不同發酵特性的嗜熱鏈球菌與保加利亞乳桿菌共發酵的特性
2015-11-02 13:00:46馬成杰徐志平徐愛才華寶珍
食品科學 2015年15期
李 莎,馬成杰*,徐志平,徐愛才,華寶珍
(乳業生物技術國家重點實驗室,光明乳業股份有限公司乳業研究院,上海 200436)
不同發酵特性的嗜熱鏈球菌與保加利亞乳桿菌共發酵的特性
李 莎,馬成杰*,徐志平,徐愛才,華寶珍
(乳業生物技術國家重點實驗室,光明乳業股份有限公司乳業研究院,上海 200436)
為探究具有不同優良性狀的嗜熱鏈球菌株與保加利亞乳桿菌株共發酵的特性,用產酸快的嗜熱鏈球菌StCH-1菌株和產黏好的嗜熱鏈球菌StTA040菌株與保加利亞乳桿菌Lb0925B菌株組合,測定其組合菌株的發酵性能。通過實驗發現組合菌株發酵速率均比單菌株發酵速率快,其中含有嗜熱鏈球菌StCH-1的組合菌株發酵速率較快,而含有嗜熱鏈球菌StTA040的組合菌株的胞外多糖產量較高,發酵乳的黏度較高,持水力較強;三組分發酵劑的發酵速率快,發酵乳在貯藏期間黏度高,持水力強,通過感官分析得出三組分發酵劑制得的發酵乳的口感和風味最佳。
嗜熱鏈球菌;胞外多糖;共發酵;發酵特性;感官評價
酸奶發酵劑一般由嗜熱鏈球菌(Streptococcus thermophilus)和保加利亞乳桿菌(Lactobacillus delbrueckii subsp. bulgaricus)組成,其中嗜熱鏈球菌在酸奶發酵前期產酸、產多糖等方面發揮著重要作用[1-3]。產酸能力強的嗜熱鏈球菌有助于酸奶產品快速發酵凝乳、縮短發酵時間、提高生產效率、降低酸奶生產成本[4]。嗜熱鏈球菌分泌的胞外多糖對發酵乳質構特性影響最大,乳酸菌胞外多糖不僅可賦予發酵乳特殊的風味,而且賦予發酵乳細膩爽滑的口感,提高發酵乳的穩定性和 持水性,減少發酵乳在生產中穩定劑的使用量[5-9]。Ruas-Madiedo等[10]認為乳酸菌分泌的胞外多糖在發酵乳制品中起到了非常重要的作用,是一種天然生物增稠劑。
然而,單一的嗜熱鏈球菌株難以具備產酸快、產黏好、抗噬菌體、風味濃郁等優良性狀。因此,將優良菌株進行組合有利于提升發酵劑的整體性能。Skriver等[4]最早提出將不同發酵性能的乳酸菌進行組合以發揮不同優良菌株的發酵性能。Urshev等[11]認為酸奶發酵劑的發展主要依賴于將一種保加利亞乳桿菌與不同嗜熱鏈球菌進行組合,從而使得發酵劑同時具備發酵速率快、產品黏度好、特征風味濃郁等優良性能。
目前,國內外酸 奶發酵劑的研究主要側重于優良菌株的選育[12-14]、菌株生產生化性能研究[15-18]、菌株高密度培養與發酵優化[19-20]、發酵劑冷凍干燥技術[21-22]等方面,少有關于具有不同優良性狀的嗜熱鏈球菌株組合的研究報道。本實驗將產酸快和產黏好的2 株嗜熱鏈球菌與1 株保加利亞乳桿菌組合,測定其組合菌株的發酵性能和對發酵乳貯藏穩定性的影響,并且通過感官評價測定不同組合菌株對發酵乳口感和風味的影響,以期為酸奶發酵劑的優化設計提供理論基礎。
1 材料與方法
1.1菌株
嗜熱鏈球菌StCH-1 丹麥Chr. Hansen公司,嗜熱鏈球菌StTA040 丹麥Danisco公司,是從酸奶發酵劑中通過選擇性LM17培養基分離,《伯杰氏細菌鑒定手冊》方法鑒定;保加利亞乳桿菌Lb 0925B 丹麥Danisco公司,是從發酵劑中通過選擇性MRS培養基分離,《伯杰氏細菌鑒定手冊》方法鑒定。
1.2材料與培養基
脫脂乳粉 新西蘭恒天然公司。
MRS(de Man, Rogosa and Sharpe)培養基、M17培養基(添加0.5%乳糖) 英國Oxoid公司。
1.3儀器與設備
Cinac發酵素酸化監控系統 法國Alliance公司;MIR-253生化培養箱 日本Sanyo公司;Bactron-1厭氧培養箱 美國Shellab公司;GFL1002恒溫水浴箱 德國GFL公司;868型pH計 美國Orion公司;冷凍高速離心機 德國Eppendorf公司;UV-1800紫外-可見分光光度計 日本Shimadzu公司;proRheo R180黏度計 德國ProRheo公司。
1.4方法
1.4.1菌種活化與菌懸液制備
將菌種保藏管中的嗜熱鏈球菌和保加利亞乳桿菌分別接種于LM17和MRS液體培養基中,轉接活化2 次后,在平板中劃線,挑選單菌落培養,經4 000 r/min離心15 min,收集菌體沉淀,經滅菌的生理鹽水洗滌去除培養基成分,制得菌懸液(OD560nm= 0.95),于4 ℃保存,12 h內使用。
1.4.2發酵乳制備工藝流程
脫脂乳粉(12%)+白砂糖(8%)+凈化水等→40~45 ℃溶解→90~95 ℃殺菌5~10 min→冷卻至42 ℃→接種→恒溫發酵至pH值為4.5→水浴冷卻至20 ℃ →500 r/min攪拌5 min→4 ℃冷藏
接種:分別向滅菌后的脫脂乳中接種保加利亞乳桿菌Lb0925B(簡稱L)、嗜熱鏈球菌StCH-1(簡稱CH-1)、嗜熱鏈球菌StTA040(簡稱040),以及保加利亞乳桿菌與這兩種嗜熱鏈球菌組成的兩組分發酵劑(L+0 4 0、L+C H-1)和三組分復合發酵劑(L+CH-1+040)。乳酸菌接種量為2%,球菌與桿菌的質量添加比例為1∶1。
1.4.3主要指標的測定
pH值測定:用pH計測定發酵乳的pH值。
酸度的測定:以酚酞為指示劑,用0.100 mol/L的NaOH標準溶液滴定。滴定酸度以吉爾涅爾度(?T)表示。
酸化曲線的測定:采用Cinac發酵素酸化監控系統對牛乳發酵過程中pH值的變化進行檢測。
黏度的測定:樣品溫度20 ℃時,將黏度計2號轉子浸沒于樣品中進行測定,穩定后讀取數值。
胞外多糖的分離與含量測定:取300 g發酵乳樣品中加入100 g 16 g/100 mL的三氯乙酸溶液,4 ℃放置2 h后,10 000 r/min離心30 min,取上清液備用;取200 mL上清液,并加入等體積的無水乙醇,4 ℃放置過夜;將混合液10 000 r/min離心30 min,取沉淀,采用苯酚-硫酸法測定多糖的含量[23]。
持水力的測定:取30 g發酵乳樣品,于4 ℃離心30 min,轉速為3 000 r/min,傾去上清液后將離心管倒置10 min,持水力[24]的計算見下式。

發酵乳感官評價:由10 名接受過專業感官分析培訓的食品從業人員分別對樣品的酸甜比、黏度、稠度、口感、風味和質地進行感官評價[25]。評價標準見表1。

表1 酸奶感官評價標準Table 1 Standards for sensory evaluation of yogurt quality
1.5數據處理與分析
所有數據均平行測定3 次,使用SAS 9.3軟件進行方差顯著性分析,Origin 9.0軟件作圖。
2 結果與分析
2.1酸化曲線
利用Cinac發酵素酸化監控系統對發酵過程pH值變化進行監測,如圖1所示。嗜熱鏈球菌和保加利亞乳桿菌單獨發酵時,嗜熱鏈球菌StCH-1的發酵速率最快,保加利亞乳桿菌Lb0925B的發酵速率最慢。當嗜熱鏈球菌與保加利亞乳桿菌共發酵時,L+CH-1的發酵速率明顯比L+040的發酵速率快,這是因為在發酵過程中嗜熱鏈球菌是主要的產酸菌株,發酵速率主要由嗜熱鏈球菌的發酵速率決定[3]。同時,兩組分發酵劑的發酵速率明顯比各自的單菌株發酵速率快。這是由于嗜熱鏈球菌與保加利亞乳桿菌之間存在共生關系,在發酵過程中保 加利亞乳桿菌產生的多肽和游離氨基酸能夠促進嗜熱鏈球菌的生長繁殖和產酸[26]。當嗜熱鏈球菌StCH-1和StTA040與保加利亞乳桿菌Lb0925B組成三組分發酵劑時,其發酵速率在2 種兩組分發酵劑之間。嗜熱鏈球菌StCH-1的發酵速率較快,而嗜熱鏈球菌StTA040的發酵速率較慢,因此三組分發酵劑的發酵速率比L+CH-1的發酵速率慢,而比L+040的發酵速率快。

圖1 單菌株和組合菌株在42 ℃條件下發酵脫脂乳的酸化曲線Fig.1 Acidification curves of single strains and combined strains growing in skim milk at 42 ℃
2.2不同菌種組合發酵乳中胞外多糖的含量
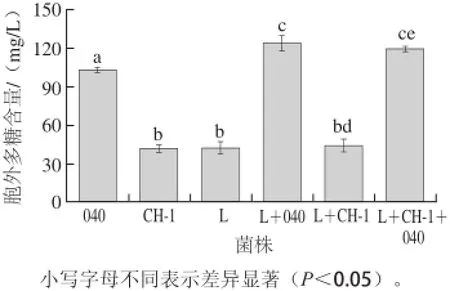
圖2 單菌株和組合菌株發酵酸乳中胞外多糖的含量Fig.2 Contents of EPS in fermented yoghurt by single strains and combined strains
酸奶在發酵過程中,乳酸菌會分泌胞外多糖,它能夠將乳酸菌和酪蛋白連接在一起,賦予酸奶細膩爽滑的口感[5]。單菌株、兩組分發酵劑和三組分發酵劑分泌的胞外多糖含量見圖2。3 種單菌株發酵時,嗜熱鏈球菌StTA040分泌的胞外多糖最多,嗜熱鏈球菌StCH-1和保加利亞乳桿菌Lb0925B分泌的胞外多糖含量較低。當嗜熱鏈球菌與保加利亞乳桿菌共發酵時,組合菌株L+040所分泌的胞外多糖比單菌株嗜熱鏈球菌StTA040分泌的胞外多糖多,而組合菌株L+CH-1所分泌的胞外多糖與單菌株嗜熱鏈球菌StCH-1相比沒有顯著提高。在酸奶發酵過程中,嗜熱鏈球菌是主要的產多糖菌株[2]。雖然保加利亞乳桿菌所分泌的胞外多糖較少,但是保加利亞乳桿菌產生的代謝產物能夠促進嗜熱鏈球菌的生長和胞外多糖的分泌[26]。當嗜熱鏈球菌StCH-1和StTA040與保加利亞乳桿菌Lb0925B組成組合菌株時,其分泌的胞外多糖與兩組合菌株L+040分泌的胞外多糖相當。
2.3發酵乳的貯藏穩定性
2.3.1貯藏期間發酵乳酸度的變化

圖3 貯藏期間發酵乳酸度的變化Fig.3 Change in titratable acidity of fermented milk samples during storage
滅菌后的脫脂乳接種發酵劑,42 ℃發酵至pH值為4.5,然后500 r/min攪拌5 min,4 ℃貯藏。貯藏期間發酵乳酸度的變化見圖3。發酵乳4 ℃貯藏14 d后,酸度都升高了,酸度上升值幾乎相同。在貯藏過程中,乳酸菌繼續生長繁殖消耗殘存的乳糖、半乳糖、葡萄糖產生乳酸,導致發酵乳的酸度進一步升高[27]。Ongol等[28]發現酸奶在低溫貯藏過程中,保加利亞乳桿菌是主要的產酸菌株,并且其產酸能力與H+-ATPase的活性相關。Wang Xinhui等[29]將保加利亞乳桿菌進行誘變,發現H+-ATPase活性降低的菌株不僅其產酸能力降低,并且其后酸化能力也減弱。所以本實驗選擇酸化能力較弱的保加利亞乳桿菌,防止發酵乳在貯藏過程中酸度上升過多。由于3 種組合酸奶發酵劑所含有保加利亞乳桿菌的菌種和濃度相同,因此3 種發酵乳的后酸化程度大致相同。
2.3.2貯藏期間發酵乳黏度的變化

圖4 貯藏期間發酵乳的黏度變化Fig.4 Change in viscosity values of fermented milk samples during storage
3 種發酵乳4 ℃貯藏,其貯藏黏度的變化見圖4。由兩組分發酵劑L+040和三組分發酵劑發酵的發酵乳的黏度顯著高于兩組分發酵劑L+CH-1發酵的發酵乳的黏度。這主要是因為含有嗜熱鏈球菌StTA040的組合菌株所分泌的胞外多糖較多,所以發酵乳的黏度等物理性質更佳。同時,3 種發酵乳在4 ℃貯藏14 d后發酵乳的黏度都變大,這可能是因為發酵乳在貯藏過程中蛋白質分子發生了重組[6]。Tsuda等[7]發現胞外多糖能夠將乳酸菌細胞和酪蛋白鏈接在一起形成網絡結構,使得發酵乳的凝膠結構更加穩定。
2.3.3貯藏期間發酵乳持水力變化
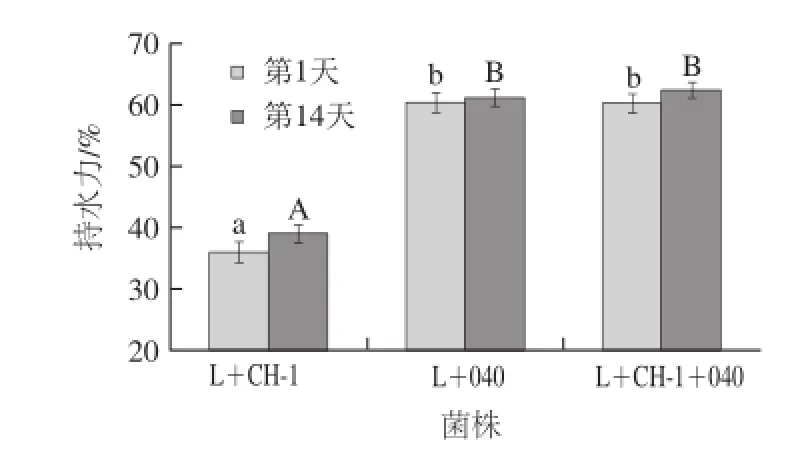
圖5 貯藏期間發酵乳持水力變化Fig.5 Change in water-holding capacity of fermented milk samples during storage
3 種發 酵乳4 ℃貯藏過程中持水力的變化見圖5。3 種發酵乳在4 ℃貯藏14 d后持水力變大,這說明冷藏有助于增加發酵乳的持水力。同時,由兩組分發酵劑L+040和三組分發酵劑發酵的發酵乳的持水力顯著高于兩組分發酵劑L+CH-1發酵的發酵乳的持水力。這主要是因為含有嗜熱鏈球菌StTA040的組合菌株所分泌的胞外多糖較多,所以發酵乳的持水性能更佳。Guzel-Seydim等[30]也發現嗜熱鏈球菌StCH-1產生的發酵乳持水能力較弱,乳清分離嚴重。Prasanna等[6]研究發現胞外多糖能夠有助于發酵乳形成孔狀結構,這種孔狀結構有助于提高發酵乳的持水性能,防止乳清的析出,并且它還能防止發酵乳在加工和運輸工程中結構的破壞,提高發酵乳的穩定性。
2.4發酵乳樣品的感官評價
3 種發酵乳4 ℃貯藏1 d后的感官分析見圖6。3 種不同的酸奶發酵劑發酵的發酵乳在感官上差異較大,其中由三組分發酵劑發酵的發酵乳感官分析結果最佳,其次是兩組分發酵劑L+040發酵的發酵乳,兩組分發酵劑L+CH-1發酵的發酵乳評價結果最差。由于胞外多糖可賦予發酵乳特殊的風味和細膩爽滑的口感,所以含有高產胞外多糖的嗜熱鏈球菌的酸奶發酵劑能夠使發酵乳的質量提高。并且三組分發酵劑發酵的發酵乳比兩組分發酵劑發酵的發酵乳品質更好,這主要是因為多組分發酵劑含有多個優勢菌種,可以使得發酵乳的發酵速率更快,發酵乳的風味和口感更佳。

圖6 專業人員對發酵乳的感官評價Fig.6 Sensory evaluation of fermented milks
3 結 論
兩株不同胞外多糖產生特性和酸化特性的嗜熱鏈球菌與保加利亞乳桿菌進行組合,不同的組合菌株有不同的發酵性能。嗜熱鏈球菌StCH-1發酵速率最快,胞外多糖產量較少,發酵乳黏度低有顆粒感;嗜熱鏈球菌StTA040發酵速率較慢,胞外多糖產量最多,發酵乳黏度高,凝乳狀態均勻。當兩株嗜熱鏈球菌與保加利亞乳桿菌Lb0925B組合后,發酵速率均比單菌株發酵速率快,顯著縮短了發酵時間。其中含有嗜熱鏈球菌StCH-1的組合菌株的發酵速率較快,而含有嗜熱鏈球菌StTA040的組合菌株的胞外多糖產量較高,發酵乳的黏度較高,持水力較強。三組分發酵劑的發酵速率快,生產的發酵乳黏度好,持水力強,具有3 種菌種的所有優良性能,同時通過感官分析得出三組分發酵劑制得的發酵乳的口感、風味和貯藏穩定性較好,發酵乳的整體質量最佳。
[1] PURWANDARI U, SHAH NP, VASILJEVIC T. Effects of exopolysaccharide producing strains of Streptococcus thermophilus on technological and rheological properties of set-type yoghurt[J]. International Dairy Journal, 2007, 17: 1344-1352.
[2] SETTACHAIMONGKON S, ROBERT NOUT M J, ANTUNES FERNANDES E C, et al. Influence of different proteolytic strains of Streptococcus thermophilus in co-culture with Lactobacillus delbrueckii subsp. bulgaricus on the metabolite profile of set-yoghurt[J]. International Journal of Food Microbiology, 2014, 177: 29-36.
[3] IYER R, TOMAR S K, UMA MAHESWARI T, et al. Streptococcus thermophilus strains: multifunctional lactic acid bacteria[J]. International Dairy Journal, 2010, 20: 133-141.
[4] SKRIVER A, STENBY E, FOLKENBERG D M, et al. Tools in the development of future starter cultures for fermented milk[C]// Proceedings of the IDF Seminar on Aroma and Texture of Fermented Milk. Kolding, Denmark, June 3-5, 2002. International Dairy Federation, Brussels, Belgium, 2003: 55-61.
[5] PHAM P L, DUPONT I, ROY D, et al. Production of exopolysaccharide by Lactobacillu s rhamnosus R and analysis of itseneymaticdegradation during prolonged fermentation[J]. Applied and Environmental Microbiology, 2000, 66(6): 2302-2310.
[6] PRASANNA P H P, GRAND ISON A S, CHARALAMPOPOULOS D. Microbiological, chemical and rheological properties of low fat set yogurt produced with exopolysaccharide (EPS) producing Bifi dobacterium strains[J]. Food Research International, 2013, 51: 15-22.
[7] TSUDA H, MIYAMOTO T. Production of exopolysaccharide by Lactobacillus plantarum and the prebiotic activity of the exopolysaccharide[J]. Food Science and Technology Research, 2010,16: 87-92.
[8] 李全陽. 酸乳質量及其胞外多糖的研究[D]. 無錫: 江南大學, 2004.
[9] LAWS A P, MARSHALL V M. The relevance of exopolysaccharides to the rheological properties in milk fermented with ropy strains of lactic acid bacteria[J]. International Dairy Journal, 2001, 11: 709-721.
[10] RUAS-MADIEDO P, HUGENHOLTZ J, ZOON P. An overview of the functionality of exopolysaccharides produced by lactic acid bacteria[J]. International Dairy Journal, 2002,12: 163-171.
[11] URSHEV Z L, PASHOVA-BALTOVA K N, DIMITROV Z P. Tracing Streptococcus thermophilus strains in three-component yogurt starters[J]. World Journal of Microbiology and Biotechnology, 2006,22: 1223-1228.
[12] 張雪, 李達, 趙玉娟, 等. 內蒙古奶豆腐中產胞外多糖乳酸菌的分離篩選[J]. 食品科學, 2010, 31(1): 141-144.
[13] SOOMRO A H, MASUD T. Selection of yoghurt starter culture from indigenous isolates of Streptococcus thermophilus and on the basis of technological properties[J]. Annals of Microbiology, 2008,58(1): 67-71.
[14] HAN Xue, ZHANG Lanwei, YU Peng, et al. Potential of LAB starter culture isolated from Chinese traditional fermented foods for yoghurt production[J]. International Dairy Journal, 2014, 34: 247-251.
[15] SHENE C, CANQUIL N, BRAVO S, et al. Production of the exopolysacchzrides by Streptococcus thermophilus: effect of growth conditions on fermentation kinetics and intrinsic viscosity[J]. International Journal of Food Microbiology, 2008, 124: 279-284.
[16] PRASANNA P H P, GRANDISON A S, CHARALAMPOPOULOS D. Effec t of dairy-based protein sources and temperature on growth,acidification and exopolysaccharide production of Bifi dobacterium strains in skim milk[J]. Food Research International, 2012, 47: 6-12.
[17] 徐愛才, 馬成杰, 華寶珍, 等. 嗜熱鏈球菌與不同乳桿菌組合在脫脂乳中的發酵特性[J]. 食品科學, 2013, 34(15): 158-161. doi: 10.7506/ spkx1002-6630-201315032.
[18] 靳志敏, 通力嘎, 段艷, 等. 1株高效將膽固醇乳酸菌的功能與發酵特性[J]. 食品科學, 2014, 35(11): 120-123. doi: 10.7506/spkx1002-6630-201411024.
[19] DONG Zixing, GU Lei, ZHANG Juan, et al. Optimisation for high cell density cultivation of Lactobacillus salivarius BBE 09-18 with response surface methodology[J]. International Dairy Journal, 2014,34: 230-236.
[20] 華寶珍, 李莎, 徐愛才, 等. 植物乳桿菌ST-Ⅲ脫脂乳的發酵工藝優化[J]. 農業工程學報, 2014, 30(11): 276-284.
[21] SCHWAB C, VOGEL R, G?NZLE M G. Influence of oligosaccharides o n the viability and membrane properties of Lactobacillus reuteri TMW.106 during freeze-drying[J]. Cryobiology,2007, 55(2): 108-114.
[22] ASCHENBRENNER M, KULOZIK U, F?RST P. The role of the glassy state in production and storage of freeze-dried starter cultures[J]. Procedia Food Science, 2011, 1: 347-354.
[23] ROBITAILLE G, TREMBLAY A, MOINEAU S, et al. Fat-free yogurt made using a galactose-positiveexopolysaccharide-producing recombinant strain of Streptococcus thermophilus[J]. Journal of Dairy Science, 2009, 92: 477-482.
[24] 馬成杰, 杜昭平, 華寶珍, 等. 植物乳桿菌ST-Ⅲ在豆乳中的發酵特性及發酵豆乳的貯藏穩定性[J]. 食品科學, 2013, 34(5): 151-155. doi: 10.7506/spkx1002-6630201305032.
[25] ESCOBAR D, CLARK S, GANESAN V, et al. High-pressure homogenization of raw and pasteurize d milk modifies the yield,composition, and texture of queso fresco cheese[J]. Journal of Dairy Science, 2011, 94: 1201-1210.
[26] SIEUWERTS S, de BOK F A M, HUGENHOLTZ J, et al. Unravelingmicrobial interactions in food fermentations: from classical to genomics approaches[J]. Applied and Environmental Microbiology,2008, 74: 4997-5007.
[27] BEAL C, SKOKANOVA J, LATRILLE E, et al. Combined effects of culture conditions and storage time on acidification and viscosity of stirred yogurt[J]. Journal of Dairy Science, 1999, 82: 673-681.
[28] ONGOL M P, SAWATANI Y, EBINA Y, et al. Yogurt fermented by Lactobacillus delbrueckii subsp. bulgaricus H+-ATPase-defective mutants exhibits enhanced viability of Bifi dobacterium breve during storage[J]. International Journal of Food Microbiology, 2007, 116: 358-366.
[29] WANG Xinhui, REN Hongyang, LIU Dayu, et al. H+-ATPasedefective variants of Lactobacillusdelbrueckii subsp. bulgaricus contribute to inhibition of postacidification of yogurt during chilled storage[J]. Journal of Food Science, 2013, 78: M297-M302.
[30] GUZEL-SEYDIM Z B, SEZGIN E, SEYDIM A C. Influences of exopol ysaccharide producing on the quality of plain set type yogurt[J]. Food Control, 2005, 16: 205-209.
Fermentation Characteristics of Lactobacillus delbrueckii subsp. bulgaricus Co-cultured with Different Streptococcus thermophilus Strains
LI Sha, MA Chengjie*, XU Zhiping, XU Aicai, HUA Baozhen
(State Key Laboratory of Dairy Biotechnology, Technology Center of Bright Dairy and Food Co. Ltd., Shanghai 200436, China)
Streptococcus thermophlius StCH-1, a strain capable of rapidly producing acid and EPS-producing S. thermophlius StTA040, a strain with an improved ability to produce exopolysaccharides (EPS) were individually or togethe r co-cultured with L. bulgaricus strain (Lb0925B) to investigate their fermentation characteristics. The influence of mixed strains on fermentation curves, EPS production, post-acidification, viscosity, water-holding capacity and sensory quality of fermented milk was investigated. The results showed that the fermentation time was significantly shortened by the combined use of the strains especially in the presence of StCH-1. The combined strains containing StTA040 could produce more exopolysaccharides (EPS) and consequently higher viscosity and water-holding capacity in fermented milk. In addition,the combination of all these three strains accelerated the fermentation process and produced fermented milk with higher viscosity and water-holding capacity and the best taste and flavor by sensory evaluation.
Streptococcus thermophilus; exopolysaccharides; co-culture; fermention properties; sensory evaluation
TS252.1
A
1002-6630(2015)15-0123-05
10.7506/spkx 1002-6630-201515023
2014-07-30
“十二五”國家科技支撐計劃項目(2013BAD18B01)
李莎(1987—),女,碩士,主要從事發酵乳制品研究與開發。E-mail:lisha@brightdairy.com
馬成杰(1976—),男,高級工程師,碩士,主要從事乳品生物技術研究。E-mail:machengjie@brightdairy.com
