若爾蓋濕地公路建設(shè)生態(tài)恢復(fù)中植物地上生物量研究
2015-10-28 21:10:37陳蓓馬睿秦紀洪等
湖北農(nóng)業(yè)科學(xué) 2015年19期
陳蓓 馬睿 秦紀洪等
摘要:在若爾蓋濕地段公路建設(shè)區(qū),草地生態(tài)系統(tǒng)恢復(fù)和重建采取了草皮切割貯存并在主體工程完工后將草皮回覆創(chuàng)面進行生態(tài)快速恢復(fù)的模式,以期實現(xiàn)有效保持水土、構(gòu)建生態(tài)公路的目標。對恢復(fù)群落和原生背景群落的地上生物量進行研究,以評價這種生態(tài)恢復(fù)技術(shù)的可行性與有效性。通過對群落種類組成基本性質(zhì)以及群落生物量與組成植物群落中的種類密度分析,結(jié)果表明,優(yōu)勢種對群落性質(zhì)和環(huán)境具有控制性影響,切割草皮與背景樣方相比,植物種類有所減少,但優(yōu)勢種組成基本未發(fā)生變化;對地上生物量結(jié)構(gòu)波動與變化規(guī)律研究結(jié)果表明,生物量隨季節(jié)的變化而變化,在8月中下旬分別達到它們的峰值生物量;對群落的結(jié)構(gòu)分析表明,切割樣地有其特定的群落結(jié)構(gòu)和功能。
關(guān)鍵詞:若爾蓋濕地;公路建設(shè)區(qū);人工恢復(fù)植被;地上生物量
中圖分類號:S157;X171.4 文獻標志碼:A 文章編號:0439-8114(2015)19-4676-06
DOI:10.14088/j.cnki.issn0439-8114.2015.19.006
Abstract:Grassland ecosystem restoration and reconstruction of highway construction in the Zoige wetland was studied combining artificial turf cutting technology with relocating technology. In this study, the aboveground biomass of the restoration community and the native community was determined to evaluate the feasibility and effectiveness of this technique. By analyzing the species composition of the community and the species density in the plant community, the results showed that the dominant species had a controlling influence on the community nature and the environment. The plant species of turf cutting plots decreased when compared with the control, while the composition of the dominant species were not changeable in turf cutting plots and the control. The results of the research on the fluctuation and variation of the aboveground biomass structure showed that the biomass varied with the seasons. The peak biomass was reached in late August. The structure analysis of the community showed that the cutting plots had specific community structure and the function.
Key words:Zoige Wetland; highway construction zone; ecological restoration; aboveground biomass
國道213線郎川段(郎木寺—川主寺)改擴建工程從若爾蓋國家級濕地保護區(qū)以及若爾蓋大草原通過,全線3/4路段位于海拔3 200~3 800 m的高寒地區(qū),有著種群豐富且保護較好的高寒自然生態(tài)系統(tǒng)。鑒于若爾蓋高寒草甸草原和高原濕地地處青藏高原東南緣的生態(tài)脆弱區(qū)的環(huán)境敏感性,以及作為長江與黃河上游重要水源地、高原濕地生物多樣性重點區(qū)域的生態(tài)重要性,在設(shè)計與建設(shè)時充分考慮了有效的生態(tài)與工程措施以降低工程對草原與濕地生境的破壞。高寒草甸草粘層形成時間漫長,一旦破壞不但植被難以恢復(fù),而且草粘層喪失會直接導(dǎo)致高寒草甸土壤侵蝕與沙化,因此在工程建設(shè)時采用了工程前將施工地表的草皮切塊堆放貯藏,主體施工完成后將切塊堆放的草皮覆蓋工程中破壞的道路兩側(cè)地表,從而實現(xiàn)地表植物群落快速恢復(fù)和有效保持水土、構(gòu)建生態(tài)公路的目標。這些措施的實施及其效果評價,對于高寒區(qū)域工程施工后的生態(tài)恢復(fù)重建與水土保持具有重要意義。鑒于若爾蓋高寒草甸草原和高原濕地的生態(tài)脆弱性和生態(tài)重要性,如何在公路建設(shè)時減小對脆弱的草原、濕地生境的破壞,同時對工程中破壞的植被進行快速恢復(fù)和有效保持水土,是郎川公路改擴建工程急需解決的問題之一。
前期的初步試驗中表明,通過收集開挖地面的草皮,回覆移植加強濕地和草原創(chuàng)面的生態(tài)恢復(fù)是可行的[1,2],在川西北高原濕地的工程建設(shè)中采取草皮移植快速恢復(fù)工程破壞生境的措施在一定程度上取得了成效,前期的研究表明草皮恢復(fù)后的植物群落結(jié)構(gòu)穩(wěn)定且成效與割取草皮厚度、面積大小與面積相關(guān)[3],對恢復(fù)群落和原生背景群落的相似性指數(shù)(Jacard指數(shù)和Sorenson指數(shù))開展對比研究發(fā)現(xiàn)[4]:取樣面積越大、與背景越相似的群落,恢復(fù)效果越好;對β多樣性指數(shù)進行研究表明,β多樣性測度指標以物種的個體數(shù)作為測度指標將導(dǎo)致較大的誤差,而以相對蓋度、重要值作為測度指標時則得到較合理的結(jié)果。當然,全面評估這種生態(tài)恢復(fù)技術(shù),還需要對恢復(fù)群落和原生背景群落的地上及地下生物量進行研究[5],地上生物量是初級生產(chǎn)力的重要組成部分和表現(xiàn)形式,本研究將初級生產(chǎn)力作為一種評價生態(tài)功能的指標,以評價在高寒地帶工程建設(shè)中通過利用原生草皮剝離后再覆蓋進行生態(tài)恢復(fù)技術(shù)的可行性與有效性。
1 材料與方法
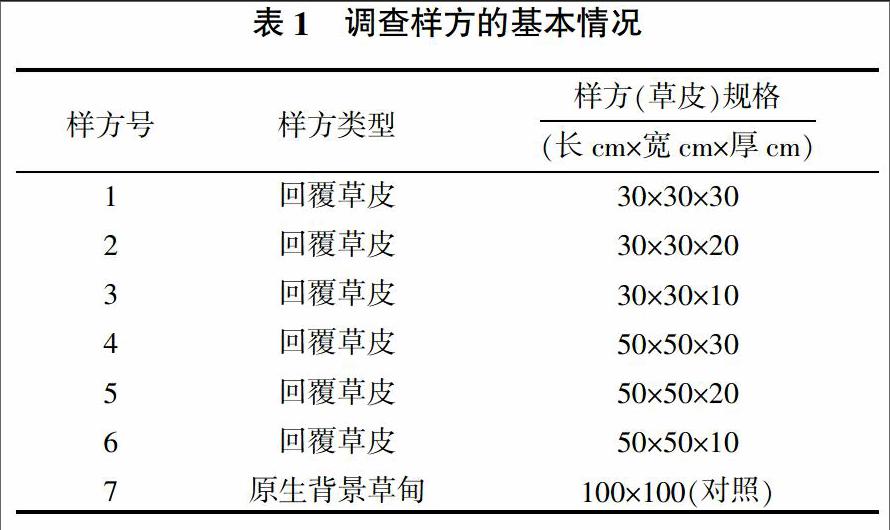
1.1 國道213郎木寺—川主寺段概況
試驗點地位于郎川路線中北段,屬若爾蓋縣,32°11′N-33°56′N,102°35′E-103°03′E,平均海拔2 900~3 850 m,具青藏高原大陸季風(fēng)氣候特征。冬季(11月至次年4月)主要受西北風(fēng)冷氣團控制,寒冷干燥、日照多而強、日溫差大。夏季(5~10月)受西南季風(fēng)控制,降水明顯增多而進入濕季。多年平均氣溫1.1 ℃,最熱月(7月)多年平均氣溫10.7 ℃,最冷月(1月)多年平均氣溫-10.7 ℃;極端最高氣溫24.6 ℃,極端最低氣溫-33.7 ℃。年平均降雨量656.8 mm,最大年降水量862.9 mm,歷年單日最大雨強65.3 mm/d,歷年平均蒸發(fā)量1 233.2 mm,歷年平均相對濕度78%。草地年均日照實際時數(shù)2 417 h,年平均風(fēng)速2.4 m/s。
1.2 樣地選擇與調(diào)查方法
野外調(diào)查地點為郎川公路若爾蓋花湖地區(qū)草皮移栽試驗樣地,離公路50 m,樣方規(guī)格如表1。選取不同切割規(guī)格(面積和厚度)的草皮回覆道路建設(shè)破壞的地表后群落恢復(fù)情況的樣方6個,背景原生草甸樣方1個,分別進行物種調(diào)查、密度、高度、蓋度、頻度的測定,在公路兩側(cè)樣地(面積為100 cm×100 cm 隨機取各植物地上部分50株(個別種為100株),將莎草科和禾草科植物的每一分蘗枝作為一株,測定其生物量(烘干重),并計算出各植物種群平均單株生物量。地上生物量與地下生物量測定同步進行,齊地剪取,樣方面積50 cm×50 cm,3次重復(fù)。地上凈生產(chǎn)量按植物枯黃前(8月底)測定得最高生物量為依據(jù)。全部生物量均以烘干記重。
1.3 物種重要值指標
重要值計算公式為:IV=(相對高度+相對蓋度+相對密度+相對頻度+相對生物量)/5。
2 結(jié)果與分析
2.1 樣地群落種類組成的基本性質(zhì)
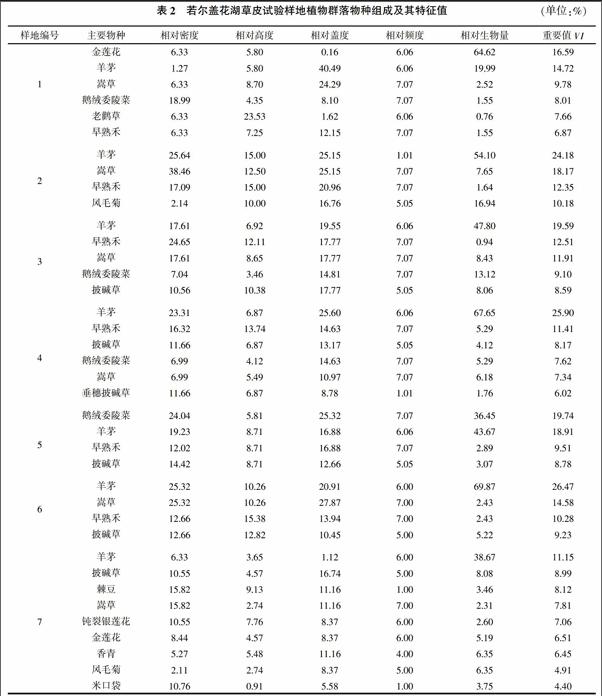
在樣地植物群落研究中,對物種進行逐個登記,主要包括物種調(diào)查、密度、高度、蓋度、頻度、地上生物量(鮮重和干重)的測定,得到一份所研究群落的植物種類名錄。群落中的主要植物種群有:羊茅(Festuca ovina Linn)、早熟禾(Poa pratensis)、垂穗披堿草(Elymus nutans)、嵩草(Kobresia)、老鸛草(Geranium pylzowianum)、風(fēng)毛菊(Saussurea supera)、金蓮花(Trollius buddae)、棘豆(Oxytropis sp.)、鈍裂銀蓮花(Anemone sp.)、鵝絨委陵菜(Potentilla anserina)、毛茛(Ranunculus japonica)、薹草(Carex sp.)、川藏蒲公英(Taraxacum maurocarpum)、馬先蒿(Pedicularis kansuensis)等。
表2列出了每個樣地中重要值大于該組平均值的物種及其特征值。調(diào)查發(fā)現(xiàn),該區(qū)草皮的主要植物有32種,隸屬8科28屬,在若爾蓋花湖草皮試驗樣地中,植物群落多以耐寒多年生禾本科和莎草科為主,人工移栽草皮上的植物群落除禾本科外,蓼科、菊科和毛茛科、豆科等類群的物種增加,群落的優(yōu)勢種和建群種以羊茅、早熟禾、披堿草和嵩草為主。
本研究樣地中優(yōu)勢種以羊茅為主,它們個體數(shù)量多、蓋度大、生物量高、生活能力強,即優(yōu)勢度大。優(yōu)勢種對群落性質(zhì)和環(huán)境具有控制性影響,如果除去了優(yōu)勢種,群落將發(fā)生根本的變化,并影響其他物種生存。亞優(yōu)勢的個體數(shù)量和作用都僅次于優(yōu)勢種,但是在決定群落性質(zhì)和控制群落環(huán)境方面仍起著一定作用的植物種。本研究樣地亞優(yōu)勢種主要有披堿草、嵩草、早熟禾、金蓮花等。伴生種與優(yōu)勢種相伴存在,但不起主要作用。樣地中常見的有鵝絨委陵菜、棘豆、車前草、鈍裂銀蓮花、香青、高原毛茛等,它們在群落中數(shù)量較少,是常見的伴生種。偶見種是指那些在群落中出現(xiàn)頻率很低的種類,多半是由于種群本身數(shù)量稀少的緣故。樣地中出現(xiàn)的偶見種有狼毒(Stellerachamae jasme)、黃帚囊吾(Ligularia virgaurea)、龍膽(Gentiana spp.)、金露梅(Potentilla fruticosa)等。偶見種可能是偶然的入侵種,也可能是群落中的衰退殘遺種。有的偶見種對生態(tài)環(huán)境的變化具有一定的指示意義。
羊茅在6個試驗樣地中重要值最大(其平均值為20.13%),為該群落的主要組成成分;次優(yōu)勢種依次是嵩草、早熟禾、披堿草、鵝絨委陵菜、老鸛草、金蓮花、垂穗披堿草、棘豆、鈍裂銀蓮花、香青(Anaphalis sp.)等;其余21種植物的重要值均小于種的平均值(7.15%),在整個群落組成中占次要地位,而且多為植株矮小、有毒有害的雜草類,如狼毒。7#樣地作為背景樣方具有最多的植物種類(23種),優(yōu)勢種是羊茅和披堿草,均為優(yōu)良牧草;次優(yōu)勢種為棘豆、嵩草、鈍裂銀蓮花、金蓮花等。經(jīng)過切割移栽的草皮與背景樣方相比植物種類有所減少,除禾本科和莎草科以外的植物及一年生植物增加,雜草增加,但優(yōu)勢種組成基本未發(fā)生變化。
對物種的特征值分析表明,羊茅在6個試驗樣地的相對密度為1.27%~25.64%,與背景樣方(6.33%)相比大部分值均偏高;相對蓋度變化范圍為19.55%~40.49%,明顯高于背景樣方的1.12%;相對生物量變化范圍為36.45%~69.87%,也高于背景樣方的38.67%。其他的主要優(yōu)勢種也具有類似的變化,說明切割的草皮仍然以優(yōu)良的牧草為主,但切割草皮上優(yōu)勢種的基本指標均高于背景樣方。此外,一些偶見種也出現(xiàn)在試驗樣地中,如鵝絨委陵菜,在背景樣方為偶見種,但在各個草皮試驗樣方內(nèi)則變成常見種。這可能由于切割的草皮具有邊緣效應(yīng),草皮邊緣的植物長勢會好于大片的原生植被。
2.2 群落生物量與組成植物群落中的種類密度分析
群落生物量直接與組成植物群落中的種類密度相關(guān),植物密度關(guān)系到植物進行光合作用葉面積的大小。樣地中嵩草和羊茅的密度最高,其次是垂穗披堿草、早熟禾等。這些植物多成叢生長,數(shù)量雖多,除垂穗披堿草植株較高大外,其余多生長較低矮。薹草、棘豆、馬先蒿的植株密度較低,但它們植叢較明顯,而且在群落中分布不太均勻。移栽草皮群落與原生群落物種特征值有差異,區(qū)內(nèi)植物群落優(yōu)勢種和建群種以羊茅、披堿草和嵩草為主(表2),羊茅為該群落的主要優(yōu)勢種,其他植物在整個群落組成中占次要地位,且多為有毒有害的雜草類。
切割移栽的草皮與背景樣方相比植物種類有所減少,除禾本科和莎草科以外的植物及一年生植物和雜草增加,但優(yōu)勢種組成基本未發(fā)生變化。草皮樣地除禾本科和莎草科以外的植物和一年生植物增加,表明人為干擾的植物群落在結(jié)構(gòu)與功能方面發(fā)生了退化。各物種的特征值與背景樣方相比大部分值均偏高,表明切割草皮仍然以優(yōu)良的牧草為主。另外,樣地形狀和邊界與多樣性密切相關(guān),長方形樣地(即樣帶或樣條)優(yōu)于面積相等的正方形樣地,長方形樣地可能會反映更多的變異情況。因此,還需進一步對不同形狀的樣地進行比較研究,確定切割草皮的形狀、面積或周長對草皮移栽后的恢復(fù)狀況的影響。
2.3 地上生物量結(jié)構(gòu)波動與變化規(guī)律研究
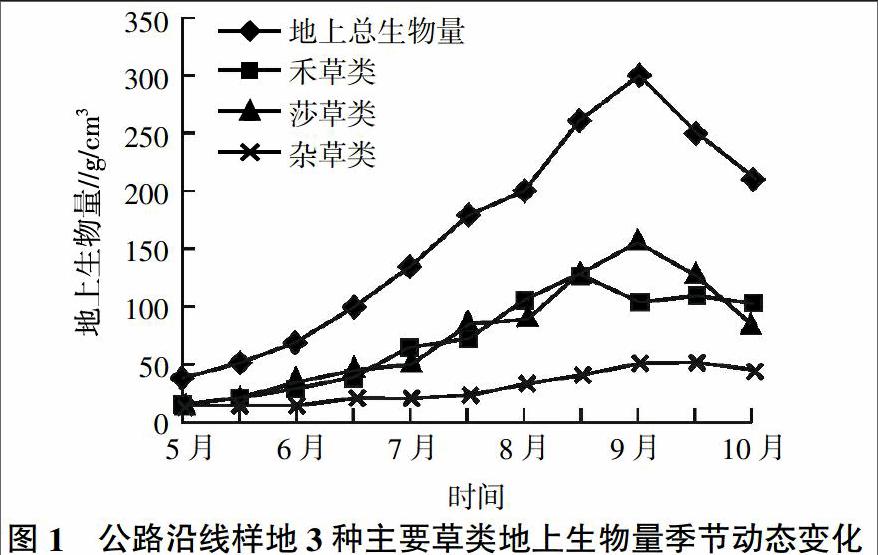
為了便于地上生物量的測定,將組成草甸的植物分為3大類:禾草類、莎草類和雜草類。禾草類主要包括禾本科的羊茅、早熟禾、洽草、垂穗披堿草等;莎草類包括嵩草、薹草等;除上述兩類外,其他所有雙子葉植物均歸于雜草類。
禾草類、莎草類、雜草類生物量隨季節(jié)的變化而變化(圖1)。總體來講,在8月中下旬分別達到它們的峰值生物量。禾草類先于其他草類達到峰值,而莎草類、雜草類及其地上生物總量均在9月中旬達到峰值。從上述現(xiàn)象分析,地上生物量峰值出現(xiàn)在7月底至8月上旬之后。不同植物類群,因它們的生態(tài)-生物學(xué)特性各異,禾草類在8月中下旬普遍果實成熟而進入枯黃期;雜草類結(jié)實較晚,莎草類枯黃較遲,故峰值生物量出現(xiàn)較禾草類晚,峰值生物量出現(xiàn)后,因氣溫降低,生物量逐漸下降,直到枯黃時停止。
地上生物量的累積動態(tài),還可以從5~9月生物量的生長強度來說明(表3)。生長強度是指在某一生長期內(nèi),平均單位時間內(nèi)生物量的生長率。從表3可以看出,在相同生長期內(nèi),不同的草類生長強度不一,如在剛返青后的4月25日至5月30日期間,禾草類的生長強度[0.538 9 g/(m2·d)]高于雜草類[0.444 4 g/(m2·d)]和莎草類[0.405 6 g/(m2·d)]。表3還說明,同一類植物在不同生長期生長強度不一,如禾草類,在4月25日至5月30日返青期,生長強度為[0.538 9 g/(m2·d)],到5月31日至6月15日拔節(jié)期間為[1.067 5 g/(m2·d)],而在7月16至7月30日孕穗期和8月16日至30日成熟期間,生長強度達到最大,分別為[2.231 3 g/(m2·d)]和[2.410 0 g/(m2·d)]。各類草生物量所占總生物量的百分比也隨著生長季節(jié)而變化。禾草、莎草類因返青較早,它們分別占總生物量的38.8%和29.2%,雜草占32%。隨著禾草類、雜草類生長發(fā)育,它們的比例不斷上升,而莎草類則相應(yīng)下降;枯黃前,禾草類為52.24%,莎草為14.16%,而雜草為33.6%。枯黃前禾草比例較高,因為禾草類植株較高(一般為30~40 cm),從拔節(jié)、孕穗、抽穗到子粒成熟,生物量相應(yīng)增加較快。雜草類植株較少,但植株平均個體單枝重量較高,特別是到了7月中旬以后,一些雜草類相繼開花,植株生長旺盛,生物量增加較快。莎草類植物生長稠密而低矮,優(yōu)勢種嵩草高5~8 cm,屬于寒冷中生和冷旱中生植物,返青早,6月中旬已進入盛花期;生長早期,生物量較高,后來隨著禾草類、雜草類迅速生長,相比之下,它們占總生物量的比率相應(yīng)下降。
不同切割草皮樣地間的地上生物量差異見表4,背景樣方為34.65 g/cm3,在切割草皮中樣地1地上生物量最大,達51.53 g/cm3,其次是樣地4、樣地5樣地6,分別為34.00、27.71、25.58 g/cm3;而樣地3、樣地2的地上生物量最小,分別為18.3 g/cm3和10.67 g/cm3。造成這種差異的主要原因是不同樣地植物種類、年齡結(jié)構(gòu)、密度、環(huán)境因子及人為活動干擾的不同。
2.4 群落的結(jié)構(gòu)分析
2.4.1 垂直結(jié)構(gòu) 草地群落的垂直結(jié)構(gòu)主要指群落的分層現(xiàn)象,也稱為群落的成層性。草地有其特定的群落結(jié)構(gòu),比較簡單,并與其功能相聯(lián)系。
第一層高20~10 cm,由紫羊茅、嵩草、高原毛茛、圓穗蓼、珠芽蓼、龍膽科、馬先蒿、高原紫菀、花錨、鵝鸛草、草玉梅、莎草、發(fā)草等組成。
第二層高10~5 cm,由垂穗披堿草、早熟禾、金蓮花、鈍裂銀蓮花、棘豆、蒲公英、黃帚橐吾、狼毒、車前草、老鸛草等組成。
第三層高5 cm以下,由鵝絨委陵菜、風(fēng)毛菊、香青等組成。
群落的垂直結(jié)構(gòu)不僅表現(xiàn)在地上部分,地下的根系也有明顯的成層性,不同種類的根系可分布在不同的土層深度。但是,最大根量分布在土壤的表層,這與土壤養(yǎng)分的分布是一致的。在本研究樣地中,表層土壤有大量草根且盤結(jié)緊實形成草氈層,而10~20 cm土壤中有少部分疏松的根系,20~30 cm土壤中則只有很少量根系。
群落的垂直結(jié)構(gòu)具有提高有限資源的時空利用范圍和利用效率的意義,因此有利于提高群落的生產(chǎn)力水平;減少物種為爭奪養(yǎng)分和空間資源的競爭,使群落擁有更高的的物種多樣性;成層性提高了生物群落的穩(wěn)定性和對環(huán)境的改造作用,增強了抵御外界干擾的能力;成層性的復(fù)雜程度是生態(tài)環(huán)境的一種良好指示,一般層次越復(fù)雜,極端環(huán)境中的生物群落越簡單。
2.4.2 水平結(jié)構(gòu) 群落的水平結(jié)構(gòu)是指群落的水平配置狀況或水平格局。本研究中的草皮形成小的群落,生長發(fā)育好,有時還可以生長一些越帶分布的植物,并形成一定數(shù)量的斑塊。每一個斑塊是一個小群落,它們彼此組合形成群落的鑲嵌性水平結(jié)構(gòu)。自然界中群落的鑲嵌性是絕對的,而均勻性是相對的。
2.4.3 時間結(jié)構(gòu) 群落的外貌常隨著時間的推移而發(fā)生周期性的變化,這是群落結(jié)構(gòu)的另一個重要特征,即時間結(jié)構(gòu)。在研究區(qū)域草原群落中,一般在5~8月水熱充沛,植物繁茂生長,百花盛開,色彩豐富;秋末植物開始干枯休眠,呈紅黃相間的秋季季相。冬季則是一片枯黃,或被白雪覆蓋。
3 小結(jié)與討論
地上生物量是初級生產(chǎn)力的重要組成部分和表現(xiàn)形式,本研究我們將初級生產(chǎn)力作為一種評價生態(tài)功能的指標。Pfistere等[6]利用植物地上生物量來度量群落生產(chǎn)力,地上生物量是總生物量的一部分,但它所占的比例不是一成不變的,因為在不同環(huán)境中植物會有不同的資源分配對策。對人工恢復(fù)高寒草甸的植物群落多樣性與地上生物量的研究表明,物種豐富度依次增加,地上生物量依次降低。植株的大小關(guān)系到物種對水分、光照的競爭能力,和對其他種的生存能否起到抑制作用。嵩草、羊茅、垂穗披堿草、早熟禾等這些植物多成叢生長、數(shù)量多,除垂穗披堿草植株較高大外,其余多生長較低矮。
在高寒環(huán)境中,水分成為限制性因素,根系競爭會增強[7],植物分配在根系生長的資源所占比例上升。所以在受到擾動時,地上生物量占總生物量的比例下降,這種下降趨勢在物種豐富的群落中更明顯,是因為這些群落有更強烈的種間根系競爭。所以對于不同物種多樣性的群落,地上生物量占總生物量的比例不同。從地上生物量變化得出的結(jié)果無法反映總生物量的變化。在這項研究中,物種豐富的群落在受到草皮切割擾動時,地上生物量占總生物量比例可能會低于物種少的群落,前者生產(chǎn)力的降低值被高估了。
隨物種多樣性上升,群落生產(chǎn)力呈升高趨勢。Tilman等[8]綜合該項試驗多年的數(shù)據(jù)發(fā)現(xiàn):生產(chǎn)力隨多樣性上升而升高,多樣性的這種正效應(yīng)逐年增強;混合群落發(fā)生超產(chǎn)的比例以及超產(chǎn)的強度逐年提高,這表明生態(tài)位互補的作用隨時間推移而強化。
參考文獻:
[1] 陳 蓓,孫 輝,秦紀洪,等.朗川公路若爾蓋濕地段人工移栽草皮的群落生態(tài)學(xué)研究[J].水土保持研究,2007,14(2):68-71.
[2] 陳 蓓,孫 輝,秦紀洪,等.高寒地區(qū)道路工程的生態(tài)效應(yīng)及其生態(tài)恢復(fù)研究進展[J].四川環(huán)境,2009,28(2):102-105.
[3] 馬 睿,孫 輝,陳 蓓,等.若爾蓋郎川公路建設(shè)區(qū)有毒有害植物擴散及生態(tài)安全性評價[J].世界科技研究與發(fā)展,2006,28(3):70-75.
[4] 陳 蓓,馬 睿,秦紀洪,等.若爾蓋濕地公路建設(shè)生態(tài)恢復(fù)中植物群落多樣性與相似性研究[J].水土保持研究,2013,20(4):115-119.
[5] 楊兆平,歐陽華,宋明華,等.青藏高原多年凍土區(qū)高寒植被物種多樣性和地上生物量[J].生態(tài)學(xué)雜志,2010,29(4):617-623.
[6] PFISTERER A B,SCHMID B. Diversity-dependent production can decrease the stability of ecosystem functioning[J]. Nature, 2002,416:84-86.
[7] WILSON MV, SCHMIDA A. Measuring beta diversity with presence-absence data[J]. J Ecol, 1984,72:1055-1064.
[8] TILMAN D. Functional diversity[A]. Encyclopedia of Biodiversity (Vol.3)[C]. New York: Academic Press, 2001.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
保健醫(yī)苑(2021年7期)2021-08-13 08:48:02
學(xué)生天地(2020年36期)2020-06-09 03:12:30
小學(xué)科學(xué)(學(xué)生版)(2020年5期)2020-05-25 07:11:32
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·少年版(2015年1期)2015-07-07 17:15:12
