重組精氨酸脫亞胺酶制備L-瓜氨酸的工藝條件優(yōu)化
2015-10-25 02:58:14馬越宿玲恰吳丹吳敬
生物技術(shù)通報(bào) 2015年8期
馬越宿玲恰吳丹吳敬
(1.江南大學(xué) 食品科學(xué)與技術(shù)國(guó)家重點(diǎn)實(shí)驗(yàn)室,無(wú)錫 214122;2.江南大學(xué)生物工程學(xué)院 工業(yè)生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室,無(wú)錫 214122)
重組精氨酸脫亞胺酶制備L-瓜氨酸的工藝條件優(yōu)化
馬越1,2宿玲恰1,2吳丹1,2吳敬1,2
(1.江南大學(xué)食品科學(xué)與技術(shù)國(guó)家重點(diǎn)實(shí)驗(yàn)室,無(wú)錫214122;2.江南大學(xué)生物工程學(xué)院工業(yè)生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室,無(wú)錫214122)
將來(lái)源于Pseudomonas putida ACCC 10185的ADI編碼基因克隆到表達(dá)載體pET-24a(+)中,轉(zhuǎn)化Escherichia coli BL21(DE3),通過(guò)超聲波破碎得到粗酶液,酶活檢測(cè)ADI酶活為26 U/mL發(fā)酵液。對(duì)酶轉(zhuǎn)化L-精氨酸鹽酸鹽生成L-瓜氨酸的反應(yīng)條件進(jìn)行了優(yōu)化,結(jié)果表明,當(dāng)?shù)孜風(fēng)-精氨酸鹽酸鹽濃度650 g/L,反應(yīng)初始pH6.0,溫度37℃,加酶量24 U/g底物,轉(zhuǎn)速100-200 r/min,轉(zhuǎn)化時(shí)間7 h,L-瓜氨酸轉(zhuǎn)化率達(dá)到100%,是目前國(guó)內(nèi)外報(bào)道的酶法制備L-瓜氨酸的最高水平。
精氨酸脫亞胺酶;L-瓜氨酸;酶轉(zhuǎn)化;重組表達(dá)
L-瓜氨酸是一種游離的氨基酸,是人體尿素循環(huán)的重要中間代謝物,具有多種生理功能。利尿、護(hù)肝,可與L-鳥(niǎo)氨酸、L-精氨酸等共同治療高氨血癥[1,2];參與內(nèi)源性細(xì)胞舒張因子一氧化氮的合成,有助于擴(kuò)張血管、抑制動(dòng)脈硬化,可用于治療高血壓、冠心病及男性性功能障礙等疾病[3-5];在腸道中L-瓜氨酸通過(guò)谷氨酸-鳥(niǎo)氨酸代謝途徑合成,可作為檢驗(yàn)小腸移植時(shí)異體排斥效應(yīng)程度的指示劑[6];L-瓜氨酸與精氨酸結(jié)構(gòu)相似,對(duì)體內(nèi)自由基表現(xiàn)出高度的活性,可作為天然高效的新抗氧化劑[7],由此可見(jiàn)L-瓜氨酸在醫(yī)藥用品、保健食品以及化妝品中應(yīng)用前景廣闊。隨著人們保健意識(shí)的提高,對(duì)L-瓜氨酸的需求量日益增多,因此生產(chǎn)L-瓜氨酸具有重大的意義。
L-瓜氨酸主要是由精氨酸脫亞胺酶(arginine deiminase,ADI,EC3.5.3.6)催化L-精氨酸鹽酸鹽水解生成L-瓜氨酸和氨[8,9]。利用ADI催化生成L-瓜氨酸由于原料易得、工藝簡(jiǎn)單、產(chǎn)物濃度高等優(yōu)點(diǎn)逐漸成為L(zhǎng)-瓜氨酸制備的潛力途徑。
本研究將來(lái)源于Pseudomonas putida ACCC 10185[10]的ADI基因(arcA)連接到表達(dá)載體pET-24a(+),獲得重組質(zhì)粒pET-24a(+)-arcA,將其轉(zhuǎn)化Escherichia coli BL21(DE3)并進(jìn)行搖瓶發(fā)酵,經(jīng)超聲波破碎,檢測(cè)破壁上清液中的ADI 酶活。在此基礎(chǔ)上,對(duì)酶轉(zhuǎn)化工藝進(jìn)行研究,旨在獲得高效制備L-瓜氨酸的方法。
1 材料與方法
1.1 材料
1.1.1 菌株和質(zhì)粒 惡臭假單胞菌(Pseudomonas putida ACCC 10185):由ACCC菌種保藏中心購(gòu)買;E.coli JM109、E.coli BL21(DE3)、表達(dá)載體pET-24a(+):為本實(shí)驗(yàn)室保藏;克隆載體pMD18-T Simple vector:購(gòu)自TaKaRa公司。
1.1.2 培養(yǎng)基 LB培養(yǎng)基(g/L):蛋白胨10,酵母粉5,NaCl 10,卡那霉素0.03,pH7.0。
TB培養(yǎng)基(g/L):蛋白胨12,酵母粉24,甘油5,K2HPO42.31,KH2PO416.43, 卡 那 霉 素0.03,pH7.0。
1.1.3 試劑 限制性內(nèi)切酶、T4 DNA連接酶、PrimerStar HS DNA聚合酶、堿性磷酸酶CIAP、核酸分子量標(biāo)準(zhǔn)及瓊脂糖:購(gòu)自TaKaRa公司;PCR產(chǎn)物純化試劑盒、瓊脂糖凝膠DNA回收試劑盒、質(zhì)粒小提試劑盒:天根生化科技有限公司;卡那霉素、氨芐青霉素、異丙基-β-D-硫代半乳糖苷(IPTG)、蛋白質(zhì)相對(duì)分子質(zhì)量標(biāo)準(zhǔn)、L-精氨酸鹽酸鹽、瓜氨酸標(biāo)樣:購(gòu)自生工生物工程(上海)有限公司;其它分析純?cè)噭┚?gòu)自國(guó)藥集團(tuán)化學(xué)試劑有限公司。
1.1.4 主要儀器 凝膠成像儀、蛋白電泳儀購(gòu)自美國(guó)Bio-Rad公司;DYY-6C型核酸電泳儀購(gòu)自北京六一儀器廠;高效液相色譜儀購(gòu)自Agilent公司;細(xì)胞破碎儀購(gòu)自北京科實(shí)興業(yè)科技有限公司;紫外可見(jiàn)光分光光度計(jì)購(gòu)自日本Shimadzu公司。
1.2 方法
1.2.1 工程菌的構(gòu)建 提取Pseudomonas putida ACCC 10185的基因組,PCR擴(kuò)增目的基因arcA,將所得片段割膠回收后,連接克隆載體pMD18-T Simple vector,將連接產(chǎn)物轉(zhuǎn)入E.coli JM109感受態(tài)細(xì)胞中,在平板培養(yǎng)基上培養(yǎng)過(guò)夜,挑取單克隆至液體培養(yǎng)基中培養(yǎng),抽提質(zhì)粒得到pMD18-T-arcA,經(jīng)NdeⅠ和Hin d Ⅲ雙酶切后,將目的基因片段割膠回收,T4連接酶將目的基因與經(jīng)相同酶切處理的表達(dá)載體pET-24a(+)連接,連接產(chǎn)物轉(zhuǎn)入E.coli JM109感受態(tài)細(xì)胞中,在平板培養(yǎng)基培養(yǎng)過(guò)夜后,挑取單克隆進(jìn)行液體培養(yǎng),抽提得到重組質(zhì)粒pET-24a(+)-arcA。將pET-24a(+)-arcA進(jìn)行雙酶切驗(yàn)證,驗(yàn)證正確后轉(zhuǎn)入E.coli BL21(DE3)感受態(tài)細(xì)胞中,涂布LB平板培養(yǎng)基,挑取單克隆進(jìn)行液體培養(yǎng),保存甘油管,-80℃保存。
1.2.2 搖瓶發(fā)酵生成ADI
1.2.2.1 種子培養(yǎng) 接一定量-80℃ 甘油管中保存的菌液至LB培養(yǎng)基,37℃、200 r/min,培養(yǎng)8 h。
1.2.2.2 搖瓶發(fā)酵 將種子液以5%的接種量轉(zhuǎn)接至TB培養(yǎng)基,37℃培養(yǎng)至OD600約為1.0時(shí),加入終濃度為0.4 mmol/L的IPTG[11,12],置于25℃、轉(zhuǎn)速200 r/min進(jìn)行重組蛋白的誘導(dǎo)表達(dá),誘導(dǎo)18 h后離心收集菌體,用50 mmol/L,pH6.0 Na2HPO4-NaH2PO4緩沖液復(fù)溶菌體至5 OD,超聲波破碎后離心,破壁上清即為ADI粗酶液,測(cè)定粗酶液中的ADI酶活,經(jīng)與發(fā)酵液OD折算后,即為每毫升發(fā)酵液中的ADI酶活。
1.2.3 ADI活性測(cè)定 利用二乙酰一肟-氨基硫脲比色法測(cè)定ADI酶活[13]。用50 mmol/L,pH6.0的磷酸鹽緩沖液配制0.2 mol/L L-精氨酸鹽酸鹽溶液作為底物,取100 μL適當(dāng)稀釋的酶液加入1 mL底物中,于37℃條件下反應(yīng)30 min,加入1 mL 10%三氯乙酸終止反應(yīng),沸水浴5 min,稀釋一定倍數(shù)后測(cè)定瓜氨酸含量,并計(jì)算酶活。酶活定義:37℃條件下,每分鐘轉(zhuǎn)化1 μmol L-精氨酸鹽酸鹽生成瓜氨酸的酶量定義為1 U。
1.2.4 L-瓜氨酸的制備及含量測(cè)定 以50 mmol/L,pH6.0的磷酸鹽緩沖液配制濃度為650 g/L的L-精氨酸鹽酸鹽溶液,加酶量為24 U/g底物,與一定量的酶液充分混勻后在37℃、100-200 r/min水浴搖床中反應(yīng)7 h。轉(zhuǎn)化后的反應(yīng)液加入等體積的三氯乙酸后,靜置2 h,12 000 r/min離心10 min,取上清進(jìn)行分析。利用HPLC進(jìn)行產(chǎn)物分析的色譜條件是:Agilent 1200 HPLC色譜儀,Agilent自動(dòng)進(jìn)樣器,色譜柱250×4.6 mm 5 μm Inertsil ODS-3 Column,Agilent紫外檢測(cè)器;流速0.8 mL/min;柱溫40℃。轉(zhuǎn)化率定義為生成的L-瓜氨酸的摩爾數(shù)/L-精氨酸鹽酸鹽的摩爾數(shù)×100%。
2 結(jié)果
2.1 ADI重組菌的構(gòu)建
2.1.1 重組質(zhì)粒pET-24a(+)-arcA的構(gòu)建 提取Pseudomonas putida ACCC 10185的基因組,PCR擴(kuò)增目的基因arcA,將攜帶目的基因的質(zhì)粒pMD18-T和本實(shí)驗(yàn)室質(zhì)粒pET-24a(+)分別經(jīng)Nde Ⅰ和Hin dⅢ雙酶切后,割膠回收目的基因片段和pET-24a(+),連接后轉(zhuǎn)入E.coli JM109感受態(tài)細(xì)胞,培養(yǎng)重組菌并抽提質(zhì)粒。重組質(zhì)粒經(jīng)Nde Ⅰ和Hin d Ⅲ雙酶切驗(yàn)證,結(jié)果(圖1)顯示,在大約5 300 bp和1 300 bp處有條帶,分別與載顯體及基因大小一致,表明重組表達(dá)載體pET-24a(+)-arcA構(gòu)建成功。
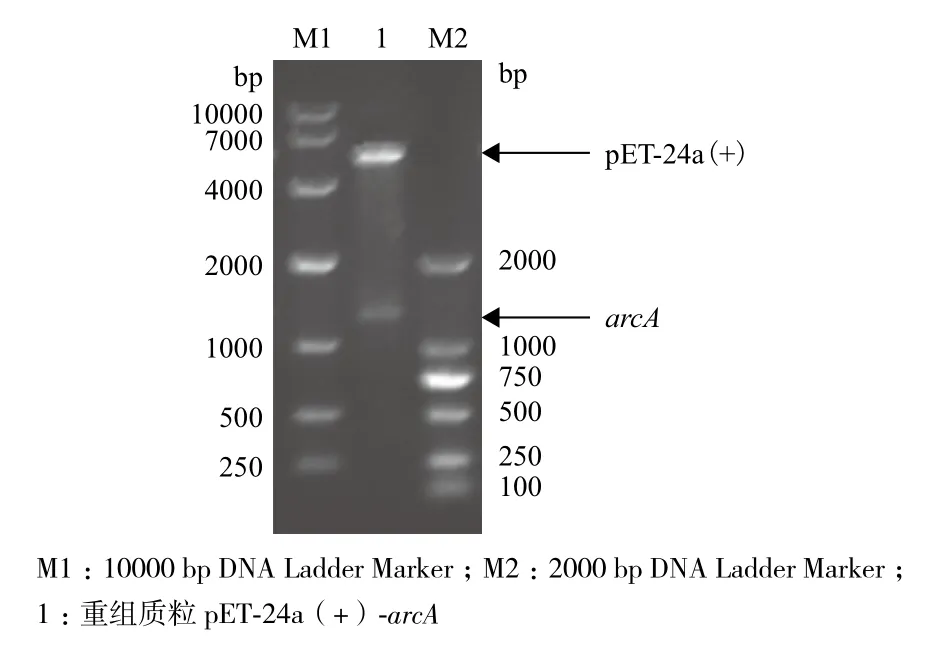
圖1 重組質(zhì)粒pET-24a(+)- arcA酶切驗(yàn)證
2.1.2 重組菌E.coli BL21(DE3)/ pET-24a(+)-arcA的構(gòu)建及培養(yǎng) 將重組質(zhì)粒pET-24a(+)-arcA轉(zhuǎn)化E.coli BL21(DE3)感受態(tài)細(xì)胞,經(jīng)0.4 mmol/L IPTG誘導(dǎo)培養(yǎng)后收集菌體,處理后得ADI粗酶液。經(jīng)檢測(cè),ADI酶活為26 U/mL發(fā)酵液。SDS-PAGE(圖2)分析顯示,ADI破壁上清液在大約43 kD處出現(xiàn)一條蛋白條帶,與理論ADI分子量相符,表明ADI在E.coli BL21(DE3)中成功表達(dá)。
2.2 重組ADI制備L-瓜氨酸轉(zhuǎn)化條件優(yōu)化
2.2.1 反應(yīng)溫度對(duì)酶轉(zhuǎn)化的影響 以650 g/L L-精氨酸鹽酸鹽為底物,加酶量為24 U/g底物,pH6.0,轉(zhuǎn)速100-200 r/min,分別在30℃、37℃、40℃、50℃、60℃和70℃水浴搖床中轉(zhuǎn)化7 h,HPLC檢測(cè)L-瓜氨酸生成量。圖3表明,高溫條件下轉(zhuǎn)化率較低,而低溫條件下轉(zhuǎn)化率明顯提高,最高值為100%。這是因?yàn)榈蜏赜欣诿傅姆€(wěn)定,而高溫時(shí)酶的熱穩(wěn)定性較差,反應(yīng)過(guò)程中酶易失活,導(dǎo)致轉(zhuǎn)化率較低,因此37℃為最佳轉(zhuǎn)化溫度。
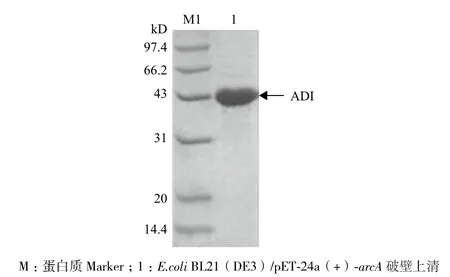
圖2 精氨酸脫亞胺酶SDS-PAGE分析
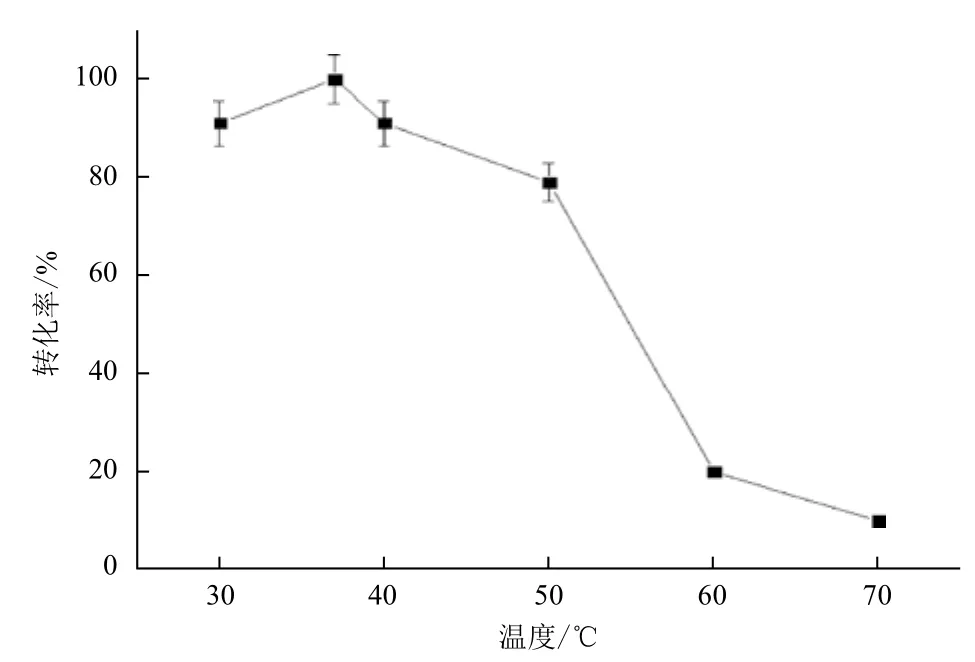
圖3 反應(yīng)溫度對(duì)酶轉(zhuǎn)化的影響
2.2.2 反應(yīng)初始pH對(duì)酶轉(zhuǎn)化的影響 以650 g/L L-精氨酸鹽酸鹽為底物,加酶量為24 U/g底物,反應(yīng)溫度37℃,轉(zhuǎn)速100-200 r/min,分別在pH4.0、5.0、5.5、6.0、6.5、7.0和8.0中轉(zhuǎn)化7 h,HPLC檢測(cè)L-瓜氨酸生成量。圖4表明,當(dāng)初始pH為6.0時(shí),L-瓜氨酸轉(zhuǎn)化率為100%。pH的改變可使酶分子上的側(cè)鏈基團(tuán)處于不同的解離狀態(tài),影響底物的結(jié)合和進(jìn)一步的反應(yīng),或者影響酶的空間結(jié)構(gòu),從而影響酶的活性,因此過(guò)酸或過(guò)堿對(duì)酶促反應(yīng)都不利,在pH為6.0時(shí)L-瓜氨酸轉(zhuǎn)化率最高。

圖4 反應(yīng)初始pH對(duì)酶轉(zhuǎn)化的影響
2.2.3 加酶量對(duì)酶轉(zhuǎn)化的影響 以100 g/L L-精氨酸鹽酸鹽為底物,加入不同量的ADI,加酶量分別為底物12、24和36 U/g,反應(yīng)溫度37℃,pH6.0,轉(zhuǎn)速100-200 r/min,轉(zhuǎn)化3.5 h,HPLC檢測(cè)L-瓜氨酸生成量。圖5表明,隨著加酶量的增多,相同時(shí)間內(nèi)L-瓜氨酸轉(zhuǎn)化率逐漸增大,但是不同加酶量條件下,L-瓜氨酸轉(zhuǎn)化率均能達(dá)到100%。考慮到經(jīng)濟(jì)性因素,選擇加酶量為底物24 U/g。

圖5 加酶量對(duì)酶轉(zhuǎn)化的影響
2.2.4 底物濃度與反應(yīng)時(shí)間對(duì)酶轉(zhuǎn)化的影響 分別配制濃度為50、100、150、200、300、400和650 g/L的L-精氨酸鹽酸鹽溶液,加酶量為24 U/g底物,反應(yīng)溫度37℃,pH6.0,轉(zhuǎn)速100-200 r/min,定時(shí)取樣,HPLC檢測(cè)L-瓜氨酸生成量。圖6表明,隨著L-精氨酸鹽酸鹽濃度的增大,相同時(shí)間內(nèi)L-瓜氨酸轉(zhuǎn)化率越來(lái)越低,但隨著轉(zhuǎn)化時(shí)間的延長(zhǎng),L-瓜氨酸的濃度不斷增加,最終均能達(dá)到100%,因此將轉(zhuǎn)化時(shí)間定為7 h。

圖6 底物濃度對(duì)酶轉(zhuǎn)化的影響
2.2.5 轉(zhuǎn)速對(duì)酶轉(zhuǎn)化的影響 以650 g/L L-精氨酸鹽酸鹽為底物,加酶量為24 U/g底物,反應(yīng)溫度37℃,pH6.0,分別在轉(zhuǎn)速100、150和200 r/min中轉(zhuǎn)化7 h,HPLC檢測(cè)L-瓜氨酸生成量。結(jié)果顯示,不同轉(zhuǎn)速對(duì)L-瓜氨酸轉(zhuǎn)化率無(wú)影響。
3 討論
早在1971年,來(lái)自日本的Kakimoto等[14]就利用Pseudomonas putida ATCC 4359 ADI生產(chǎn)瓜氨酸,以L-精氨酸鹽酸鹽為底物,反應(yīng)62 h后轉(zhuǎn)化率達(dá)到90.5%;1973年,Chibata等[9]報(bào)道同一來(lái)源的ADI,以L-精氨酸或DL-精氨酸為底物,可生產(chǎn)瓜氨酸濃度達(dá)80 g/L以上;1974年,Yamamoto等[15]用聚丙烯酰胺凝膠包埋上述來(lái)源的細(xì)胞,可將濃度0.5 mol/L的L-精氨酸鹽酸鹽完全轉(zhuǎn)化為L(zhǎng)-瓜氨酸;2014年,王穎等[16]將來(lái)源于Pseudomonas putida ATCC 4359的ADI在E.coli中克隆表達(dá),利用重組菌制備的粗酶液可將20 g/L L-精氨酸有效轉(zhuǎn)化為L(zhǎng)-瓜氨酸,轉(zhuǎn)化率高于90%,該酶顯示出了與來(lái)源于Pseudomonas putida ACCC 10185的ADI 99.76%的同源性。
我國(guó)對(duì)于其他來(lái)源的ADI轉(zhuǎn)化L-精氨酸鹽酸鹽生成L-瓜氨酸的研究也有一些報(bào)道。2005年,曹瑜等[17]利用Streptococcus faecalis NJ402游離細(xì)胞的ADI,在10 L轉(zhuǎn)化罐中,可將質(zhì)量濃度200 g/L的L-精氨酸完全轉(zhuǎn)化成L-瓜氨酸。2006年,張鵬等[18]運(yùn)用海藻酸鈉包埋法固定化Streptococcus faecalisCGMCC 1866細(xì)胞生產(chǎn)L-瓜氨酸,L-精氨酸轉(zhuǎn)化率高達(dá)99%。2008年,鄭璞等[19]利用填充床反應(yīng)器固定化假單胞菌細(xì)胞連續(xù)制備L-瓜氨酸,固定化細(xì)胞對(duì)底物的摩爾轉(zhuǎn)化率在95%以上。同年,姚海峰等[20]利用Streptococcus faecalis BT001,作用于濃度為96 g/L的L-精氨酸,轉(zhuǎn)化率達(dá)到98%;2010年,趙艷杰等[21]利用海藻酸鈉固定化上述細(xì)胞,填充床反應(yīng)器中連續(xù)轉(zhuǎn)化濃度為100 g/L的L-精氨酸底物,摩爾轉(zhuǎn)化率為95.1%;2011年,屈冉[22]利用聚氨酯固定化上述細(xì)胞,填充床反應(yīng)器中轉(zhuǎn)化濃度為100 g/L的L-精氨酸底物,L-瓜氨酸的產(chǎn)量在65 g/L左右。同年,鄭雄敏等[23]利用高產(chǎn)ADI Streptococcus faecalis BM-2 CGMCC No.4990 菌株,在30 L發(fā)酵罐上小試發(fā)酵,產(chǎn)L-瓜氨酸量達(dá)到98 g/L。2012年,胡延奇[24]利用工程菌中的ADI作用于質(zhì)量濃度60 g/L的精氨酸,瓜氨酸的產(chǎn)量達(dá)55.1 g/L。
本實(shí)驗(yàn)在E.coli BL21(DE3)中成功表達(dá)了來(lái)源于Pseudomonas putida ACCC 10185的ADI,酶活力單位為26 U/mL。重點(diǎn)考察了該重組酶制備L-瓜氨酸的應(yīng)用,考察了溫度、pH、底物濃度、加酶量等對(duì)酶轉(zhuǎn)化的影響,在最優(yōu)條件下能將650 g/L的L-精氨酸鹽酸鹽完全轉(zhuǎn)化成L-瓜氨酸,均高于上述研究。下一步的研究重點(diǎn)是將該重組菌在3 L發(fā)酵罐中進(jìn)行優(yōu)化,獲得更高表達(dá)量的酶液,為工業(yè)化奠定基礎(chǔ)。同時(shí),該工藝流程簡(jiǎn)單,操作方便,這為L(zhǎng)-瓜氨酸的工業(yè)化生產(chǎn)奠定了堅(jiān)實(shí)的基礎(chǔ),具有廣闊的實(shí)際應(yīng)用前景。
4 結(jié)論
本研究成功獲得了能夠胞內(nèi)表達(dá)ADI的重組E.coli BL21(DE3)菌株,初步實(shí)驗(yàn)表明重組菌發(fā)酵液ADI酶活可達(dá)26 U/mL。采用該重組ADI優(yōu)化了酶法制備L-瓜氨酸的工藝條件。結(jié)果表明,底物L(fēng)-精氨酸鹽酸鹽濃度650 g/L,反應(yīng)pH6.0,溫度37℃,加酶量24 U/g底物,轉(zhuǎn)速100-200 r/min,轉(zhuǎn)化時(shí)間7 h,L-瓜氨酸轉(zhuǎn)化率可達(dá)100%。
[1]張克旭. 氨基酸發(fā)酵工藝學(xué)[M]. 北京:中國(guó)輕工業(yè)出版社,1992.
[2] 胡學(xué)智. 日本功能性食品研發(fā)信息[J]. 山東食品發(fā)酵, 2007(3):14-15.
[3]王晶, 趙晶. 一氧化氮的生物學(xué)效應(yīng)[J]. 中國(guó)基層醫(yī)藥,2003, 10(12):1316-1318.
[4]王冰梅. 瓜氨酸——一氧化氮循環(huán)[J]. 中國(guó)體育科技, 2005,41(5):130-131.
[5]劉娟, 路欣欣, 孟慧. 瓜氨酸的藥理作用及生產(chǎn)方法的研究進(jìn)展[J]. 藥學(xué)實(shí)踐雜志, 2011, 29(3):173-175.
[6]Lutgens L, Lambin P. Biomarkers for radiation- induced small bowel epithelial damage:An emerging role for plasma Citrulline[J]. World Journal of Gastroenterology, 2007, 13(22):3033-3042.
[7]Foyer CH, Descourvoeres P, Kunert KJ. Protect ion against oxygenradicals:an important defence mechanism studied in transgenic plants[J]. Plant Cell and Environment, 1994, 17(5):507-523.
[8]Shinji O, Yo kohama S, Masao S. Method of production citrulline by bacterial fermentation:US, 3282794[P]. 1966-07-16.
[9]Chibata I, Shi S, Kakimoto T, et al. Enzymatic preparation of L-citrulline:US, 3730838[P]. 1973-05-13.
[10]Hermes H, Croes L, Peeters W, et al. Metabolism of amino acid amides in Pseudomonas putida ATCC 12633[J]. Appl Microbiol Biotechnol, 1993, 40:519-525.
[11]Ni Y, Li Z, Sun Z. Exp ression of arginine deim inase from Pseudomonas plecoglossicida CGMCC2039 in Escherichia coli and its anti-tumor activity[J]. Curr Microbiol, 2009, 58(6):593-598.
[12]劉詠梅, 倪燁, 李娜, 等. 精氨酸脫亞胺酶基因在大腸桿菌中的克隆、表達(dá)及純化[J]. 食品與生物技術(shù)學(xué)報(bào), 2011, 30(5):750-756.
[13]錢嘉南, 孫志浩, 劉宇鵬, 等. 二乙酰一肟-氨基硫脲比色法測(cè)定酶轉(zhuǎn)化液中的L-瓜氨酸[J]. 中國(guó)醫(yī)藥工業(yè)雜志, 2007,38(7):519-521.
[14]Kakimoto T, Shibatani T, Nishimura N, et al. Enzymatic production of L-citrulline by Pseudomonas putida[J]. Applied Microbiology,1971, 22(6):992-999.
[15]Yamamoto K, Sato T, Tosa T, et al. Continuous production of L-citrulline by immobilized Pseudomonas putida cells[J]. Biotechnol Bioeng, 1974, 16(12):1589-1599.
[16]王穎. 酶法轉(zhuǎn)化制備L-瓜氨酸暨精氨酸脫亞氨基酶的性質(zhì)研究[D]. 濟(jì)南:山東大學(xué):2014.
[17] 曹瑜, 李加友, 焦慶才. 酶法轉(zhuǎn)化制備L瓜氨酸[J]. 精細(xì)化工,2005, 22(10):759-761.
[18]張鵬, 張淑榮, 劉春巧, 等. 固定化細(xì)胞連續(xù)生產(chǎn)L-瓜氨酸的方法:中國(guó), CN1948464A[P]. 2006- 11- 24.
[19]鄭璞, 倪曄, 張文. 填充床反應(yīng)器中固定化假單胞菌細(xì)胞連續(xù)制備L-瓜氨酸[J]. 食品與生物技術(shù)學(xué)報(bào), 2008, 27(5):33-38.
[20]姚海峰, 張淑榮, 歐成武, 等. 糞鏈球菌轉(zhuǎn)化合成L-瓜氨酸的研究[J]. 食品工業(yè)科技, 2008, 29(6):256-258.
[21]趙艷杰, 曾倡, 張淑榮, 等. 固定化糞鏈球菌酶法連續(xù)生產(chǎn)L-瓜氨酸[J]. 北京化工大學(xué)學(xué)報(bào):自然科學(xué)版, 2010, 37(4):98-102.
[22]屈冉. 聚氨酯固定化糞鏈球菌制備L-瓜氨酸的研究[D]. 北京:北京化工大學(xué), 2011.
[23]鄭雄敏, 王震, 劉倩高. 產(chǎn)精氨酸脫亞胺酶菌株及用它生產(chǎn)L-瓜氨酸的方法:中國(guó), CN102703339A[P]. 2011-8-29.
[24]胡延奇. L-瓜氨酸的工程菌制備及在卷煙中的應(yīng)用[J]. 食品工業(yè), 2012, 33(8):55-58.
(責(zé)任編輯 馬鑫)
Optim ization of Preparing L-citrulline by Recombinant Arginine Deim inase
Ma Yue1,2Su Lingqia1,2Wu Dan1,2Wu Jing1,2
(1. State Key Laboratory of Food Science and Technology,Jiangnan University,Wuxi214122;2. School of Biotechnology,Key Laboratory of Industrial Biotechnology of Ministry of Education,Jiangnan University,Wuxi214122)
The arcA gene encoding ADI from Pseudomonas putida ACCC 10185 was cloned into the expression vector pET-24a(+). The vector was then transformed into Escherichia coli BL21(DE3)for intracellular production of ADI. The crude enzyme was obtained by ultrasonic treatment, and activity in the fermentation broth of recombinant E. coli BL21(DE3)was 26 U/mL. Furthermore, the condition for enzymatic conversion of L-arginine monohydrochloride to L-citrulline by the recombinant ADI was optimized. At 650 g/L of L-arginine monohydrochloride, pH6.0, 37℃, 100-200 r/min, and 24 U ADI per gram substrate incubated for 7 hours, 100% of the L-arginine monohydrochloride was transformed into L-citrulline, which was the highest level of preparing L-citrulline by enzyme method in home and abroad presently.
arginine deiminase;L-citrulline;enzymatic conversion;recombinant expression
10.13560/j.cnki.biotech.bull.1985.2015.08.026
2014-12-10
馬越,女,碩士研究生,研究方向:重組酶的制備和應(yīng)用;E-mail:fionamy0912@sina.com
吳丹,男,碩士生導(dǎo)師,研究方向:食品與發(fā)酵研究;E-mail:bioenwu@aliyun.com
