一種新的鐮刀菌Q7—31木聚糖酶Xyn9的分離純化鑒定及酶學特性
2015-10-20 21:10:49趙聯正謝占玲趙朋
江蘇農業科學 2015年5期
趙聯正 謝占玲 趙朋
摘要:為對植物病原菌鐮刀菌(Fusarium sp.)Q7-31所產木聚糖酶進行鑒定及酶學特性分析,在發酵產酶培養基上對Q7—31進行發酵培養,獲得能夠高效降解植物細胞壁的粗酶液。然后,采用Sephacry S-100凝膠柱層析和DEAE弱陰離子交換柱層析對粗酶液進行分離純化后得到木聚糖酶Xyn9,并對其開展蛋白質組學研究和酶學特性分析。結果表明:粗酶液中木聚糖酶比活為16.58U/mg,纖維素酶比活為0.428U/mg,細胞壁降解酶比活為0.0198U/mg,蛋白含量為2.17mg/mL。Xyn9的蛋白質組學研究結果顯示,其分子量為21 ku、等電點為6.86,接近GHl0家族。純化后Xyn9的酶學特性表明,最適反應溫度為47℃,最適pH值為5.6,該酶在33℃以下和在pH值5~6的范圍內較穩定;金屬離子K+對該酶有激活作用,Na+、Ca2+、Mg2+、Zn2+抑制該酶活性,Cu2+、Hg+使該酶完全失活。綜合Xyn9的分子量、蛋白質組學結果以及酶學特性,最終將其鑒定為一種新的介于GHl0家族與GHll家族之間的內切木聚糖酶。
關鍵詞:鐮刀菌;木聚糖酶;分離純化;串聯質譜鑒定;酶學特性
中圖分類號:Q55;Q814.1 文獻標志碼:A 文章編號:1002-1302(2015)05-0042-04
植物細胞壁主要由纖維素、半纖維素和木質素等物質組成。半纖維素的主要成分是木聚糖,約占植物細胞干質量的30%-35%,是繼纖維素之后含量第二豐富的可再生物質資源。其中,木聚糖酶(1,4-β-D—xylanase,EC 3.2.1.8)是一種重要的工業用酶,在造紙工業、能源、飼料及環境等領域中具有很好的應用前景,特別是在紙漿的生物漂白方面的應用。生物降解木質素,就是利用微生物產生的胞外酶使木質素逐步分解轉化,從而實現其資源轉化。所以,木質素酶作為廢棄秸稈回收利用過程中的關鍵角色,已引起人們的高度重視。我國是農業生產大國,每年秸稈產量大約5.7億t,占世界秸稈總產量的20%~30%。其中絕大部分因為難以利用,僅作為農家燃料或肥料。如果能夠將這些秸稈通過轉化成為低聚糖,就可以用作基本碳源,生產各種發酵產品,包括各種氨基酸、有機酸、單細胞蛋白、糖醇類、工業酶類、溶劑或燃劑等。因此,對木聚糖酶的研究具有十分可觀的經濟效益和社會效益。
根據疏水性聚類分析方法,將木聚糖酶分為GHl0和GHl 1等2個家族。GHl0家族木聚糖酶多為高分子量,通常大于30ku,等電點較低,不僅有催化結構域,還常有碳水化合物結合結構域;GHl1家族木聚糖酶多為單結構域蛋白,相對分子量一般小于30ku,等電點較高。而且2個家族的木聚糖酶在空間結構上的折疊類型不同,即GHl0家族木聚糖酶主要由重復出現的α-螺旋和β-折疊片構成,整體結構像碗狀,上面略大下面較小;而GHll家族木聚糖酶具有的典型折疊結構是由β-折疊片為主所構成的單個結構域,這個結構域由2個β-折疊片層扭曲成將近90°角,構成1個深且狹長的溝縫狀結構。
近10年來,基因重組、基因定位突變、細胞融合等技術在纖維素酶菌株的構建上取得了巨大的成果。對于木聚糖酶而言,酶活力偏低是研究開發中遇到的普遍問題,低酶含量造成發酵成本過高,限制了該酶制劑的工業化生產和應用。一般來說,真菌木聚糖酶比細菌的木聚糖酶產量大,例如木霉、曲霉等真菌所產木聚糖酶的酶活力較高,對木聚糖酶的研究也主要集中在這些方面。鐮刀菌是一種世界性的土生絲狀真菌,其中有一些可以產生多種酶,其能夠降解細胞壁中的果膠與纖維素。由于鐮刀菌屬于病原菌,對鐮刀菌的研究多集中在對致病性鐮刀菌的檢測鑒定、多樣性與致病基因等方面,對利用其產酶的研究鮮有報道。本研究對鐮刀菌(Fusarium sp.)Q7-31產細胞壁降解酶進行分離純化,并對純化后的木聚糖酶進行蛋白鑒定與酶學特性分析,鑒定出介于GHl0家族與GHll家族的內切木聚糖酶Xyn9,為后續研究細胞壁降解酶降解細胞壁的協同反應機制、反應條件及酶系的結構組成奠定基礎。
1.材料與方法
1.1材料
1.1.1菌株鐮刀菌Q7-31菌株,由青海大學生態環境工程學院微生物實驗室于2007年從青海省大通縣寶庫鄉所采土壤中自行分離純化獲得,并保存。
1.1.2培養基
固體活化培養基:PDA培養基粉末39g,1 000mL水,121 qC滅菌20min;液體種子培養基:葡萄糖2%,蛋白胨0.3%,Mendels營養鹽,121℃高壓滅菌20min;發酵產酶培養基:燕麥秸稈粉0.3%,蛋白胨0.3%,Mendels營養鹽,121℃高壓滅菌20min;Mendels營養鹽:(NH4):SO4 1.4g/L、KH2PQ 2.0g/L、尿素0.3g/L、CaCl20.3g/L、MgSO4 0.3g/L、FeSO4 5.0 mg/L、MnSO4 1.6mg/L、ZnSO4 1.4mg/L、COCl2 2.0mg/L。
1.2方法
1.2.1菌種活化與發酵將保存在4℃下液體石蠟封存的斜面菌種Q7-31轉接至固體活化培養基上,于25℃恒溫培養箱中培養,傳代2~3次,使菌種充分活化。液體種子制備:在1000mL三角瓶中裝入200mL液體種子培養基,從固體活化培養基上接種直徑為1.0cm的菌塊至液體種子培養基中,于25℃恒溫培養箱中靜止培養2d后制成液體種子備用。液體發酵培養:在1000mL三角瓶中裝入350mL發酵產酶培養基,將制備好的液體種子按10%接入發酵產酶培養基中,25℃ 120r/min條件下搖床培養。1.2.2酶活力的測定采用3',5'二硝基水楊酸(DNS)測定還原糖的方法,測定分別以1%木聚糖、0.5%羧甲基纖維素(cMC)及5mg/mL燕麥秸稈粉為底物時酶液中木聚糖酶、纖維素酶和植物細胞壁降解酶的活力。將1min產生1ug還原糖所需的酶液量定義為1個酶活力單位(u)。
1.2.3蛋白含量的測定
蛋白含量的測定采用Bradford法,以牛血清白蛋白(BsA)為標準繪制標準曲線。在2mL離心管中加入稀釋3倍的酶液樣品200uL,加入1mL考馬斯亮藍G-250染液,充分振蕩混勻反應5min,60min內在595nm下測定吸光度。將測得的吸光度代入蛋白標準曲線方程,計算樣品中的蛋白含量。
1.2.4酶的分離純化用50%~80%飽和度的硫酸銨對蛋白含量較高的酶液進行分級沉淀,后將6mL沉淀后的樣品上樣到Sephacry s-100葡聚糖凝膠層析柱(26mm×700mm),層析柱預先用pH值為7.5的Tris-HCl緩沖液平衡,并以相同緩沖液進行洗脫,速度為1.3mL/min,用核酸蛋白檢測儀在280nm波長下檢測蛋白分布,對每個收集管進行標號;將初步分離的蛋白樣品上樣到DEAE弱陰離子交換柱(5×5mL),柱子預先用20mmol/L Tris—HCl緩沖液(pH值為7.5)平衡,以O~1mol/L的NaCl進行梯度洗脫,速度為0.5mL/min,每4min收集1管,用核酸蛋白檢測儀在280nm波長下檢測蛋白分布。
1.2.5SDS一聚丙烯酰胺凝膠電泳(SDS-PAGE)將制好的分離膠和濃縮膠灌入電泳槽后,把蛋白Loading Buffer與待測收集管中的樣品混合,于100℃沸水浴中加熱5min,取出后冷卻至室溫。每個點樣孔上樣10uL,并使用標準蛋白分子量作為參照。當溴酚藍前沿到達距凝膠下沿約0.5cm時,停止電泳。使用小刀將凝膠輕輕撬離玻璃板,然后將凝膠剝離到裝有染色液(考馬斯亮藍R-250染色液1%,甲醇40%,冰醋酸10%,混勻并過濾)的培養皿中,染色3~4 h。染色完畢后,倒出染色液,加入脫色液(甲醇10%,冰醋酸10%,混勻),脫色3~10h,定時更換脫色液,直至凝膠背景變得清晰可見。
1.2.6酶的質譜鑒定酶的質譜鑒定由華大基因(北京)完成。切下的目的蛋白凝皎位點,加入50mmol/L NH4HCO,振蕩至無色;加入乙睛100uL約放置30min',通過1ug/mL胰蛋白酶酶解37℃水浴16h,加入100uL提取液(50%乙睛,5%TFA),40℃水浴1h,真空干燥除去乙睛,加入5uLO.1%三氟乙酸。取2ug的樣品用Angilent 1100液相系統Finnigan線性離子阱(LTQ)質譜儀系統進行檢測,質譜結果經MASCOT軟件檢索,結合NCBI數據庫信息以及酶學特性確定酶的種類。
1.2.7酶的最適溫度、pH值及穩定性測定最適溫度的測定:將底物處于pH值6.8的緩沖液中,在4~90℃范圍中進行酶促反應,確定酶的最適反應溫度;在上述不同溫度下溫浴處理1h測定酶的穩定性。最適pH值的測定:將底物處于不同pH值的緩沖液中,pH值范圍為3~8,在45℃條件下反應并測定酶活力,確定最適反應pH值。穩定性測定:將樣品分別在pH值為3.0~11.0的條件下處理1h,測定酶的pH值穩定性。
1.2.8金屬離子對酶活力的影響分別在酶液中加入金屬離子化合物Na+、K+、zn+、Cu2+、Fe2+、Mg2+、Mn2+、Ca2+、Hg2+,各種金屬離子在反應體系中的終濃度分別達到4、8mmol/L。40℃處理30min后測定殘余木聚糖酶活力,未加金屬離子的酶活力為100%。
2.結果與分析
2.1Q7-31發酵液的酶活力和蛋白含量
2.1.1酶活力Q7-31發酵液的酶的木聚糖酶活力最強,在酶液稀釋100倍情況下其木聚糖酶活力高達35.96U/mL;其次具有一定的纖維素酶活力,在酶液稀釋10倍的情況下,纖維素酶活力為0.929U/mL;而酶液在沒有稀釋的情況下,細胞壁降解酶活力只有0.043U/mL(表1)。以上結果表明,Q7-31發酵液的酶主要具有木聚糖酶活力。
2.1.2蛋白含量過濾后的酶液總體積為1 400 mL,595 nnl下的吸光度為0.840,代入標準曲線公式(y=0.8609x,r2=0.9994),得到酶液總蛋白含量為3037.26mg。
2.2酶的分離純化及SDS-PAGE
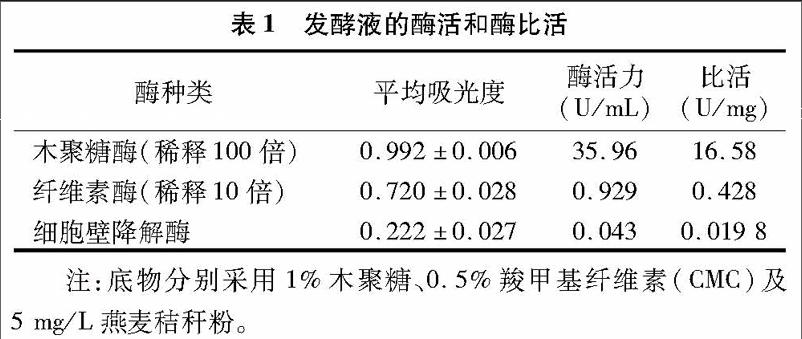
1400mL粗酶液經過50%~80%硫酸銨分級沉淀后,得到12 mL酶液,蛋白含量為1248.99mg,蛋白回收率為41.12%。將酶液上樣到Sephacry s-100葡聚糖凝膠層析柱中進行初步分離純化,得到只具有木聚糖酶活性的蛋白,將其命名為Xyn9,其酶活力為47.26U/mL,蛋白含量為5.824mg,比活為24.34U/mg,SDS-PAGE檢測結果證明其為單一蛋白,分子量為21ku(圖1)。再將初步分離純化的Xyn9上樣到EzFast DEAE弱陰離子交換柱中進一步分離純化,柱子預先用pH值為8.5的Tris-HCl緩沖液平衡,以0~1mol/L NaCl進行梯度洗脫,速度為0.5mL/min,得到純化過的Xyn9。
2.3Xyn9的質譜鑒定
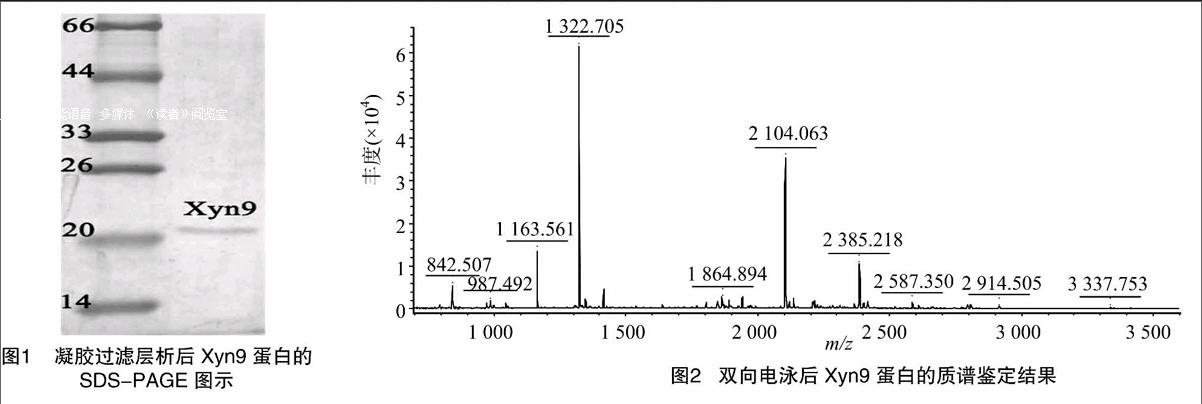
選取雙向電泳為1個點的Xyn9蛋白進行串聯質譜鑒定,
2.4xyn9的酶學特性
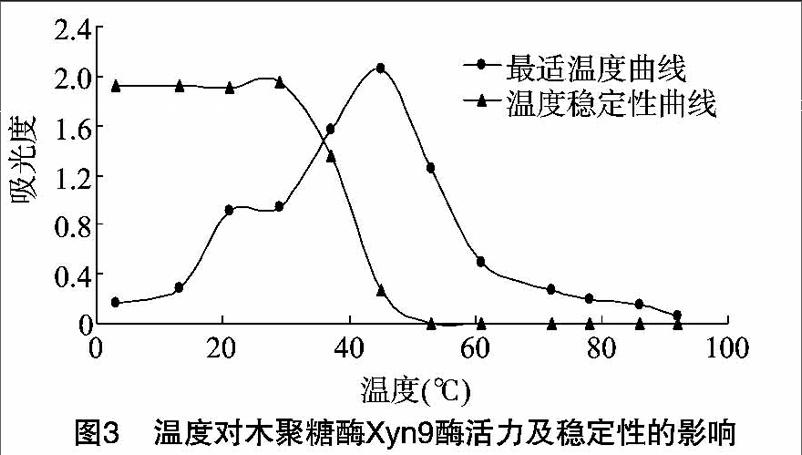
2.4.1Xyn9的最適溫度及溫度穩定性Xyn9的最適反應溫度為47℃,在37~52℃范圍內都有較高的酶活力,溫度過低或過高都不利于酶促反應;Xyn9在33℃以下穩定性高,當溫度高于33℃酶活力迅速降低直至為O(圖3)。
2.4.2Xyn9的最適pH值及pH值穩定性純化的低分子量木聚糖酶Xyn9的最適反應pH值為5.6;Xyn9在pH值為5~6的范圍內具很好的穩定性(圖4)。
2.4.3金屬離子對Xyn9酶活力的影響
金屬離子對木聚糖酶Xyn9酶活力具有明顯的影響。K+在4、8 mmol/L離子強度條件下對該木聚糖酶活力都有一定的激活作用,且離子濃度越大,激活效果越好;Na+、Ca2+、Mg2+、Zn2+對該酶有不同程度的抑制作用;Hg2+、cu2+則完全抑制該木聚糖酶的活力(圖5)。
2.5Xyn9的鑒定
由于Xyn9的質譜鑒定結果表明其接近但不屬于GH10結果表明:由于Mascot Score為37分,不足證明其為GHl0家族內切木聚糖酶的47分(P<0.05),因此Xyn9接近但并不屬于GHl0家族內切木聚糖酶(NCBI注冊號:gi1170139)。理論等電點(PI)為6.86,質譜鑒定結果如圖2所示,匹配的肽段序列為VLGEDFVGIAFR,覆蓋范圍為3%。家族內切木聚糖酶,而低分子量以及酶學特性表明其接近GHll家族內切木聚糖酶。因此,最終將其鑒定為一種新的介于GHl0與GHll家族之間的內切木聚糖酶。
3.結論與討論
對鐮刀菌Q7-31發酵液進行酶活力測定,結果表明,發酵液具有木聚糖酶、纖維素酶與植物細胞壁降解酶活力,并且植物細胞壁降解酶活力較強,這種多種酶的組合在真菌中并不常見。這可能是其生活方式與環境相適應而長期進化的結果,作為植物病原菌,多種酶的協同作用可能有助于其高效感染植物,完成其生活過程。此外,病原菌木聚糖酶可能作為激發子,誘發植物的抗病防御反應,在植物與病原菌互作機制研究和植物抗病防病領域中備受關注,病原菌木聚糖酶由于能分解植物細胞壁而可能促進病原菌的侵染,且病原菌木聚糖酶還可能同時誘發植物的抗病防御系統,病原菌木聚糖酶的這種雙重作用已成為近年來植物一病原菌互作關系研究中的一個熱點,因此侵染過程需要一系列酶的協同作用。植物半纖維素的完全水解需要一系列酶協同完成,并且這些酶共同作用致使粗酶液具有較高的比活力。其中的小分子木聚糖酶更容易穿透植物纖維壁結構,發揮高效的降解作用。而一些木聚糖酶具有一些特殊結構區域,有利于和底物結合,熱穩定區域和糖基化結構使其穩定性在不同環境中更好。
筆者在之前的研究中,已經克隆出編碼Xyn8蛋白(同樣從Q7-31中分離出)的Xyn8基因,并且實現了其在大腸桿菌中的原核表達,發現其為25.7ku的GHll家族內切木聚糖酶,其最適溫度、pH值分別為40℃、6.0。將本研究所得到的Xyn9與Xyn8對比后發現,Xyn9的分子量以及酶學特性與Xyn8類似,即接近GHll家族內切木聚糖酶,但兩者基因序列的對比結果(57%)證明其不屬于GHll家族。此外,雙向電泳后的串聯質譜鑒定結果表明其接近但不屬于GHl0家族內切木聚糖酶。因此,根據Xyn9的分子量、質譜鑒定結果以及其酶學特性,將其最終鑒定為一種新的介于GHl0與GHl 1家族之間的內切木聚糖酶。但由于其作為病原菌所產酶的特殊性,后續著重解決其歸屬問題。
Xyn9的酶學特性研究表明其最適反應pH值為5.6,低于33~C時酶活力較穩定,最適反應溫度為47℃。多數真菌木聚糖酶的最適反應pH值在5左右,pH值穩定范圍為5~6,最適反應溫度為40~70℃,低于50℃較穩定。但也有一些報道認為最適pH值在更酸或偏中性環境中,如lefuji等從酵母菌Cryptococcus sp.S-2中分離得到一種小分子量內切木聚糖酶,其最適反應pH值為2.0,且當pH值為1.0時仍能保持75%的酶活力。另外一些木聚糖酶在極端pH值和溫度條件下酶活力不但高而且穩定,在工業應用方面有很好的前景。如木聚糖酶應用于造紙業中需要其能夠受耐高溫(55~70℃)和堿性環境,在生物轉化方面甚至需要一些嗜熱嗜堿和嗜熱嗜酸性的木聚糖酶。這就需要通過菌株篩選、誘變、基因工程或者蛋白工程等方法來提高木聚糖酶的水解效率和穩定性,這也是目前木聚糖酶研究的一個主要方向。金屬離子K對木聚糖酶Xyn9具有很強的激活作用,在一定范圍內,其濃度越大作用越強,而Na+、ca2+、Mg2+、zn2+對該酶有不同程度的抑制作用很少有報道。重金屬離子Hg2+、cu2+對木聚糖酶具有完全的抑制作用,這與許多報道結果一致,可能由于Hg2+、Cu2+能夠改變酶的構象從而使酶失活。
