咖啡幼苗對(duì)鎘脅迫的生理響應(yīng)
2015-11-05 11:08:56王婭玲李維峰王永林蘇志龍
安徽農(nóng)業(yè)科學(xué) 2015年12期
王婭玲,李維峰,王永林,蘇志龍
(1.云南農(nóng)業(yè)大學(xué)熱帶作物學(xué)院,云南普洱665000;2.普洱學(xué)院,云南普洱665000)
鎘是一種高生物毒性重金屬元素,是我國(guó)農(nóng)田重金屬污染的主要元素之一[1]。有研究表明,低濃度的鎘能夠刺激植物生長(zhǎng)發(fā)育,但是會(huì)給植物帶來(lái)更多的是毒害效應(yīng)[2-4]。近年來(lái),有關(guān)鎘對(duì)植物的毒害作用國(guó)內(nèi)外進(jìn)行了較多的研究。諸多研究表明,鎘能夠誘導(dǎo)植物體內(nèi)活性氧的大量增加,引起細(xì)胞膜質(zhì)過(guò)氧化[5-7],從而影響植物生長(zhǎng)發(fā)育,嚴(yán)重時(shí)甚至導(dǎo)致植物死亡。此外,鎘還能夠通過(guò)食物鏈進(jìn)入人體,對(duì)人類(lèi)健康產(chǎn)生極大危害[8]。因此,研究鎘對(duì)植物生理特征的影響對(duì)于指導(dǎo)農(nóng)業(yè)生產(chǎn)及保護(hù)農(nóng)業(yè)生態(tài)環(huán)境具有重要的意義。目前,已有很多關(guān)于鎘對(duì)植物生理影響的研究[9-13],但是對(duì)于鎘對(duì)咖啡幼苗生理特征的影響還未見(jiàn)到報(bào)道。筆者采用盆栽法,研究在不同濃度梯度鎘脅迫下咖啡幼苗葉片葉綠素和抗氧化酶的變化情況,探討重金屬鎘脅迫對(duì)咖啡幼苗超氧化物歧化酶(SOD)、過(guò)氧化物酶(POD)、過(guò)氧化氫酶(CAT)、丙二醛(MDA)以及葉綠素含量的影響,為揭示咖啡受到鎘脅迫后響應(yīng)機(jī)制提供理論參考。
1 材料與方法
1.1 試驗(yàn)材料 試驗(yàn)所用咖啡幼苗來(lái)自云南熱帶作物職業(yè)學(xué)院溫室育苗大棚,品種為卡蒂姆P3。咖啡種子在沙床上育苗,待幼苗生長(zhǎng)42 d后,選擇長(zhǎng)勢(shì)均勻、具三對(duì)葉的幼苗進(jìn)行試驗(yàn)。洗凈幼苗根部泥沙,用濃度0.1%高錳酸鉀浸泡根部,消毒10 min,將消毒好的咖啡幼苗用去離子水沖洗干凈。培養(yǎng)液母液為Hoagland-Arnon營(yíng)養(yǎng)液。鎘脅迫溶液則用氯化鎘配制,先將氯化鎘與去離子水配制成1 g/L的溶液,然后按照不同比例加入Hoagland-Arnon營(yíng)養(yǎng)液中,配制成0、10、20、40、60、80 mg/L 6個(gè)梯度濃度的鎘脅迫溶液。用濃度0.1%高錳酸鉀浸泡消毒60~80目分析純石英砂,蒸餾水洗凈、烘干后用作栽培基質(zhì)。
1.2 試驗(yàn)設(shè)計(jì) 試驗(yàn)于2013年7~9月在云南熱帶作物職業(yè)學(xué)院智能溫室中進(jìn)行。用口徑18 cm、高13 cm的花盆,塑料盆消毒洗凈后裝入適量石英砂,每盆植入4株咖啡幼苗。將栽好的試驗(yàn)盆放置于溫室大棚內(nèi)培養(yǎng),先用Hoagland-Arnon營(yíng)養(yǎng)液澆灌,待咖啡幼苗恢復(fù)正常生長(zhǎng)7 d后再將配好的鎘脅迫營(yíng)養(yǎng)液分別加入不同花盆內(nèi)進(jìn)行脅迫試驗(yàn)。每盆每日澆灌20 ml不同脅迫濃度的鎘溶液,使得盆內(nèi)砂基濕度維持在75%左右。每個(gè)脅迫濃度設(shè)3次重復(fù)。脅迫處理2個(gè)月后取樣,測(cè)定各項(xiàng)生理指標(biāo)。
1.3 測(cè)定項(xiàng)目與方法 SOD活性采用氮藍(lán)四唑(NBT)比色法[14]測(cè)定;POD 活性采用愈創(chuàng)木酚法[15]測(cè)定;CAT活性采用紫外吸收法[16]測(cè)定;MDA含量采用硫代巴比妥酸(TBA)比色法[17]測(cè)定;葉綠素含量采用分光光度法[15]測(cè)定。
1.4 數(shù)據(jù)分析 試驗(yàn)數(shù)據(jù)用Excel2003進(jìn)行基本處理。結(jié)果用SPSS進(jìn)行方差分析、顯著性檢驗(yàn)。
2 結(jié)果與分析
2.1 鎘脅迫對(duì)咖啡幼苗葉綠素含量的影響 葉綠素是植物利用光能的主要色素。葉綠素含量是植物光合能力、營(yíng)養(yǎng)脅迫和發(fā)育情況的重要指標(biāo)。從表1可以看出,與空白對(duì)照相比,鎘脅迫下葉綠素a、b及葉綠素總量均顯著降低(P<0.05);在鎘濃度達(dá)80 mg/L時(shí),葉綠素a、葉綠素b含量分別下降了24.13%和20.76%,葉綠素總量下降了24.45%。
2.2 鎘脅迫對(duì)咖啡幼苗POD活性的影響 POD是植物體內(nèi)清除活性氧酶系統(tǒng)的重要組成部分,在控制膜脂的過(guò)氧化、清除超氧自由基等方面起重要的作用。從表2可以看出,POD活性在低濃度鎘脅迫下上升,在鎘脅迫強(qiáng)度>20 mg/L后POD活性下降至低于空白對(duì)照,在鎘脅迫濃度為80 mg/L時(shí)POD活性下降了20.96%。
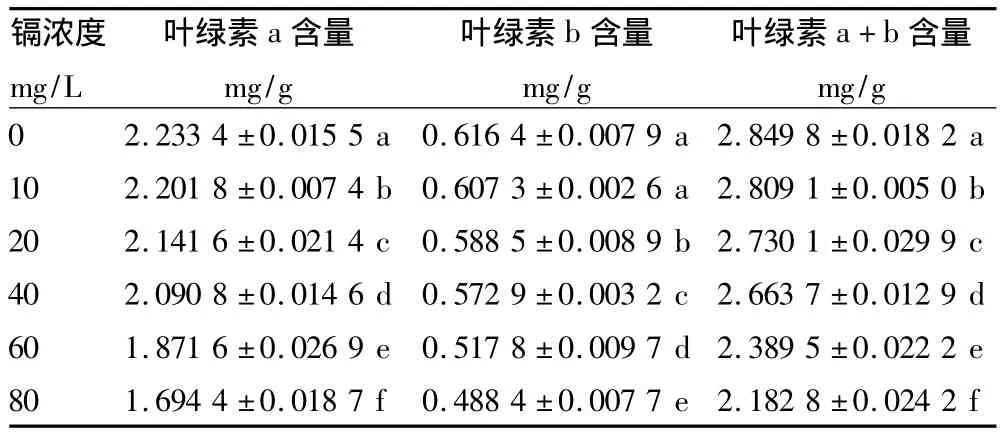
表1 不同濃度鎘脅迫下咖啡幼苗葉片葉綠素含量
2.3 鎘脅迫對(duì)咖啡幼苗CAT活性的影響 CAT是植物細(xì)胞內(nèi)清除活性氧的一種重要的抗氧化酶。它可以將過(guò)氧化氫分解為水和氧氣,從而維持細(xì)胞內(nèi)過(guò)氧化氫的正常水平。從表2可以看出,在鎘濃度<10 mg/L脅迫下,CAT活性呈上升趨勢(shì);當(dāng)鎘濃度>20 mg/L時(shí),CAT活性較空白對(duì)照明顯下降;在鎘脅迫濃度為80 mg/L時(shí),與對(duì)照相比,CAT活性下降了32.35%。
2.4 鎘脅迫對(duì)咖啡幼苗SOD活性的影響 SOD能夠清除活性氧自由基,其活性與植物適應(yīng)逆境能力密切相關(guān)。從表2可以看出,鎘脅迫對(duì)咖啡幼苗SOD活性影響顯著(P<0.05)。在低濃度鎘脅迫下,咖啡幼苗SOD活性呈上升趨勢(shì);在10 mg/L鎘脅迫下,SOD活性比空白對(duì)照上升了11.78%;但在鎘濃度>20 mg/L時(shí),SOD活性明顯受到抑制,在80 mg/L鎘脅迫時(shí)SOD活性?xún)H為空白對(duì)照的70.9%。
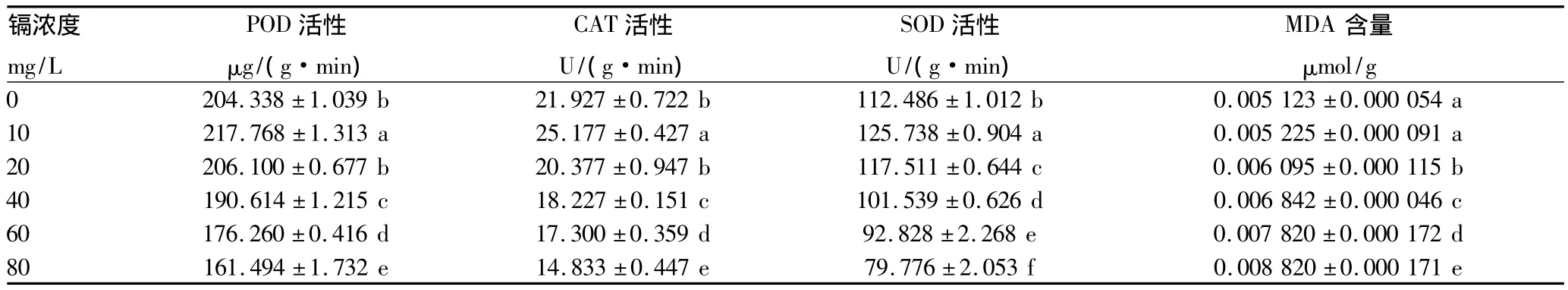
表2 不同濃度鎘脅迫下咖啡幼苗抗氧化酶活性及MDA含量
2.5 鎘脅迫對(duì)咖啡幼苗MDA活性的影響 細(xì)胞膜的穩(wěn)定是植物進(jìn)行正常生理代謝的基礎(chǔ)。在逆境脅迫下,植物細(xì)胞內(nèi)產(chǎn)生的活性氧會(huì)使細(xì)胞膜上的不飽和脂肪酸發(fā)生活性氧反應(yīng),其產(chǎn)物MDA含量是反映細(xì)胞膜受損程度的一個(gè)重要指標(biāo)。從表2可以看出,在重金屬鎘脅迫下,咖啡幼苗葉片MDA含量隨鎘濃度的增加而顯著增加(P<0.05),80 mg/L鎘脅迫下脅迫咖啡幼苗MDA含量比對(duì)照增加了41.9%,說(shuō)明細(xì)胞膜受到較嚴(yán)重的損害。
3 結(jié)論與討論
鎘是植物的生長(zhǎng)的非必需元素。它被植物的吸收后會(huì)對(duì)植物生長(zhǎng)發(fā)育造成危害。研究中,在0~80 mg/L濃度鎘脅迫下,咖啡幼苗體內(nèi)葉綠素a、葉綠素b含量均隨著鎘濃度的增加而顯著降低(P<0.05),而且鎘濃度越高,葉綠素總量下降越多。這可能是由于鎘脅迫下咖啡幼苗體內(nèi)活性氧增加,從而導(dǎo)致葉綠素降解。此外,鎘也會(huì)影響有關(guān)葉綠素合成相關(guān)酶的活性,導(dǎo)致葉綠素含量降低[18]。
在植物體內(nèi),POD、CAT和SOD等酶共同組成抗氧化酶體系,協(xié)同防御活性氧自由基對(duì)植物細(xì)胞產(chǎn)生傷害,維持植物體內(nèi)代謝平衡,增強(qiáng)植物的抗逆境脅迫能力,延緩葉綠素、蛋白質(zhì)等降解。研究表明,咖啡幼苗葉片POD、CAT及SOD活性在鎘濃度≤10 mg/L的脅迫下隨鎘濃度增加而升高,而后當(dāng)鎘濃度>20 mg/L時(shí),酶活性隨鎘濃度的增加而降低。這可能是由于高濃度的鎘影響3種酶的空間結(jié)構(gòu)或活性氧濃度過(guò)高導(dǎo)致酶合成受到抑制。
在正常生長(zhǎng)情況下,植物體內(nèi)活性氧自由基產(chǎn)生與清除處于動(dòng)態(tài)平衡狀態(tài),但是逆境脅迫會(huì)打破這種平衡,導(dǎo)致植物體內(nèi)活性氧自由基的大量產(chǎn)生,而葉片中活性氧自由基會(huì)引起膜質(zhì)過(guò)氧化作用,造成細(xì)胞膜損傷,影響植物正常生理代謝。MDA是膜脂過(guò)氧化的重要產(chǎn)物,其含量是判斷膜脂過(guò)氧化程度的一個(gè)重要指標(biāo)[19]。研究中,MDA含量隨著鎘濃度的升高而顯著增加,說(shuō)明在鎘脅迫下咖啡幼苗細(xì)胞膜發(fā)生較嚴(yán)重的過(guò)氧化,其生長(zhǎng)代謝受到較大的影響。
[1]QIU Q,WANG Y T,YANG Z Y,et al.Responses of different Chinese flowering cabbage(Brassica parachinensis L.)cultivars to cadmium and lead exposure:Screening for Cd+Pb pollution-safe cultivars[J].Clean-Soil,Air,Water,2011,39:925-932.
[2]張玲,李俊梅,王煥校.鎘脅迫下小麥根系的生理生態(tài)變化[J].土壤通報(bào),2002,33(1):61-65.
[3]崔瑋,張芬琴,金自學(xué).Cd2+處理對(duì)兩種豆科作物幼苗生長(zhǎng)的影響[J].農(nóng)業(yè)環(huán)境科學(xué)學(xué)報(bào),2004,23(1):60-63.
[4]GREGER M,LINDBERG S.Effects of Cd2+and EDTA on young sugar beets(Beta vulgaris).I.Cd2+uptake and sugar accumulation[J].Physiol Plantarum,1986,66(1):69-74.
[5]TEWARI R K,KUMAR P,SHARMA P N.Morphology and physiology of zinc-stressed mulberry plants[J].Journal of Plant Nutrition and Soil Science,2008,171(2):286-294.
[6]WANG C,ZHANG S H,WANG PF,et al.The effect of excess Zn on mineral nutrition and antioxidative response in rapeseed seedlings[J].Chemosphere,2009,75(11):1468-1476.
[7]CAO H C,LUAN Z Q,WANG J D,et al.Potential ecological risk of cadmium,lead and arsenic in agricultural black soil in Jilin Province,China[J].Stochastic Environmental Research and Risk Assessment,2009,23(1):57-64.
[8]龔玉蓮,楊中藝.蕹菜典型品種的根系形態(tài)學(xué)特征及與Cd吸收積累的關(guān)系[J].華南師范大學(xué)學(xué)報(bào):自然科學(xué)版,2012,44(3):100-106.
[9]馮世靜,楊途熙,張艷軍,等.鎘脅迫對(duì)楊樹(shù)光合特性的影響[J].農(nóng)業(yè)環(huán)境科學(xué)學(xué)報(bào),2013,32(3):539-547.
[10]YIDMZ D D,PARLAK K U.Changes in proline accumulation and antioxidative enzyme activities in Groenlandia densa under cadmium stress[J].Ecological Indicators,201l,11(2):417-423.
[11]YING R R,QIU R L,TANG Y T,et al.Cadmium tolerance of carbon assimilation enzymes and chloroplast in Zn/Cd hyperaccumulator Picris divaricata[J].Journal of Plant Physiology,2010,167(2):81-87.
[12]GOGORCENA Y,LARBI A,ANDALUZ S,et al.Effects of cadmium on cork oak(Quercus suber L.)plants grown in hydroponics[J].Tree Physiology,2011,31(12):1401-1412.
[13]MOHAMED A A,CASTAGNA A,RANIERI A,et al.Cadmium tolerance in Brassica juncea roots and shoots is affected by antioxidant status and phytochelatin biosynthesis[J].Plant Physiology and Biochemistry,2012,57:15-22.
[14]沈文飚,張榮銑.氮藍(lán)四唑光化還原法測(cè)定超氧化物歧化酶活性的適宜條件[J].南京農(nóng)業(yè)大學(xué)學(xué)報(bào),1996,19(2):101-102.
[15]郝建軍,劉延吉.植物生理學(xué)試驗(yàn)技術(shù)[M].沈陽(yáng):遼寧科學(xué)技術(shù)出版社,2001:178-190.
[16]陳曉敏.測(cè)定切花中過(guò)氧化氫酶活性的3種常用方法的比較[J].熱帶農(nóng)業(yè)科學(xué),2002,22(5):13-16.
[17]高俊鳳.植物生理學(xué)實(shí)驗(yàn)指導(dǎo)[M].北京:高等教育出版社,2006:74-77.
[18]惠俊愛(ài),黨志,葉慶生.鎘脅迫對(duì)玉米光合特性的影響[J].農(nóng)業(yè)環(huán)境科學(xué)學(xué)報(bào),2010,29(2):205-210.
[19]陳會(huì),任艷芳,陳秀蘭,等.鎘脅迫下不同耐性水稻植株幼苗生長(zhǎng)和抗氧化酶的變化[J].江西農(nóng)業(yè)大學(xué)學(xué)報(bào),2012,34(6):1099-1104.
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年3期)2017-02-21 01:22:28
爆笑show(2016年7期)2017-02-09 09:36:13
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
