紅掌小孢子母細(xì)胞減數(shù)分裂觀察及2n花粉遺傳的細(xì)胞學(xué)分析
2015-10-21 16:47:39徐洪偉冷青云覃海燕賴杭桂尹俊梅
熱帶作物學(xué)報(bào) 2015年11期
徐洪偉 冷青云 覃海燕 賴杭桂 尹俊梅

摘 要 與無性多倍化相比,2n花粉在三倍體育種方面有明顯優(yōu)勢。為了在紅掌育種中使用2n花粉,本研究對(duì)紅掌肉穗花序發(fā)育過程的小孢子母細(xì)胞減數(shù)分裂進(jìn)行觀察。以前期I為誘導(dǎo)時(shí)機(jī),使用秋水仙素溶液棉浸法對(duì)紅掌肉穗花序進(jìn)行誘導(dǎo),觀察到二分體和三分體,獲得了加倍的2n花粉。在小孢子母細(xì)胞第二次減數(shù)分裂過程中,觀察到融合紡錘體,三極紡錘體和加倍的4n小孢子母細(xì)胞與2n花粉形成有直接關(guān)系。
關(guān)鍵詞 紅掌;減數(shù)分裂;2n花粉;秋水仙素誘導(dǎo)
中圖分類號(hào) Q945 文獻(xiàn)標(biāo)識(shí)碼 A
Abstract Compared with mitotic polyploids, 2n pollens are useful for triploid breeding. For further use of 2n pollens in Anthurium andraeanum breeding, we observed the cytological process of meiosis of the pollen mother cell. Moreover, 2n pollen induction was performed with ‘Dakotaanther at the stage of prophase I via treated with 0.3% colchicine+2%dimethyl sulfoxide(DMSO) for 3 days. Our results showed that 2n pollens were induced after treated with colchicines, with triads and dyads. Furthermore, tripolar spindles, fused spindles in male meiosis II and 4n pollen mother cell were produced, which may have related to the formation of 2n pollens.
Key words Anthurium andraeanum;Meiosis;2n pollen gametes;Colchicine induction
doi 10.3969/j.issn.1000-2561.2015.11.021
紅掌(Anthurium andraeanum),又名火鶴花,隸屬于天南星科,原產(chǎn)于哥倫比亞和厄瓜多爾,體細(xì)胞含30條染色體(2n=30)[1-2]。紅掌是僅次于熱帶蘭花的全球第二大熱帶花卉,在全世界有著廣泛的分布和栽培,是研究最深入的花卉之一;紅掌植株能有效吸收空氣中的甲醛、二甲苯、甲苯、氨氣等有毒氣體[3],對(duì)凈化室內(nèi)環(huán)境和美化生活具有積極意義,因而備受青睞。紅掌基因組具有高度的雜合性,其減數(shù)分裂過程往往伴隨著復(fù)雜的染色體行為[2]。減數(shù)分裂是物種有性生殖和遺傳重組的基礎(chǔ),在物種進(jìn)化過程中發(fā)揮重要作用[4]。
基因組加倍(多倍化)在植物進(jìn)化和多樣化中起到非常關(guān)鍵的作用,并且被認(rèn)為是植物物種形成和適應(yīng)中的一個(gè)重要機(jī)制[5]。幾乎所有的植物物種都以某種頻率產(chǎn)生2n配子,并且2n配子對(duì)多倍體源起非常重要[6]。在植物育種中使用2n配子,即有性多倍化,在作物改良方面已經(jīng)被證實(shí)非常有效[7]。2n配子育種不僅可用于創(chuàng)造新的變異,在轉(zhuǎn)移優(yōu)良目標(biāo)基因、穩(wěn)定傳遞雜合性及保持上位性等方面具備獨(dú)特的育種優(yōu)勢[8]。三倍體育種作為多倍體育種的一種主要方式, 具有重大的育種價(jià)值,例如:葉片變大,變厚,花變大,單株的莖變少,株型變緊湊,對(duì)環(huán)境壓力的抵抗力變強(qiáng),對(duì)疾病的抵抗力變強(qiáng)[9]。利用2n花粉直接與正常配子雜交,可以直接獲得三倍體后代,育種周期短。本研究在觀察紅掌小孢子母細(xì)胞減數(shù)分裂進(jìn)程和染色體行為基礎(chǔ)上,使用秋水仙素誘導(dǎo)獲得了紅掌2n花粉,并探討了2n花粉形成的細(xì)胞學(xué)機(jī)制。
1 材料與方法
1.1 材料
供試材料為中國熱帶農(nóng)業(yè)科學(xué)院熱帶作物品種資源研究所提供的紅掌品種:大哥大(Dakota),種植于溫室大棚(自然光照條件,溫度20~30 ℃)。
1.2 方法
1.2.1 倍性分析 倍性分析采用根尖壓片法,卡寶品紅染色。在 Leica DM2500光學(xué)顯微鏡下鏡檢,尋找清晰、分散的分裂中期相拍照。
1.2.2 小孢子母細(xì)胞減數(shù)分裂 小孢子母細(xì)胞減數(shù)分裂觀察采用花藥壓片法。取肉穗花序,分離出花藥,將花藥置卡諾固定液中固定24 h以上;取出花藥,用濾紙吸除固定液,1 mol/L鹽酸60 ℃水浴解離;將花藥轉(zhuǎn)移到載玻片上,滴加卡包寶品紅染色,壓片。在 Leica DM2500 光學(xué)顯微鏡下鏡檢拍照。
1.2.3 誘導(dǎo)2n花粉 以大多數(shù)小孢子母細(xì)胞處于前期I時(shí)的肉穗花序作為2n花粉的誘導(dǎo)時(shí)期。采用棉浸法,肉穗花序被連續(xù)處理3 d,含有2% DMSO的0.3%秋水仙素每天上午7:00、下午7:00滴藥,至脫脂棉全部浸潤。處理完后,對(duì)肉穗花序進(jìn)行清洗,防止殘留藥液毒害肉穗花序。
1.2.4 n花粉和2n花粉直徑的測定 采集新鮮花粉,放置于載玻片,使用醋酸洋紅染色。在 Leica DM2500光學(xué)顯微鏡下觀察,測量花粉直徑,使用SAS軟件分析n花粉和2n花粉直徑在0.01水平上顯著性。
1.2.5 2n花粉形成細(xì)胞學(xué)特點(diǎn) 當(dāng)肉穗花序被處理時(shí),每隔2 h,采集肉穗花序,置卡諾固定液中固定24 h以上,卡寶品紅染色,壓片。在Leica DM2500光學(xué)顯微鏡下,觀察小孢子母細(xì)胞減數(shù)分裂過程中的染色體行為。
2 結(jié)果與分析
2.1 ‘Dakota的倍性分析
根據(jù)根尖染色壓片法,確定‘Dakota紅掌是二倍體(2n=2x=30)(圖1)。
2.2 減數(shù)分裂過程
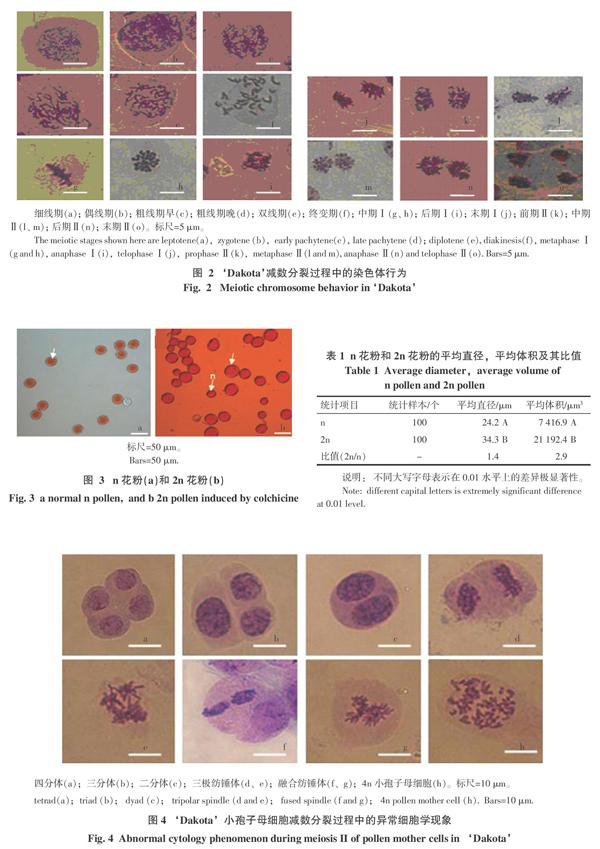
本研究中發(fā)現(xiàn),‘Dakota小孢子母細(xì)胞減數(shù)分裂過程的持續(xù)時(shí)間只有幾天,從肉穗花序的底端開始,自下而上,頂端最后。前期I最復(fù)雜,前期I分為5個(gè)子時(shí)期,分別為:細(xì)線期(圖2-a),偶線期(圖2-b),粗線期(圖2-c~d),雙線期(圖2-e),終變期(圖2-f)。在終變期(圖2)停留很短時(shí)間便進(jìn)入中期I(圖2-g~h)。在同一花藥中,可以看到從偶線期(圖2-b)到末期I(圖2-i~j)的所有分裂相,分裂不同步性高,但都能在1 d內(nèi)完成第一次減數(shù)分裂。第2天,采集相近位置的花藥,很少觀察到第一次減數(shù)分裂的圖像,觀察到的是從前期Ⅱ至末期Ⅱ的圖像(圖2-k~o),第3天,全部停留在四分體階段(圖4-a)。
2.3 n花粉和2n花粉的直徑
采集‘Dakota的正常n花粉(圖3-a),測量n花粉直徑,計(jì)算n花粉體積, 計(jì)算樣本均值(表1)。采集經(jīng)秋水仙素處理后的‘Dakota的花粉(圖3-b),測量其中大花粉的直徑,計(jì)算大花粉體積,計(jì)算樣本均值(表1)。從表1可知:‘Dakota紅掌2n花粉直徑比n花粉大40%左右,SAS軟件分析表明n花粉和2n花粉直徑在0.01水平上達(dá)到極顯著性;2n花粉的體積比n花粉大190%左右,SAS軟件分析表明n花粉和2n花粉體積在0.01水平上達(dá)極顯著性。
2.4 2n花粉遺傳細(xì)胞學(xué)特點(diǎn)
‘Dakota小孢子母細(xì)胞經(jīng)減數(shù)分裂后,形成4個(gè)單倍體孢子,稱作四分體(圖4-a)。在被秋水仙素處理的‘Dakota小孢子母細(xì)胞經(jīng)減數(shù)分裂后,在正常四分體(圖4-a)周圍,還觀察到了較多的三分體(圖4-b)和二分體(圖4-c)。為了確定三分體和二分體的形成機(jī)制,采集經(jīng)秋水仙素處理后的肉穗花序,觀察減數(shù)分裂過程中染色體的異常行為,發(fā)現(xiàn)了三極紡錘體(圖4-d、e)和融合紡錘體(圖4-f、g)。三極紡錘體(圖4-d、e)和融合紡錘體(圖4-f、g)導(dǎo)致三分體和二分體的形成。此外還觀察到4n小孢子母細(xì)胞(圖4-h),該細(xì)胞的染色體數(shù)明顯大于30條,約60條。也就是說,2n小孢子母細(xì)胞被加倍為4n小孢子母細(xì)胞,經(jīng)減數(shù)分裂,同樣能形成2n花粉。
3 討論與結(jié)論
觀察‘Dakota小孢子母細(xì)胞減數(shù)分裂過程發(fā)現(xiàn),前期I持續(xù)時(shí)間最長,用時(shí)約占減數(shù)分裂過程的90%,進(jìn)一步證明前期I是減數(shù)分裂過程中最關(guān)鍵且復(fù)雜的一個(gè)階段[4]。本研究發(fā)現(xiàn)‘Dakota小孢子母細(xì)胞減數(shù)分裂進(jìn)程到達(dá)細(xì)線末期后,在一個(gè)花藥中,減數(shù)分裂有明顯的不同步性,常能同時(shí)觀察到4~8個(gè)不同的減數(shù)分裂相,甚至能觀察到從偶線期到四分體時(shí)期的所有減數(shù)分裂相。‘Dakota小孢子母細(xì)胞減數(shù)分裂的不同步性可以延長花期,增加花粉的有效作用時(shí)間,有利于種群繁殖后代[10]。
細(xì)胞的尺寸是隨著DNA含量的增加而增加的,DNA含量與細(xì)胞體積呈正相關(guān)[11]。花粉的倍性水平能根據(jù)花粉粒的尺寸來判斷。2n花粉的直徑比n花粉大30%左右,在作物甘薯、芭蕉、玫瑰柿子中已經(jīng)證實(shí)[12-15],本研究結(jié)果發(fā)現(xiàn)‘Dakota2n花粉直徑比n花粉大40%。‘Dakota的花粉近似球形,所以可以根據(jù)測量的直徑計(jì)算花粉的體積。
2n花粉是減數(shù)分裂失調(diào)產(chǎn)生的。根據(jù)2n花粉形成的遺傳機(jī)理,把2n花粉分為FDR(first division restitution)型2n花粉和SDR(second division restitution)型2n花粉[11]。FDR型2n花粉,除了重組片段,染色體與體細(xì)胞是一致的,具有較高的雜合性,能高效的傳遞親本雜合性和上位性,具有很高的育種價(jià)值。SDR型2n花粉含有母本一半的染色體,每條染色體都是雙份,雜合性較低。減數(shù)分裂結(jié)束后,三分體和二分體在四分體的周圍被發(fā)現(xiàn),表明三分體和二分體與2n花粉的形成有直接關(guān)系。1個(gè)四分體形成4個(gè)n花粉;1個(gè)三分體形成1個(gè)2n花粉和2個(gè)n花粉;1個(gè)二分體形成2個(gè)2n花粉。本試驗(yàn)通過對(duì)誘導(dǎo)過程的小孢子母細(xì)胞減數(shù)分裂進(jìn)程觀察表明,三極紡錘體和融合紡錘體是發(fā)生在第一次減數(shù)分裂失敗導(dǎo)致細(xì)胞學(xué)異常現(xiàn)象,這些異常現(xiàn)象是第一次減數(shù)分裂復(fù)原形成的,而第二次減數(shù)分裂正常進(jìn)行。因此,導(dǎo)致形成的2n花粉是FDR型2n花粉。
參考文獻(xiàn)
[1] Sheffer R D, Croat T B. Chromosome numbers in the genus Anthurium(Araceae)Ⅱ[J]. American Journal of Botany, 1983, 70: 858-871.
[2] Mari Marutani, Sheffer R D, Kamemoto H. Cytological analysis of Anthurium andraeanum(Araceae), Its related taxa and their hybrids[J]. American Journal of Botany, 1993, 80(1): 93-103.
[3] Wolverton B C, Rebecca C McDonald, Watkins E A Jr. Foliage plants for removing indoor air pollutants from energy-efficient homes[J]. Economic Botany, 1984, 38(2): 224-228.
[4] Bogdanov Y F. Variation and evolution of meiosis[J]. Russian Journal of Genetics, 2003, 39(4): 363-381.
[5] Otto SP, Whitton J. Polyploid incidence and evolution[J]. Annual Review of Genetics, 2000, 34: 401-437.
[6] Harlan J, De Wet, J.w.j-On O. Winge and a prayer:the origins of polyploidy[J]. The Botanical Review, 1975, 41(4):361-390.
[7] Ramanna M S, Jacobsen E. Relevance of sexual polyploidization for crop improvement-a review[J]. Euphytica,2003, 133(1): 3-18.
[8] 廖瑋瑋,邢虎成,揭雨成. 植物2n配子誘導(dǎo)及其應(yīng)用研究進(jìn)展[J]. 作物研究, 2011, 25(6): 599-603.
[9] Younis A, Hwang Y J, Lim K B. Exploitation of induced 2n-gametes for plant breeding[J]. Plant Cell Rep, 2011, 33(2):215-223.
[10] Wyatt R. Inflorescence architecture: How flower number,arrangement,and phenology affect pollination and fruit-set[J]. American Journal ofBotany, 1982, 69(4): 585-594.
[11] Bretagnolle F,Thompson J D. Gametes with the somatic chromosome number: mechanisms of their formation and role in the evolution of autoploid plants[J]. New Phytologist, 2005, 129(1): 1-22.
[12] Becerra Lopez-Lavalle L A, Orjeda G. Occurrence and cytological mechanism of 2n pollen formation in a tetraploid accession of Ipomoea batatas(sweet potato)[J]. Journal of Heredity, 2002, 93(3): 185-192.
[13] Ortiz R. Occurrence and inheritance of 2n pollen in Musa[J]. Annals of Botany, 1997, 79(4): 449-453.
[14] Crespel L, Ricci S C,Gudin S. The production of 2n pollen in rose[J]. Euphytica, 2006, 151(2): 155-164.
[15] Sugiura A, Ohkuma T, Choi Y A, et al. Production of nonaploid(2n=9x)japanese persimmons(Diospyros kaki)by pollination with unreduced(2n=6x) pollen and embryo rescue culture[J]. Journal of the American Society for Horticultural Science, 2000, 125(5): 609-614.
