貴州地方豬種及三元商品豬氟烷基因的檢測
2015-10-25 02:57:56馮文武龍威海丁玫陳祥陳村年
生物技術通報 2015年8期
關鍵詞:檢測
馮文武龍威海丁玫陳祥,陳村年
(1.貴州大學動物科學學院 高原山地動物遺傳育種與繁殖省部共建教育部重點實驗室,貴陽 550025;2.貴州省生豬健康養殖工程技術研究中心,貴陽 550025;3.貴州省畜牧獸醫學校,貴陽 550018)
貴州地方豬種及三元商品豬氟烷基因的檢測
馮文武1龍威海1丁玫3陳祥1,2陳村年2
(1.貴州大學動物科學學院高原山地動物遺傳育種與繁殖省部共建教育部重點實驗室,貴陽550025;2.貴州省生豬健康養殖工程技術研究中心,貴陽550025;3.貴州省畜牧獸醫學校,貴陽550018)
旨在對貴州地方豬種“抗應激敏感基因專門化品系”的培育提供技術支撐,運用PCR-RFLP技術對白洗豬、黔北黑豬、宗地花豬、糯谷豬和從江香豬5個地方豬種及三元商品豬共151份樣本進行了氟烷基因檢測,篩選出豬群中應激敏感基因隱性攜帶型HalNn、抗應激敏感基因型HalNN和應激敏感基因型Halnn。結果表明,貴州5個地方豬種全為HalNN型;三元商品豬群無Halnn型,HalNn型檢出率為52.63%,Haln型等位基因頻率為26.32%。
氟烷基因;RYR1基因檢測;繁殖力;PSE肉;肉質
豬肉品質已經被列入豬的品種選育的重要技術指標,劣質豬肉表現為PSE(Pale,soft,exudative)肉 和DFD(Dark,firm,dry) 肉 豬。 自1953年Ludvigsen首次提出PSE豬肉概念以來,在世界各地均有類似報道[1]。美國PSE豬肉發生率為5%-20%,日本為8.5%-11.5%[2],丹麥為10%-15%[3]。我國各地PSE豬肉發生率在10%-30%之間,個別地區高達60%-70%。豬骨骼肌蘭尼定受體(Ryanodine receptor 1,RYR1)基因的有害突變型為氟烷基因(halothane,Haln),是導致豬應激綜合征(Porcine stress syndrome,PSS)、發生惡性高溫綜合癥狀(malignant hyperthermia syndrome,MHS)、產生PSE肉或DFD肉的遺傳基礎[4],由于蘭尼定受體cDNA的C1843→T1843突變使蘭尼定受體蛋白結構和功能的改變,導致了劣質豬肉的產生[5]。現大量研究表明,氟烷有害基因還對豬的繁殖力有影響,Haln型相對于HalN型在繁殖性能上表現為一定的劣勢[6]。在我國大多數地方豬種未見報道有Haln基因[7-9],僅少數品種有Haln基因存在,如民豬Haln基因頻率為15.62%[9]。隨著人們生活水平的不斷提高,消費者對肉質提出了更高的要求,豬肉不僅要衛生,而且還要瘦肉率高、口感好,生產優質豬肉已迫在目睫。因此,對豬進行基因診斷,對PSE豬肉和DFD豬肉的形成機理進行探討,找出有效的防控措施,建立“抗應激敏感基因專門化品系”,具有重要的經濟價值和實際意義。本研究針對貴州省5個地方豬種及較為普及的三元商品豬生產基地進行氟烷基因檢測研究,旨為貴州地方豬種“抗應激敏感基因專門化品系”的培育提供技術支撐。
1 材料與方法
1.1 材料
白洗豬36份血樣采集于貴州省施秉縣,地方豬種DNA樣黔北黑豬14份、糯谷豬22份、宗地花豬31份、從江香豬29份皆為本實驗室保存樣,三元商品豬19份血樣采集于貴陽市某種豬場。
血液基因組柱式小量提取試劑盒與2×Es Taq MasterMix購于康為世紀生物科技公司,限制性內切酶Hha I購于寶生生物工程(大連)有限公司,DNA Marker購于生工生物工程(上海)有限公司,引物合成由生工生物工程(上海)有限公司完成。
1.2 方法
1.2.1 PCR擴增 按血液基因組柱式小量提取試劑說明書提取DNA后,擴增RYR1基因的DNA片段。引物設計[5],上游引物:5'-TCCAGTTTGCCACAGGTCCTACCA-3';下游引物:5'-ATTCACCGGAGTGGAGTCTCTGAG-3'。欲擴增產物是含RYR1基因cDNA第1 843位點長度為660 bp的片段。PCR反應體系20 μL:2×Es Taq MasterMix 10 μL,上、下游引物各0.8 μL(10 μmol/L),基因組DNA模板2 μL,加滅菌水至終體積20 μL。PCR擴增條件:95℃預變性5 min,95℃變性30 s,55-65℃退火30 s,72℃延伸30 s,35個循環;72℃延伸7 min,4℃保存。擴增產物經1%瓊脂糖凝膠電泳檢測。
1.2.2 酶切反應 酶切反應體系50 μL:10×Quick Cut Green Buffer 3 μL,10 μL PCR產物,限制性內切酶Hha I 1 μL,加滅菌水至終體積30 μL。混勻試劑后瞬時離心,37℃水浴酶切10 min。酶切產物經1%瓊脂糖凝膠電泳檢測。
2 結果
2.1 PCR反應條件優化
通過DNA池技術摸索最佳PCR反應條件,RYR1基因cDNA含第1 843位點長度為660 bp(全序列17 721 bp-18 381 bp片段)的片段在20 μL PCR反應體系中,退火溫度63℃反應條件下擴增良好。對各樣分別進行PCR擴增,擴增出660 bp的目的條帶(圖1)。
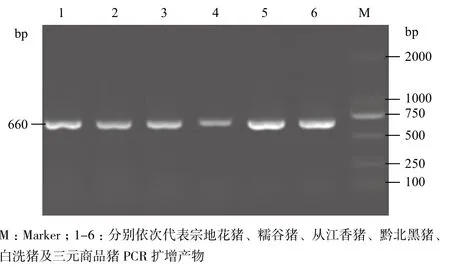
圖1 各豬種RYR1基因PCR產物
2.2 RYR1基因RFLP分析
Hha I內切酶的識別位點為5'-GCG↓C-3'。PCR擴增產物是含RYR1基因cDNA第1 843位點長度為660 bp的片段,根據電泳圖譜中的帶型判斷氟烷基因型。如果豬的基因型為HalNN,由于胞嘧啶未突變成胸腺嘧啶,擴增產物被酶切為494 bp和166 bp,因此電泳形成兩條帶;如果基因型為Halnn時,發生基因突變的酶切位點消失,Hha I內切酶不能識別,對PCR產物不能酶切,所以只能看到660 bp 1條帶;如果豬的基因型為HalNn雜合子,擴增產物則可以電泳形成660 bp、494 bp和166 bp 3條帶[11]。酶切產物經1%瓊脂糖凝膠電泳,可以觀察到RYR1基因的DNA酶切片段(圖2)。經檢驗,三元商品豬19份樣品中檢測出10份樣品RYR1基因型為HalNn、9份樣品為HalNN、無Halnn。
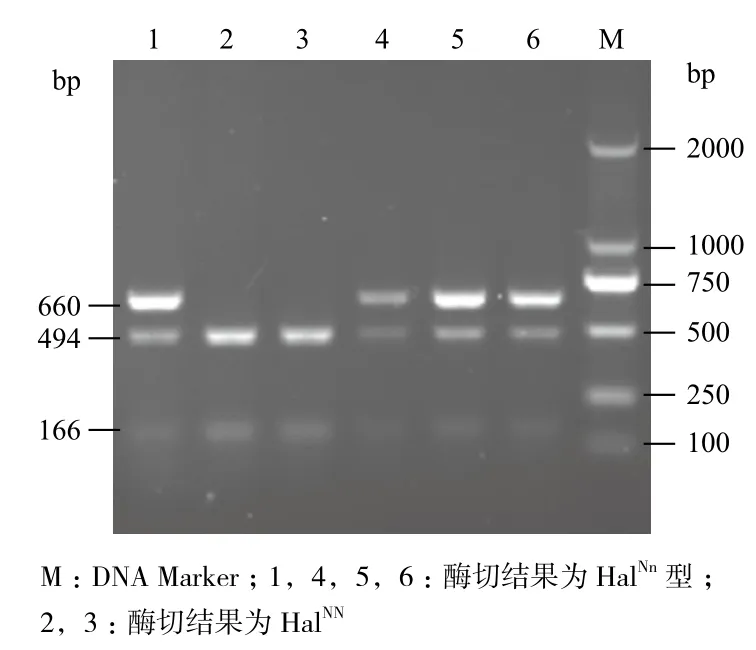
圖2 三元商品豬PCR產物酶切結果
2.3 RYR1基因C1843→T1843突變的基因型頻率和基因頻率
對白洗豬、黔北黑豬、宗地花豬、糯谷豬及從江香豬5個貴州省地方豬種132份樣品進行了檢測,全為RYR1基因抗應激敏感HalNN型,三元商品豬19份樣品中檢測到兩種基因型,應激敏感基因隱性攜帶型HalNn與抗應激敏感HalNN型,未檢測出應激敏感基因型Halnn,RYR1基因cDNA C1843→T1843突變的基因型頻率和基因頻率,如表1所示。

表1 三元商品豬RYR1基因C1843→T1843突變的基因型頻率和基因頻率
3 討論
3.1 氟烷基因檢測在生產中的引物設計
氟烷基因檢測技術在指導生產過程中引物設計至關重要,引物設計可根據GenBank中的豬RYR1基因序列進行設計,亦可參考已發表研究報告中提供的參考序列。常見的氟烷基因3個引物設計序列見表2,本研究采用Fujii等[5]設計的引物,引物在PCR擴增體系中最適溫度條件為63℃。
3.2 RYR1基因型頻率和基因頻率對比
在近年的研究報道中,國外引進豬種n型等位基因頻率較高,皮特蘭豬、英國大白豬、臺灣杜洛克豬、丹麥杜洛克、迪卡豬C系、迪卡豬F系的Haln型等位基因頻率分別為100%、12.5%、5.3%、4.6%、8.3%及4.2%[13]。本實驗對5個貴州省地方豬種及三元商品豬共151份樣本的RYR1基因限制性內切酶片段長度多態性進行了檢測。結果表明,5個貴州省地方豬種中全為抗應激敏感基因HalNN型,三元商品豬群中檢測到10例應激敏感基因隱性攜帶型HalNn,占豬群體的52.63%(n=19),Haln型等位基因頻率為26.32%。結果中三元商品豬Haln型等位基因與國外豬種相比,國外種豬Haln型等位基因頻率保持較高水平;與朱鋒釗[14]報道的外三元豬群RYR1基因HalNn型比例為11.11%、Haln型等位基因頻率為5.56%相比,HalNn型基因比例上升了41.52%,Haln型等位基因頻率上升了27.62%,HalNn型基因在試驗三元商品豬群中的比例呈明顯上升趨勢;本實驗5個貴州本地豬種檢測全為抗應激敏感基因HalNN型,與朱鋒釗[14]報道相同,且首次對貴州省優良地方豬種白洗豬、黔北黑豬進行了RYR1基因檢測,二者皆為抗應激敏感基因HalNN型。
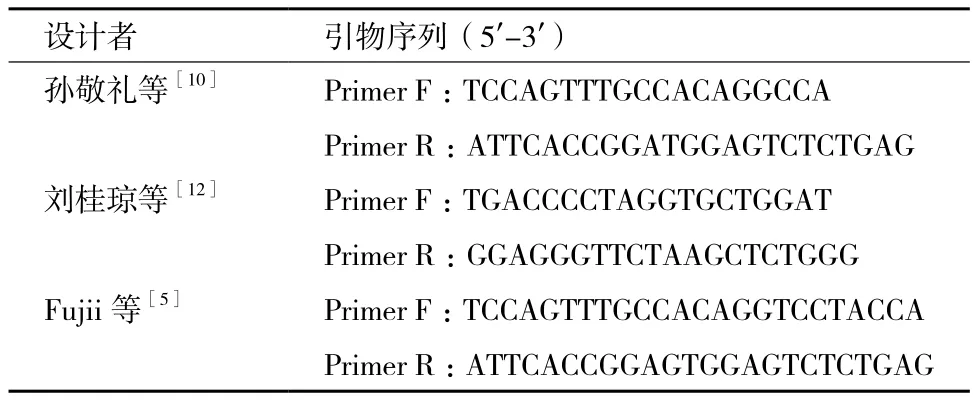
表2 常見的3對引物設計序列
3.3 RYR1基因對肉質的影響
近來的研究表明,Haln等位基因的出現與歐洲豬種的高瘦肉率選育直接相關[15],RYR1基因HalNn型豬與HalNN型比較,HalNn型豬骨骼肌應激敏感性高,應激易使肌細胞肌漿網終池中的鈣離子大量快速地釋于肌漿,引起肌細胞快速收縮,隨著ATP降解供能,鈣離子又被泵回肌漿網終池,肌漿內鈣離子濃度下降,肌細胞轉入靜息期。含Haln型等位基因豬遇到應激時,如此反復刺激肌細胞并促使其生長,肌肉生長的需求增加致使Haln型豬對飼料轉化成瘦肉的需求增加,瘦肉比重亦隨之增大,故Haln型豬比正常豬肌肉更為發達。RYR1基因的n型等位基因與高瘦肉率正相關,但也具高的PSE發生率。據研究,RYR1基因Halnn型豬即使采用最優處理,其產生PSE肉的比率也高達80%以上,RYR1基因HalNn型豬產生各種劣質肉的比率達到36.8%,顯著高于HalNN型豬22.5%的水平[16]。
3.4 RYR1基因對繁殖力的影響及效益分析
杜立新等[17]報道,HalNn型母豬的產活子數比Halnn型多1.74頭。Stalder等[18]對某應激敏感合成系研究表明,HalNN型的21日齡成活率則比HalNn型高出9.33%(P<0.05)。蔣思文等[19]報道,HalNN型和HalNn型豬的窩總產仔數、窩產活仔數、初生窩重及育成數、斷奶窩重均極顯著高于Halnn型豬(P<0.01),斷奶頭數HalNN顯著高于Halnn型(P<0.05)。劉月環[20]報道,HalN基因對總產仔數、20日齡仔豬成活率、斷奶前后仔豬的生長效應顯著(P<0.05)。總的來說,Halnn型相對于HalNN和HalNn型在繁殖性能上表現出一定的劣勢。劉桂瓊等[12]研究表明,Haln基因對產仔數的遺傳效應影響較大,凡帶有RYR1基因Haln型的母豬,其產仔數、產活仔數比純合HalNN母豬少,從頭3胎總平均數看,Halnn母豬比HalNN母豬窩產仔數、產活仔數、斷奶仔數少1.92、1.72和1.57頭,HalNn母豬比HalNN母豬少0.80、0.84和0.72頭。本研究商品群體中HalNn、HalNN、Halnn型比例為52.63%、47.37%和0。根據孟德爾遺傳定律可測算出該商品豬群父母代豬群基因型為HalNN、HalNn、Halnn,基因型比例閾值為0-47.37%、0-94.74%、5.26%-52.6%,按試驗場300頭基礎母豬,利用年限8胎次,可計算出該場僅因RYR1基因對繁殖性能的影響,至少損失808.55頭豬仔。若對該場300頭基礎母豬及公豬進行RYR1基因檢測,約花費檢測成本0.5萬元,對該場進行氟烷基因篩選投資回報可觀。
3.5 豬場氟烷基因型的凈化
氟烷有害基因Haln型等位基因屬氟烷敏感基因,Fujii等[5]對氟烷基因的克隆分析和比較正常豬與應激豬蘭尼定受體cDNA序列發現,蘭尼定受體cDNA的C1843→T1843突變使蘭尼定受體蛋白結構和功能的改變。這種改變導致劣質豬肉的產生[4]。氟烷基因一方面導致肌細胞肥大,提高胴體瘦肉率,另一方面易誘導發生惡性高熱綜合征產生PSE肉,降低肌肉品質,隨氟烷基因n型頻率的增加,豬的肉質性狀、生長發育性狀和繁殖性狀都逐漸變劣[21],已給世界養豬生產造成了巨大損失。在常規育種過程中,只是淘汰掉有應激綜合征表現的隱性純合個體,而讓雜合體自由繁殖,那么隱性基因頻率在選育前幾年降低很快,以后則越來越緩慢,難以完全消除,而氟烷基因雜合子豬在生產性能和體型外貌上具有雜合優勢,通過常規選種技術更易被選作種用[10]。本研究對三元商品豬19份樣本的RYR1基因多態性檢測結果說明,在豬群的父母代種豬中一定含有RYR1基因HalNn型豬、HalNN型和Halnn型。從整個養豬生產綜合來看,Haln等位基因具降低產出的效應,因此有必要應用目前各種檢測手段指導實際生產,消除氟烷隱性及雜合有害基因,在種豬群中淘汰這一基因,建立“抗應激敏感基因專門化品系”,達到豬場氟烷應激敏感基因型的凈化,最終生產出優質的豬肉。
4 結論
本實驗利用PCR-RFLP法對白洗豬、黔北黑豬、宗地花豬、糯谷豬、從江香豬5個地方豬種及三元商品豬共151份樣本進行了RYR1基因檢測,首次對黔北黑豬和白洗豬RYR1基因檢測進行報道,5個地方豬種均不含氟烷敏感Haln型等位基因。本實驗三元商品豬群中HalNn基因型占豬群體的52.63%(n=19),Haln型等位基因頻率為26.32%,HalNn基因型在實驗豬群中的比例較大。建議運用PCRRFLP法對基礎母豬RYR1基因型凈化,在種豬群淘汰RYR1基因Haln型等位基因,最終建立“抗應激敏感基因專門化品系”。
[1]丁玉庭, 劉化章, 周曉云. 豬PSE肉與正常肉的肌原纖維形態學比較研究[J]. 中國食品學報, 2005, 5(2):28-33.
[2]韓杰, 邊連全, 曹新民. PSE豬肉的發生機制及調控措施[J].黑龍江畜牧獸醫, 2011, 4:33-34.
[3]李建穎. 上海地區種豬氟烷基因的檢測[D]. 杭州:浙江大學,2006.
[4]帥素容, 李學偉, 趙秀娟, 朱礪. 中國6個地方豬種與3個外種豬氟烷基因PCR產物序列比較研究[J]. 中國畜牧雜志,2005, 2:14-17.
[5] Fujii J, Otsu K, Zorzato F, et al. Identification of a mutation in porcine ryanodine receptor associated with malignant hyperthermia[J]. Science, 1991, 253(5018):448-451.
[6]趙中權. 藏豬氟烷基因, 雌激素受體基因和生長激素基因PCRRFLP分析[D]. 雅安:四川農業大學, 2005.
[7]陳蘊穎, 步宏, 李幼平, 等. 中國內江豬的氟烷基因型[J]. 華西醫科大學學報, 1999, 2:5-7.
[8]方美英, 姜志華, 劉紅林, 等. 不同豬種中氟烷基因頻率調查分析[J]. 浙江農業學報, 1999, 3:39-41.
[9]李來記, 張沅, 李寧. 豬惡性高熱綜合征(MHS)基因群體檢測及部分片段DNA序列研究[J]. 中國獸醫學報, 1996, 16(6):540-545.
[10]孫敬禮, 武磊, 李大全, 等. 新疆某規模化豬場氟烷基因的檢測[J]. 豬業科學, 2011, 1:96-98.
[11]張金玲, 吳海洪, 錢永華, 等. PCR-RFLP技術檢測新嘉興黑豬氟烷基因[J]. 中國畜牧獸醫, 2008, 09:133-134.
[12]劉桂瓊, 丁家桐, 姜勛平, 張牧. RYR1基因對母豬繁殖性能的影響[J]. 中國獸醫學報, 2002, 22(4):392-394.
[13]姚德標, 廖波, 謝丕雄, 等. 引進種豬氟烷敏感基因檢測結果報告[J]. 養豬, 2002, 4:30.
[14]朱鋒釗. 七種貴州地方豬品種的三種遺傳缺陷主效基因多態性研究[D]. 貴陽:貴州大學, 2009.
[15]?echová M, Wolf J, Tr?ka P. Impact of RYR1 genotype of Piétrain boars on litter traits of Czech Large White×Czech Landrace crossbred sows[J]. Journal of Animal Breeding and Genetics,2007, 124(2):86-93.
[16]Christian L. Clarifying the impact of the stress gene[J]. Natl Hog Farmer, 1995, 40(6):44-46.
[17]杜立新, 王愛華, 姜運良, 張金花. 豬RYR1基因的檢測及其對生產性能的遺傳效應[J]. 畜牧獸醫學報, 2001, 32(6):481-486.
[18]Stalder KJ. Effects of porcine stress syndrome genotype on maternal traits in swine[D]. Iowa State University, 1995.
[19]蔣思文, 鄧昌彥, 熊遠著, 汪紹南. 氟烷敏感性與繁殖性能關系的研究[J]. 中國畜牧雜志, 1995, 31(5):20-21.
[20] 劉月環. 氟烷基因與 FUT1 基因對豬生產性能效應的研究[D].杭州:浙江大學, 2001.
[21]徐小波, 劉鐵錚, 許康樸, 葛云山. 蘇鐘Ⅰ系和Ⅱ系豬的氟烷基因及其對產肉性能的影響[J]. 中國獸醫學報, 2000, 20(4):397-400.
(責任編輯 狄艷紅)
Detection of the RYR1 Gene in Guizhou Indigenous Pigs and Three-crossbred Pigs
Feng Wenwu1Long Weihai1Ding Mei3Chen Xiang1,2Chen Cunnian2
(1. Key Laboratory of Animal Genetics,Breeding and Reproduction in the Plateau Mountainous Region,Ministry of Education,College of Animal Sciences,Guizhou University,Guiyang550025;2. Guizhou Engineering Technology Research Center for Healthy Pig Breeding,Guiyang550025;3. Guizhou Husbandry Animal Science Vocational School,Guiyang550018)
To provide the technical support for the cultivation of "strain of specialization with anti-stress and sensitive gene" of Guizhou local pig genes, the RYR1 gene in 151 samples from 5 local pig species of Baixi, Qianbei Black, Zongdi Hua, Nuogu, Congjiang Xiang and threecrossbred commercial pig were detected by PCR-RFLP technique. The pigs with HalNn of recessive anti-stress and sensitive genotype, HalNN of anti-stress and sensitive genotype and Halnn of stress and sensitive genotype were screened out. The results showed that all of 5 Guizhou indigenous pigs belonged to HalNN , there was no Halnn gene in the three-crossbred pigs, HalNn accounted for 52.63 % of the samples, and Haln allele frequency was 26.32%.
halothane gene;detection of RYR1 gene; fertility;PSE pork; meat quality
10.13560/j.cnki.biotech.bull.1985.2015.08.010
2014-09-22
貴州省農業動植物育種專項項目[黔農育專字(2011)019號],貴州省農業攻關項目[黔科合NY(2013)3071號],貴州省工程中心建設項目[黔科合農C字(2011)4022號]
馮文武,男,碩士研究生,研究方向:動物繁殖與動物生物技術;E-mail:15285116946@139.com
陳祥,博士,教授,碩士生導師,研究方向:動物繁殖技術與動物生物技術;E-mail:as.xchen@ gzu.edu.cn
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
