山羊Apaf-1和Apaf-2基因的cDNA克隆、序列分析及組織表達
2015-10-25 07:02:33羅斌盧建遠馬力胡亮劉霜楊珂偉字向東
生物技術通報 2015年9期
羅斌 盧建遠 馬力 胡亮 劉霜 楊珂偉 字向東
(西南民族大學 國家民委動物科學重點實驗室,成都 610041)
山羊Apaf-1和Apaf-2基因的cDNA克隆、序列分析及組織表達
羅斌 盧建遠 馬力 胡亮 劉霜 楊珂偉 字向東
(西南民族大學國家民委動物科學重點實驗室,成都610041)
采集5只多胎金堂黑山羊和5只單胎藏山羊在發情期的卵巢、垂體等組織樣,進行Apaf-1和Apaf-2基因的cDNA克隆、序列分析,以及定量PCR技術對其mRNA進行組織表達量研究。結果表明,克隆出Apaf-1基因長度為3 750 bp,編碼1 249個氨基酸,Apaf-2基因編碼區全長為318 bp,編碼105個氨基酸。這兩個基因在兩種山羊中序列相同,沒有突變,且在5種組織中的表達均無差異。表明凋亡基因Apaf-1和Apaf-2在動物的進化中比較保守,與山羊多羔性狀的相關性需要進一步研究。
山羊;Apaf-1基因;Apaf-2基因;定量PCR
山羊產羔率是一個低遺傳力的數量性狀,其遺傳力只有0.1左右[1],所以用常規育種技術難以改良產羔數性狀,而產羔率主要取決于每個發情周期中母羊的排卵數。山羊在出生前,卵巢上便形成了大量原始卵泡。初情期前,卵泡雖能發育,但不能成熟排卵,當發育到一定程度時,便閉鎖(atresia)退化[2]。初情期后,在每個發情周期中可發育的卵泡多達幾十個,但單胎品種最終能發育到成熟排卵的卵泡一般只有1-2個,其余卵泡中途閉鎖死亡,而多胎品種則有多個卵泡能發育到成熟排卵[3]。藏山羊發育較慢,性成熟較晚,一年1胎[4,5],金堂黑山羊一年2-3胎[6],在分子水平上對這兩種山羊繁殖性能進行比較研究具有重要意義。
Apaf-1(Apoptotic protease activating factor-1)即凋亡蛋白酶活化因子-1和Apaf-2是體內重要的凋亡激活劑。細胞凋亡(Apoptosis)是程序性細胞死亡(Programmed cell death,PCD)的一種表現形態。1996年在細胞凋亡研究中,首次揭示出兩個與凋亡密切相關的蛋白Apaf-1和Apaf-2。而Apaf-1的cDNA是Zou等[7]在1997年克隆出來的,并證實是線蟲CED-4的人類同源物,它在哺乳動物線粒體依賴性凋亡通路和胚胎發育中至關重要,電子顯微鏡確認的Apaf-1分散于細胞質中,而不是在線粒體或其他的細胞器[8]。Apaf-1在細胞凋亡中處于核心地位,其功能涉及有細胞增殖、分化和凋亡等各個方面[9,10]。人體內一系列信號轉導級聯反應均以Apaf-1為靶而調節凋亡體。Apaf-2是細胞色素C(CytC),是一個線粒體起源的細胞凋亡信號,進行線粒體CytC信號轉導途徑[11],參與Apaf-1和Casp-9等因子結合構成凋亡體。正常狀態下CytC不能通過外膜,而在NO、過氧化物酶等因子的誘導下線粒體發生聚集,CytC便釋放到胞質中[12]。排卵前卵泡發育過程中涉及一系列的細胞增殖和凋亡事件,目前對卵泡細胞增殖機制的研究比較深入。有研究結果顯示不同綿羊、山羊品種FSH和LH的血清濃度及其受體在卵巢中的表達量與其排卵率之間無顯著相關性[13,14],而對卵泡發育過程中的細胞凋亡機制的研究則未見報道。因此,本研究對單胎山羊和多胎山羊Apaf-1和Apaf-2基因進行序列分析和組織表達分析,以期為研究山羊多胎機制提供理論依據。
1 材料與方法
1.1 材料
1.1.1 實驗動物及樣品采集 本實驗選取健康藏山羊、金堂黑山羊各5只,金堂黑山羊選自四川金堂,連續3胎產3羔山羊品種;藏山羊選自四川省理縣,連續3胎單羔的單胎山羊品種。由于藏山羊一般在秋季發情,且較晚,所以本實驗在10月份對選取的10只羊同期發情處理:將孕酮陰道栓(CIDR)放入山羊陰道內,11 d后取出,取出前一天肌肉注射氯前列醇納2 mL,在取出CIDR后40 h左右進行屠宰山羊,屠宰后采取垂體、子宮、卵巢、輸卵管和肝臟組織樣于已經標記的凍存管中,迅速投入液氮中帶回實驗室,-80℃保存備用。
1.1.2 主要酶及試劑 孕酮陰道栓(CIDR)產自新西蘭,前列腺素產自加拿大;2×Taq PCR MasterMix、動物組織總RNA提取試劑盒、DNA Marker DL2000均購自上海天根生物科技有限公司;反轉錄試劑盒購自Fermentas(MBI)公司;X-Gal、IPTG、氨芐青霉素、克隆載體pMD19-T Vector 購自大連寶TaKaRa生物工程有限公司;DNA凝膠回收試劑盒購自愛思進生物技術(杭州)有限公司;E.coli DH5α感受態細胞購自康迪生物技術有限公司,八連管、蓋子及SsoAdvancedTMSYBRòGreenSupermix購自百樂公司。
1.2 方法
1.2.1 樣品總RNA提取及反轉錄 同一時間將10只羊(金堂黑山羊和藏山羊各5只)的5種組織:子宮、卵巢、輸卵管、垂體和肝,采用Trizol法提取RNA[15];cDNA使用反轉錄試劑盒Thermo Scientific RevertAid First Strand Cdna Synthesis Kit進行合成。
1.2.2 引物設計與合成 目前羊中暫無Apafs基因序列的公布,所以參照牛的Apafs基因,根據GenBank中已公布的牛Apaf-1和Apaf-2(CYCS)基因序列(登錄號NM001191507和NM001046061),利用引物設計軟件Primer Premier5設計克隆引物和熒光定量引物,定量引物參照牛的基因和克隆出的山羊的全基因序列跨內含子設計,見表1。其中,Apaf-1采用分段克隆的策略設計簡并引物,設計3對引物分段擴增cDNA片段,引物由上海Invitrogen公司合成。
1.2.3 山羊Apafs基因的PCR擴增 以反轉錄cDNA為模板進行PCR反應,反應體系為25 μL:上下游引物(20 pmol/L)cDNA模板各1 μL,2×Taq PCR Master Mi×12.5 μL,ddH2O 9.5 μL。PCR反應條件為:95℃預變性3 min;94℃變性30 s,退火溫度Apaf-1三個片段分別是50.6℃、54.4℃和54.4℃,Apaf-2是58℃ 40 s,72℃延伸90 s,35個循環;72℃延伸5 min;4℃保存。PCR產物用1%凝膠電泳后,按照愛思進生物工程有限公司膠回收試劑盒進行回收純化。
1.2.4 克隆測序及生物信息學分析 將膠回收產物與克隆載體pMD19-T Vector于16℃連接12 h。將連接反應產物轉化宿主菌E.coli DH5α感受態細胞,涂布于含氨芐青霉素(Amp)的LB固體平板上,于37℃恒溫箱培養12 h。挑選白色單克隆菌落于含Amp的LB液體培養基37℃振蕩培養16 h,并進行菌液PCR鑒定,產物用1%瓊脂糖凝膠電泳檢測后,挑選陽性克隆菌液金堂和藏山羊的各2 mL送交上海英駿生物技術有限公司測序。

表1 克隆及定量引物序列
1.2.5 實時熒光定量PCR檢測 將提取的山羊5個組織的總mRNA,運用Real-time PCR方法檢測不同組織mRNA以及內參β-actin基因的表達量。反應體系為10 μL:上下游引物各0.8 μL、模板dDNA 0.5 μL、SYBR Green Supermix 5 μL、ddH2O 2.9 μL。PCR反應條件為:95℃預變性3 min;95℃變性10 s,61.2℃退火20 s,65℃延伸5 s,40個循環;65℃延伸5 min;4℃保存。實驗結果用SPSS17.0及Excel2007進行統計分析,用Pfaffi[16]法分析Apaf-1、Apaf-2基因在兩種山羊不同組織中的相對表達量,計算公式為:
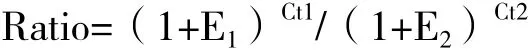
其中,E1表示目的基因的擴增效率;E2表示內參基因的擴增效率;Ct1表示對照樣本中目的基因的CT值與實驗樣本中目的基因CT值之差,Ct2表示對照樣本中內參基因的CT值與實驗樣本中內參基因的CT值之差。
2 結果
2.1 Apafs基因膠回收結果
吸取5 μL Apafs基因膠回收產物與1 μL loading buffer混勻,對其進行電泳檢測,用1%瓊脂糖凝膠電泳檢測條帶。結果(圖1)顯示,第1、2泳道與預期大小1 942 bp基本一致,第3、4泳道與預期大小1 369 bp基本一致,第5、6泳道與預期大小774 bp基本一致;第7、8泳道為金堂黑山羊和藏山羊Apaf-2基因目的條帶,與預期大小639 bp基本一致,且目的條帶較明亮,有利于后續實驗的進行。

圖1 Apaf-1和Apaf-2基因PCR產物電泳檢測
2.2 Apafs基因的核苷酸序列分析
測序結果顯示,兩種山羊的基因序列無堿基變化。將獲得的基因序列提交NCBI,Apaf-1基因全長CDS為3 750 bp,登錄號KP245916;Apaf-2基因CDS全長是318 bp,登錄號KMB823520。通過DNAman分析金堂黑山羊和藏山羊與其他物種Apaf-1和Apaf-2基因CDS區的核苷酸序列同源性,Lasergene建立同源圖,結果(圖2和圖3)顯示,Apaf-1基因CDS區的核苷酸序列與綿羊(XM_004006637)、牛(NM_001191507.1)、野豬(XM_003481742)、馬(XM_005606535)、家貓(XM_003989131)、人(AF-149794.1)、獼猴(XM_001086717.2)和褐家鼠(AF218388)的同源性分別為99.4%、97.3%、92.3%、91.5%、91.5%、86.9%、86.7%和84.2%;Apaf-2基因CDS區的核苷酸序列與綿羊(XM_004013157.1)、藏羚羊(NM_005976593.1)、牛(NM_001046061.2)、野 豬(NM_001129970.1)、 虎 鯨(XR_183516.1)、家馬(HQ889872.1)、狼(NM_001197045.1)和黃家鼠(XM_005340301.1)、小家鼠(NM_007868.4)、人類(NM_018947.5)的同源性分別為99.7%、 99.4%、98.4%、94%、93.1%、92.1%、91.2%、91.2%、90.3%和89.3%。

圖2 山羊Apaf-1基因的同源性分析
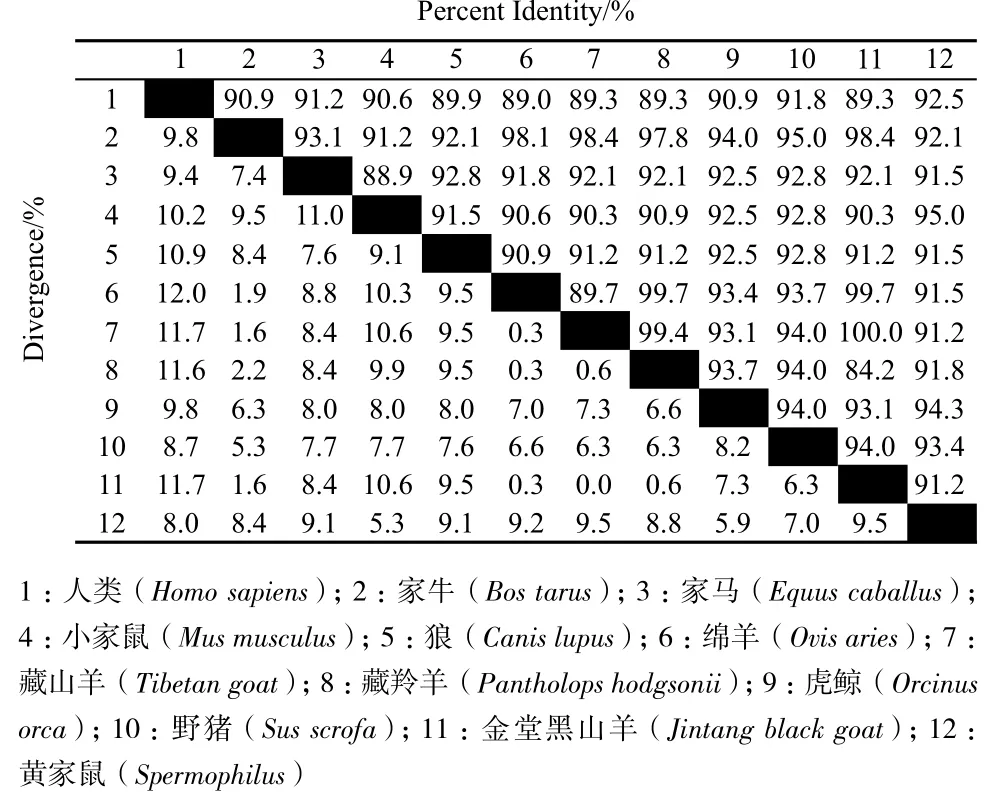
圖3 山羊Apaf-2基因的同源性分析
用MEGA6.0軟件的鄰接法(Neighbor-Joining method,NJ)構建系統進化樹,Apaf-1基因系統進化樹結果(圖4)表明,山羊和綿羊親緣關系最近,山羊先與綿羊聚為一類,再與牛聚為一類,與野豬聚為一類,然后與馬聚成一大類后,最后與人、猴、鼠為一類;Apaf-2基因系統進化樹結果(圖5)表明,山羊和綿羊親緣關系最近,山羊先與綿羊聚為一類,與牛聚為一類,然后與虎鯨、家馬、狼聚為一類,最后與人、黃家鼠和小鼠為一類,這與哺乳動物的進化程度一致。
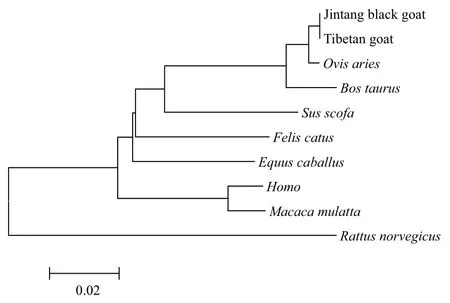
圖4 藏山羊和金堂黑山羊Apaf-1基因的系統進化樹
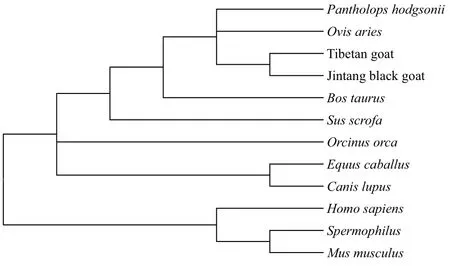
圖5 藏山羊和金堂黑山羊Apaf-2基因的系統進化樹
2.3 氨基酸序列分析
克隆獲得的山羊Apaf-1基因CDS全長3 747 bp,用ExPASy在線對其蛋白質組成進行預測發現,該基因編碼1 249個氨基酸,相對分子量為142.0 kD,等電點(p I)為5.94,其氨基酸組成中亮氨酸的含量最高,占11.2%,分子式為C6302H9874N1716O1885S67;該基因編碼的蛋白質為中性蛋白,既有親水性也有疏水性,存在于細胞質中。Apaf-2基因CDS全長315 bp,編碼105個氨基酸,相對分子量為14.5 kD,p I5.31,丙氨酸含量最高為32.7%;在第15、第80和第97位氨基酸附近,該蛋白質為疏水,其余部分為親水,共價結合于線粒體內膜。
2.4 Apafs基因定量結果的分析
采用實時熒光定量(Real-time PCR)的方法,以β-actin為參照,對Apaf-1和Apaf-2基因在藏山羊和金堂黑山羊5個組織中的mRNA相對表達量進行實驗,結果(圖6,圖7)表明,Apaf-1和Apaf-2的mRNA在山羊輸卵管、卵巢、子宮、垂體、肝中都有表達;金堂黑山羊的Apaf-1在卵巢和輸卵管中表達量較高,藏山羊的Apaf-2基因在肝臟和子宮中表達量較高,總體上Apaf-1相對表達量低于Apaf-2,但兩個品種間差異不顯著(P>0.05)。
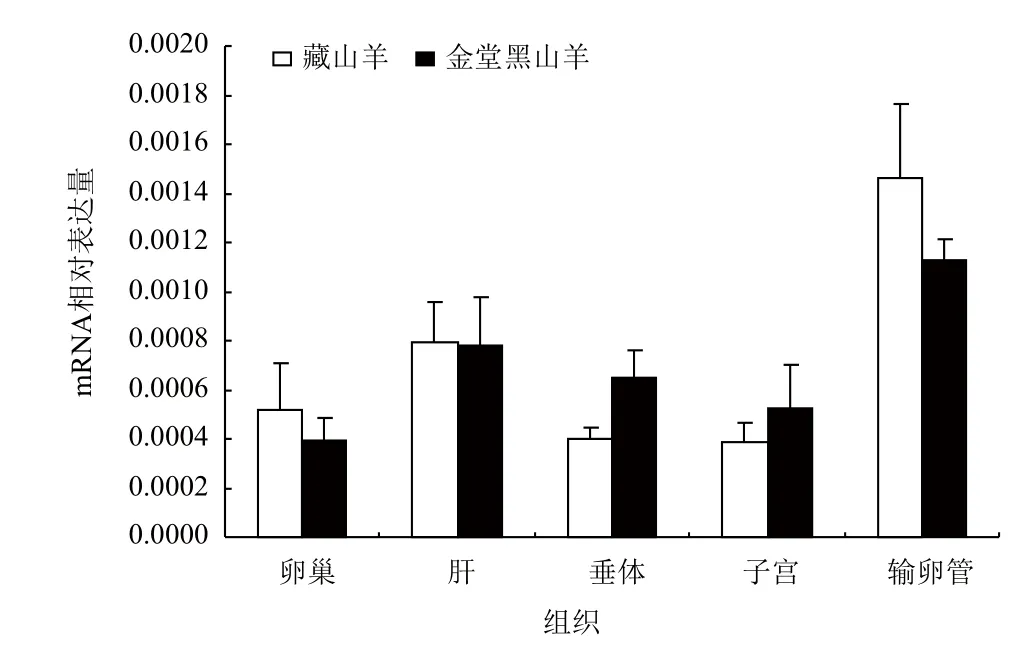
圖6 Apaf-1基因在藏山羊和金堂黑山羊不同組織中的相對表達量(x-±s)
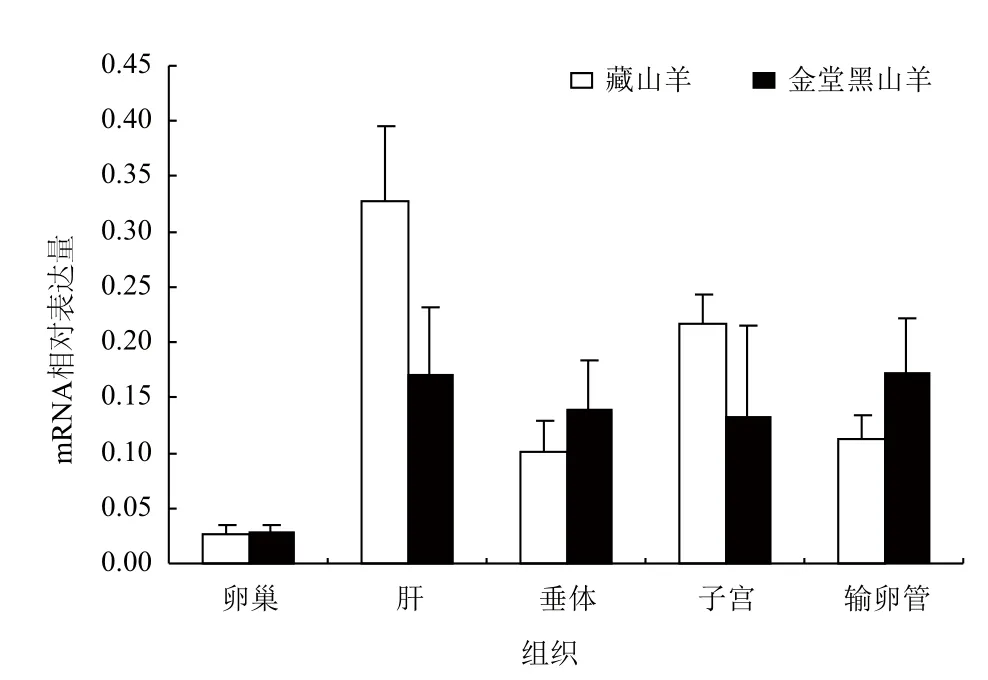
圖7 Apaf-2基因在藏山羊和金堂黑山羊不同組織中的相對表達量(x-±s)
3 討論
細胞凋亡(PCD)[17]對于多細胞生物個體發育的正常進行,自穩平衡的保持,抵御外界各種因素的干擾等起著重要的作用。近年來,對細胞凋亡的研究已從聚焦細胞核的改變,轉向對于線粒體呼吸鏈變化的研究。在細胞凋亡的研究[18]中揭示出3個與凋亡密切相關的蛋白細胞凋亡蛋白酶活化因子參與了細胞凋亡的過程,包括Apaf-1、細胞色素c(Cytc/Apaf-2)和Casp-9(Apaf-3)3個成員,參與激活caspase,通過與Bcl-2家族等蛋白因子的相互作用來調控PCD進程。Apaf-1與凋亡抑制基因是獨立的,如Bcl-2通過胱天蛋白酶非依賴性機制保護線粒體功能,從而保護細胞,但不依賴于Apaf-1[19]。而Apaf-2(細胞色素C,cytochrome C,Cytc)是呼吸鏈中的一個基本成分,在氧化還原和能量代謝中起著重要的作用,同時細胞色素C是線粒體啟動凋亡程序的關鍵物質[20]。Gabriel等[21]給細胞注射外源性Cytc后發現線粒體調節的凋亡通路可不依賴Apaf-1/Casp-9,而由線粒體膜釋放凋亡誘導因子AIF介導完成。本實驗蛋白質分析證實了Apaf-1蛋白存在于細胞質中,與Kirsten等[22]的研究結果一致,人的Apaf-1基因位于染色體12q23,Apaf-2蛋白為水溶性蛋白,位于線粒體內膜外側。
與凋亡相關的基因很多,但在哺乳動物上很少通過凋亡因子來研究對繁殖的影響。本實驗在兩種山羊不同組織中對凋亡蛋白酶活化因子Apaf-1和Apaf-2基因進行克隆、定量表達在多胎和單胎上進行對比,兩個基因核苷酸序列無差異,表達也沒有差異,與綿羊同源性達99.7%和99.4%。藏山羊和金堂黑山羊Apaf-1和Apaf-2基因編碼區序列與牛、人、小鼠、褐家鼠均有較高的同源性,說明Apaf-1和Apaf-2基因在哺乳動物中均具有較高的保守性。根據Apaf-1和Apaf-2基因編碼區核苷酸序列構建的分子系統進化樹和同源圖,山羊和綿羊親緣關系最近,其次為藏羚羊、牛,各個分支置信度高,表明系統進化樹的結果可靠。Apaf-1和Apaf-2的mRNA在研究的兩個山羊品種的卵巢、子宮、輸卵管、肝臟和垂體都有表達,但品種間無顯著差異(P>0.05),說明這兩個基因不是影響單胎和多胎的主要基因。
4 結論
本實驗采用RT-PCR技術首次克隆了山羊Apaf-1基因和Apaf-2基因,高繁金堂黑山羊與低繁藏山羊的這兩種基因CDS編碼區序列無核苷酸差異,山羊和其他物種有著很高的同源性,這兩種基因在生物進化上高度保守。Apaf-1基因包含一個3 750 bp的開放閱讀框,編碼1 249個氨基酸;Apaf-2基因CDS為318 bp編碼105個氨基酸。兩個基因在兩種山羊的表達無顯著差異。
[1] Notter DR. Genetic improvement of reproductive efficiency of sheep and goats[J]. Anim Reprod Sci, 2012, 130(3-4):147-151.
[2] Paster LM, Pallares J, Roca J. Histological characterization and in situ localization of apoptosis in the pig follicular atresia[J]. Ital J Anat Embryol, 2001, 106(2Suppl.2):257-262.
[3] Fabre S, Pierre A, Mulsant P, et al. Regulation of ovulation rate in mammals:contribution of sheep genetic models[J]. Reprod Biol Endocrinol, 2006, 4:20.
[4] 趙有璋, 張英杰, 岳文斌, 等. 羊生產學[M]. 第3版. 北京:中國農業出版社, 2011.
[5] 王杰, 王永, 歐陽熙, 等. 藏山羊研究[J]. 中國畜牧雜志,1993, 29(1):10-13.
[6] 許德貴, 張家明, 唐詩軍, 等. 金堂黑山羊生產性能研究[J].四川畜牧獸醫, 2006, 33(11):25-26.
[7] Zou H, Henzelw J, Liu X, et al. Apaf-1, a human protein homologous to c, elegans CED-4, participates in cytochrome c dependent activation of caspase-3[J]. Cell, 1997, 90(3):405-413.
[8] Hausman G, O’Reilly LAvan Driel-R, Beaumont JG. Pro-apoptotic apoptosis protease-activating factor1(Apaf-1)has a cytoplasmic localization Distinct from Bcl-2 or Bcl-x L[J]. J Cell Biol, 2000,149(3):623-633.
[9] Hoppe J, Kilic M, Hoppe V, et al. Formation of caspase-3 complexes and fragmentation of caspase-12 during an isomycin induced apoptosis inAKR-2B cells without aggregation of Apaf-1[J]. Eur Cell Biol, 2002, 81(10):567-576.
[10] Ferraro E, Corvaro M, Cecconi F. Physiological and pathological roles of Apaf1 and the apoptsome[lJ]. Cell Mol Med, 2003, 7(1):21-34.
[11] Wang X. The expanding role of mitochondria in apoptosis[J]. Genes and Development, 2001, 15(22):2922-2933.
[12] Haga N, Fujita N, Tsuruo T. Mitochondrial aggregation precedes cytochrome c release from mitochondria during apoptosis[J]. Oncogene and Oncogene Reviews, 2003, 22(36):5579-5585.
[13] Drouilhet L, Taragnat C, Fontaine J, et al. Endocrine characterization of the reproductive axis in highly prolific Lacaune sheep homozygous for the FecL(L) mutation[J]. Biol Reprod, 2010,82(2):815-824.
[14] Zi XD, Huang L, Wang Y, et al. Comparative messenger RNA expression of FSHβ, LHβ, FSHR, LHR, and ERβ in high and low prolific goat breeds[J]. Anim Biotechnol, 2013, 24(4):307-311.
[15] 薩姆布魯克J, 拉塞爾DW. 分子克隆實驗指南[M]. 北京:科學出版社, 2002.
[16] Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR[J]. Nucleic Acids Res, 2001, 29(9):e45.
[17] Kerr JF, Wyllie AH, Currie AR. Apoptosis:a basic biological phenomenon with wide ranging implications in tissue kinctics[J]. Cancer, 1972, 26:239-257.
[18] Li P, Nijhawan D, Budihardjo I, et al. Cytochrome c and dATP-dependent formation of Apaf-1/Caspase-9 complex initiates an apoptotic protease cascade[J]. Cell, 2004, 116:479-489.
[19] Haraguchia M, Torii S. Apoptotic protease activating factor 1(Apaf-1)-Independent Cell death suppression by Bcl-2[J]. J Exp Med,2000, 191(10):1709-1720.
[20] 吳蘭芳, 楊愛珍, 劉和, 等. 線粒體調控細胞凋亡的研究進展[J]. 中國農學通報, 2010, 26(8):63-68.
[21] Gabriel B, Sureau F, Casselyn M, et al. Retroactive pathway involving mitochondria in electroloaded cytochrome c-induced apoptosis:Protective properties of Bcl-2 and Bcl-XL[J]. Exp Cell Res, 2003, 289(2):195-210.
[22] Kirsten L, Helga AE, Stephan FS, et al The adapter protein apoptotic protease-activating factor-1(Apaf-1)is proteolytically processed during apoptosis[J]. J Biol Chem, 2001, 276(32):29772 -29781.
(責任編輯 馬鑫)
cDNA Cloning,Sequence Analysis and Tissue Expression of Apaf-1 and Apaf-2 Gene of Goats
Luo Bin Lu Jianyuan Ma Li Hu Liang Liu Shuang Yang Kewei Zi Xiangdong
(The Key Laboratory of Animal Science of State Ethnic,Affairs Commission,Southwest University for Nationalities,Chengdu610041)
We collected the pituitary, ovary and other tissue samples of 5 Tibetan goats and 5 Jintang black goats at estrus stage, cloned cDNA of Apaf-1 and Apaf-2, analyzed their sequences, and investigated their mRNA expression of tissues by qPCR. The results showed that:the coding region sequence(CDS)of goat Apaf-1 gene was 3 750 bp long encoding 1 249 amino acids, and the CDS of Apaf-2 gene was 318 bp encoding 105 amino acids. The two genes showed the same sequences in both of goats, no mutation and no differential expression in 5 tissues. Our data suggested that gene Apaf-1 and Apaf-2 in animal’s evolution were conservative, and the correlation with the trait of goat’s prolificacy needs further studies.
goat;Apaf-1 gene;Apaf-2 gene;qPCR
10.13560/j.cnki.biotech.bull.1985.2015.09.017
2014-12-21
西南民族大學研究生創新型科研項目(CX2014SZ71)
羅斌,男,碩士研究生,研究方向:動物遺傳育種與繁殖;E-mail:romben@sina.com
字向東,碩士,教授,研究方向:動物遺傳育種與繁殖;E-mail:zixd@sina.com
