 化學誘變及其在農作物育種上應用[J]. 核農學報,2003,17(3):239-242.
[6]杜連恩,魏玉昌,可福存,等. 大豆化學誘變育種及其規律的研究[J]. 華北農學報,1989,4(2):39-43.
[7]董穎蘋,連 勇,何慶才,等. 植物化學誘變技術在育種中的運用及進展 Ⅱ.突變體的篩選及分子檢測[J]. 種子,2005,24(8):54-58.
[8]陳庭,王愛敏,劉運權,等. NaN3對寶巾的誘變效應初步研究[J]. 中國農學通報,2012,28(25):191-195.
[9]Codrea M C,Hakala Y M,Krlund M A,et al. Mahalanobis distance screening of Arabidopsis mutants with chlorophyll fluorescence[J]. Photosynthesis Research,2010,105(3):273-283.
[10]Ptushenko V V,Ptushenko E A,Samoilova O P,et al. Chlorophyll fluorescence in the leaves of Tradescantia species of different ecological groups:induction events at different intensities of actinic light[J]. Bio Systems,2013,114(2):85-97.
[11]陳燦,徐慶國,彭波,等. 不同化學誘變劑對水稻種子萌發和生長的影響[J]. 種子,2008,27(3):9-13.
[12]楊子儀. 野生型與栽培型馬齒莧萌發特性與抗逆能力比較[D]. 南京:南京師范大學,2014.
[13]楊子儀,徐亞莉,葛峰,等. 獸藥洛克沙胂處理對空心菜生長代謝和營養價值的影響[J]. 地球與環境,2013,41(4):441-450.
[14]van Kooten O,Snel J F H. The use of chlorophyll fluorescence nomenclature in plant stress physiology[J]. Photosynthesis Research,1990,25(3):147-150.
[15]Walters R G,Horton P. Resolution of components of non-photochemical chlorophyll fluorescence quenching in barley leaves[J]. Photosynthesis Research,1991,27(2):121-133.
[16]Schreiber U,Bilger W,Hormann H,et al. Chlorophyll fluorescence as a diagnostic tool:basics and some aspects of practical relevance[M]. Cambridge:Cambridge University Press,1998:320-336.
[17]Hodges M,Comic G,Briantais J M. Chlorophyll fluorescence from spinach leaves:resolution of non-photochemical quenching[J]. Biochimica et Biophysica Acta(BBA)-Bioenergetics,1989,974(3):289-293.
[18]Müller P,Li X P,Niyogi K K. Non-photochemical quenching:a response to excess light energy[J]. Plant Physiology,2001,125(4):1558-1566.
[19]Demmig A B,Adams Iii W W. The role of xanthophyll cycle carotenoids in the protection of photosynthesis[J]. Trends in Plant Science,1996,1(1):21-26.
[20]Fork D C,Bose S,Herbert S K. Radiationless transitions as a protection mechanism against photoinhibition in higher plants and a red alga[J]. Photosynthesis Research,1986,10(3):327-335.
[21]Rohácˇek K,Soukupová J,Barták M. Chlorophyll fluorescence:a wonderful tool to study plant physiology and plant stress[M]//Schoefs B.Plant cell compartments-selected topics. Kerala,India:Research Signpost,2008:41-104.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
