硫色鐮刀菌(Fusarium sulphureum)接種對抗病/易感品種馬鈴薯塊莖苯丙烷代謝的影響比較
2015-12-13 03:41:18包改紅李永才馬朝玲白小東
食品科學 2015年6期
包改紅,畢 陽*,李永才,王 毅,王 婷,唐 瑛,馬朝玲,白小東
(甘肅農業大學食品科學與工程學院,甘肅 蘭州 730070)
硫色鐮刀菌(Fusarium sulphureum)接種對抗病/易感品種馬鈴薯塊莖苯丙烷代謝的影響比較
包改紅,畢 陽*,李永才,王 毅,王 婷,唐 瑛,馬朝玲,白小東
(甘肅農業大學食品科學與工程學院,甘肅 蘭州 730070)
以抗病品種“青薯 168”和易感品種“隴薯3號”馬鈴薯塊莖為試材,比較2個品種馬鈴薯塊莖、切片接種硫色鐮刀菌(F. sulphureum)后苯丙烷代謝關鍵酶活性和相關產物積累的動態變化。結果表明,“青薯168”馬鈴薯塊莖及其切片接種F. sulphureum后的病斑直徑顯著小于“隴薯3號”。2個品種馬鈴薯接種F. sulphureum后,在前期苯丙氨酸解氨酶(PAL)、4-香豆酰-輔酶A連接酶(4CL)活性以及總酚、類黃酮含量增加,但隨著病斑的擴展,PAL、4CL活性以及總酚、類黃酮含量迅速降低并低于未接菌對照。肉桂酸-4-羥化酶(C4H)活性和木質素含量在整個培養過程中均顯著低于未接菌對照 。抗病品種“青薯168”接種F. sulphureum后,PAL、4CL和C4H活性以及總酚、類黃酮和木質素含量均顯著高于“隴薯3號”。表明苯丙烷代謝在F. sulphureum與馬鈴薯塊莖互作的早期發揮了積極的作用,一旦病斑開始擴展,苯丙烷代謝的抗病作用將顯著降低;抗病塊莖的苯丙烷代謝活性顯著高于感病塊莖,表明苯丙烷代謝在馬鈴薯塊莖抗干腐病方面具有重要的作用。
馬鈴薯;硫色鐮刀菌;接種;苯丙烷代謝
馬鈴薯(Solanum tuberosum)是甘肅省重要的經濟作物,塊莖采收后一般要經過3~6 個月甚至更長時間的貯藏,期間腐爛頗為嚴重,腐爛率達20%~25%,經濟損失巨大[1-2]。干腐病是造成馬鈴薯塊莖采后損失最主要的病害,由其引起的塊莖腐爛約占病薯的88.5%[3]。多種鐮刀菌(Fusarium spp.)與干腐病的發生密切相關,其中硫色鐮刀菌(F. sulphureum)是引起甘肅省馬鈴薯塊莖干腐病最主要的病原物[4-5]。苯丙烷代謝在植物抗病防衛反應中具有重要作用[6],主要的抗菌物質如酚類化合物、植保素、木質素、類黃酮等均需經過該途徑合成[7-8]。此外,苯丙烷代謝還是馬鈴薯塊莖愈傷的物質代謝基礎,在減輕采后病害的發生中具有積極的作用[9]。因此,探討馬鈴薯塊莖接種F. sulphureum后苯丙烷代謝的差異,對于深入 了解塊莖的抗病機理,以及抗病品種的選育和干腐病的控制具有重要意義。
病原物侵染通常會激活植物體內的苯丙烷代謝,以此來增強寄主的抗病性[10]。如F. oxysporum和F. culmorum侵染亞麻后苯丙烷代謝的關鍵基因被激活,p-香豆酸、綠原酸和咖啡酸的含量均有不同程度的提高[11]。Rhizoctonia solani侵染豆類[12]和水稻[13]后激活了寄主體內一系列的防衛相關基因,導致病程相關蛋白和苯丙烷代謝產物的積累。R. solani和Phytophthora infestans侵染馬鈴薯后苯丙烷代謝關鍵酶苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)的活性和代謝產物的積累顯著增強[14-15]。比較不同抗性蘋果品種苯丙烷代謝的差異時發現,抗病品種的代謝活性要顯著高于感病品種[16]。Ustilago scitaminea侵染甘蔗后抗病品種后PAL和4-香豆酰-輔酶A連接酶(4-coumarate A ligase,4CL)活性上升的幅度和持續時間顯著大于感病品種,綠原酸和黃酮類化合物的含量也顯著增高[17]。但上述研究均以生長狀態的植株為試材,缺乏病原物侵染不同抗/感采后果蔬的苯丙烷代謝差異比較。
本研究擬以抗病品種“青薯168”和易感病品種“隴薯3號”馬鈴薯塊莖和切片為試材,研究F. sulphureum接種對馬鈴薯塊莖干腐病病斑直徑的影響,比較接種后塊莖組織苯丙烷代謝的差異,以期揭示苯丙烷代謝在馬鈴薯塊莖與鐮刀菌互作中的抗病作用,為干腐病的控制和抗病品種的選育提供理論依據。
1 材料與方法
1.1 材料
供試馬鈴薯“青薯168”和“隴薯3號”分別于2012年10月采自青海省農林科學院作物育種栽培研究所和甘肅省渭源縣會川鎮。選取外觀整齊,大小一致,無病蟲害,無損傷的馬鈴薯塊莖裝入網袋并于當日運回實驗室,并在5~8 ℃條件下貯藏。塊莖用2%的次氯酸鈉溶液表面消毒2 min,然后用無菌水沖洗,晾干待用。
供試病原菌F. sulphureum由甘肅省農業科學院植物保護研究所提供,病原物在馬鈴薯葡萄糖瓊脂培養基(potato dextrose agar,PDA)培養基上黑暗培養7 d待用。
1.2 方法
1.2.1 切片接種
參照楊志敏等[18]的方法。用無菌刀和打孔器把馬鈴薯塊莖制成圓片,圓片厚1 cm,直徑45 mm。切片用無菌水清洗后再用75%酒精擦洗,并在酒精燈火焰上灼燒,以去除多余酒精,之后置于已滅菌且鋪有無菌濕濾紙的托盤上,黑暗恒溫恒濕培養4 h后,在切片中央接種培養1 周的F. sulphureum菌餅(直徑6.5 mm),含菌絲面與切片接觸,并在23~25 ℃,相對濕度(realative humidity,RH)為72%~75%條件下培養2~3 d后測定病斑直徑。每個處理包括10 個切片,重復3 次。
1.2.2 塊莖接種
參照Li Yongcai等[1]的方法。病原物在PDA培養基上培養1 周以后,在培養皿中加5 mL含有0.05% Tween 80的無菌水。懸浮液用4 層紗布過濾以除去菌絲,用顯微鏡計數并將袍子懸浮液的濃度調整為1.0×106孢子/mL待用。在馬鈴薯的赤道部位均勻的打3 個3 mm深、直徑3 mm的小孔,1 h后每個孔接種20 ?L F. sulphureum的孢子懸浮液。用聚乙烯袋包裝后裝入紙箱中在室溫(25±2)℃條件下貯藏10、12、14、16 d后測病斑直徑。每個處理包括5 個馬鈴薯塊莖,重復3 次。
1.2.3 取樣
參照楊志敏等[18]的方法并稍作修改。按照1.2.1節描述的方法制作馬鈴薯塊莖切片,并接種F. sulphureum菌餅(8 mm),在23~25 ℃、RH為72%~75%條件下培養。以接種相同大小PDA培養基作為對照。分別在接種后第0、1、2、3、4、5、6 d取接種表面(3 mm厚)的馬鈴薯組織3 g,用錫箔紙包裝后液氮冷凍并在—80 ℃條件下貯藏待用。
1.2.4 苯丙烷代謝相關酶和產物的測定
1.2.4.1 PAL活性的測定
PAL活性的測定參照Yin Yan等[19]的方法。稱取3 g樣品置于預冷的研缽中,加入5 mL的經4 ℃預冷的100 mmol/L硼酸緩沖液(pH 8.8,含1%聚乙烯吡咯烷酮、1 mmol/L乙二胺四乙酸和50 mmol/L β-巰基乙醇),冰浴條件下充分研磨成勻漿后全部轉入到離心管中,于4 ℃、11 250 ×g離心20 min,收集上清液立即用于PAL活性的測定。取3 支試管分別標記為Ⅰ、Ⅱ和Ⅲ,然后向Ⅰ和Ⅱ2支試管分別加入3 mL 7 mmol/L L-苯丙氨酸溶液(用50 mmol/L硼酸緩沖液配制),再分別向Ⅰ和Ⅲ2支試管中加入500 μL上清液后立即于37 ℃保溫1 h,保溫完畢后立即在290 nm波長測定其光密度值,以Ⅱ和Ⅲ混合后測定的光密度值為初始值,Ⅰ中測定值為終止值。另以提取液代替上清液按照上述方法測得的光密度值分別作為初始值和終止值的對照。PAL活性表示為0.01 ΔOD290nm/(h·g)。
1.2.4.2 肉桂酸-4-羥化酶(cinnamate-4-hydroxylase,C4H)和4CL活性的測定
C4H活性測定參照范存斐等[20]的方法。稱取3 g樣品置于預冷的研缽中,加入5 mL的經4 ℃預冷的提取液(50 mmol/L Tris-HCl pH 8.9、4 mmol/L MgCl2、15 mmol/L β-巰基乙醇、10 μmol/L亮抑酶肽、5 mmol/L還原型抗壞血酸、1 mmol/L苯甲基磺酰氟、0.15%聚乙烯吡咯烷酮、10%甘油),冰浴條件下充分研磨成勻漿后用4 層紗布過濾,然后于4 ℃、9 000×g離心20 min,收集上清液立即用于C4H活性的測定。在一支試管中加入0.8 mL上清液和2.0 mL緩沖液(8 μmol/L反式肉桂酸、50 mmol/L Tris-HCl pH 8.9、3 μmol/L NADPNa2、6 μmol/L六磷酸葡萄糖二鈉)。搖勻后25 ℃條件下振蕩反應30 min,之后加入100 μL 6 mol/L HCl溶液終止反應,在4 ℃、9 000 ×g離心10 min,取上清液在340 nm波長處測定其光密度值,用提取液代替上清液作為參比。以每分鐘光密度值變化0.1為1 個C4H活力單位,酶活性表示為0.1 ΔOD340nm/(min·g)。
4CL活性測定參照范存斐等[20]的方法并修改。稱取3 g樣品置于預冷的研缽中,加入5 mL的經4 ℃預冷的0.2 mol/L Tris-HCl緩沖液(pH 8.0,含25%甘油和0.1 mol/L DTT),冰浴條件下充分研磨成勻漿后全部轉入到離心管中,于4 ℃、11 250×g離心20 min,收集上清液立即用于4CL活性的測定。取一支試管,向其中加入0.45 mL 15 μmol/L MgCl2,0.15 mL 5 μmol/mL p-香豆酸,0.15 mL 50 μmol/mL ATP,0.15 mL 1 μmol/mL CoA和0.5 mL上清液(對照為不加p-香豆酸),在333 nm波長處測定其光密度值,4CL活性表示為0.01 ΔOD333nm/(min·g)。
1.2.4.3 總酚和類黃酮含量的測定
總酚和類黃酮含量的測定參照Yin Yan等[19]的方法。稱取3 g樣品置于預冷的研缽中,加入5 mL的經4 ℃預冷的1% HCl-甲醇溶液,冰浴條件下充分研磨成勻漿后全部轉入到離心管中,于4 ℃、9 000×g離心10 min,收集上清液立即用于總酚和類黃酮的測定。取上清液分別在280 nm和325 nm波長處測定其光密度值,總酚含量用ΔOD280nm/g表示;類黃酮含量用ΔOD325nm/g表示。
1.2.4.4 木質素含量的測定
木質素含量的測定參照Yin Yan等[19]的方法。稱取3 g樣品置于預冷的研缽中,加入5 mL的經4 ℃預冷的95%乙醇,冰浴條件下充分研磨成勻漿后全部轉入到離心管中,于4 ℃、12000×g離心10 min,倒去上清液,沉淀物用95%乙醇溶液沖洗3 次,再用乙醇與正己烷的體積比為1∶2的溶液沖洗3 次,將沉淀干燥至恒質量,干燥物用1 mL 25%溴化乙酰溶液(用冰醋酸溶解),在70 ℃條件下水浴條件下孵育30 min,然后加入1 mL 2 mol/L NaOH溶液中止反應,再加入0.1 mL 7.5 mol/L羥胺鹽酸溶液和2 mL冰醋酸,然后再于4 ℃、12 000×g離心10 min,吸取上清液0.67 mL,用冰醋酸定容至10 mL,在280 nm波長處測定其光密度值,木質素含量用ΔOD280 nm/g表示。
1.3 數據統計
2 結果與分析
2.1 F. sulphureum接種對抗病/易感馬鈴薯塊莖和切片病斑直徑的影響

圖11 F. sulphureum接種對抗病/易感馬鈴薯塊莖(A)和切片(BB)病斑直徑的影響Fig.1 Effect of inoculation with F. sulphureum on lesion diameters on potato tubers (A and slices (B)
“青薯168”塊莖接種F. sulphureum后病斑擴展速度顯著小于“隴薯3號”,在整個培養過程中病斑直徑顯著小于“隴薯3號”,例如在接種后第14天和第16天,病斑直徑分別低于“隴薯3號”28.88%和29.91%。同樣,“青薯168”切片接種F. sulphureum后病斑擴展速度和病斑直徑均顯著小于“隴薯3號”,接種后第3天和第4天病斑直徑分別低于“隴薯3號”20.50%和12.65%。由此表明,“青薯168”比“隴薯3號”具有更好的干腐病抗性。
2.2 F. sulphureum接種對抗病/易感馬鈴薯塊莖切片苯丙烷代謝關鍵酶活性和相關產物積累的影響

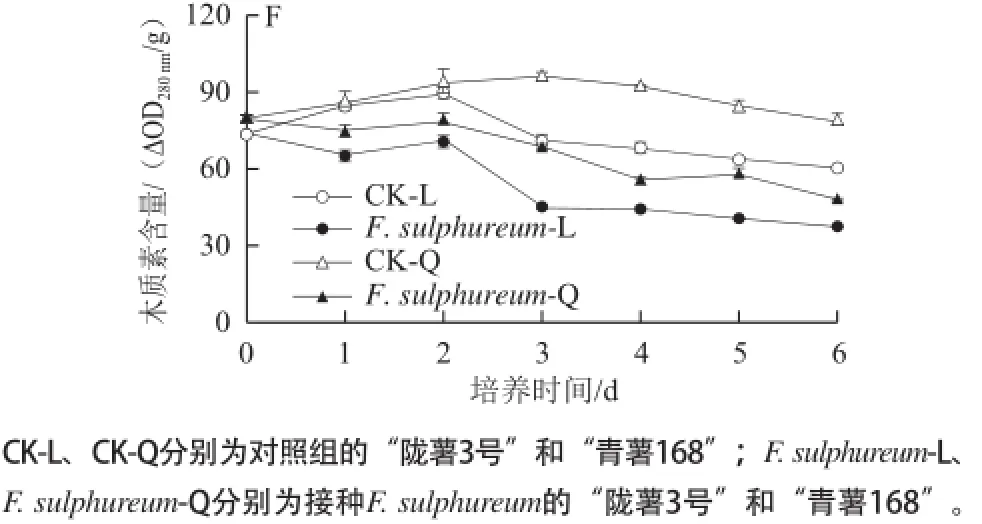
圖22 F. sulphureum 接種對抗病/易感馬鈴薯塊莖PAL(A)、4CL(B)、C4H(C)的活性和總酚(D)、類黃酮(EE)、木質素(F)含量的影響Fig.2 Changes in the activities of PAL(A)′4CL(B and C4H(C)′and the contents of total phenolics (D)′flavonoids (E and lignin (F in potato tuber slices of cv Longshu No.3 (L and cv Qingshu No.168 (Q inoculated with F. sulphureum
培養期間,2個品種塊莖切片組織中的PAL活性均呈單峰形變化,在第2天達到最大值后急劇下降,但“青薯168”的PAL活性始終高于“隴薯3號”。接種F. sulphureum后,2個品種塊莖切片組織中PAL活性在接種后的早期(1 d)均顯著增高,且“青薯168”的PAL活性上升幅度大于“隴薯3號”,之后“隴薯3號”的PAL活性持續降低,且低于未接種的對照;而“青薯168”的PAL活性則持續增高至第2天,隨后PAL活性持續降低,且于第4天后低于未接種的對照。接種后,前3 d“隴薯3號”的PAL活性不僅高于未接種的對照,也顯著高于接種的“隴薯3號”,接種后第2天和3天的活性分別高于“隴薯3號”43.39%和83.18%(圖2A)。
2個品種塊莖切片培養期間的4CL活性也呈單峰形變化,“青薯168”和“隴薯3號”分別在第3天和第4天達到最大值后有所下降,但“青薯168”的活性顯著高于“隴薯3號”,在第2天和第3天分別高出1.50、1.87 倍。接種F. sulphureum后,2個品種塊莖切片組織中的4CL活性在接種早期(2 d)均略有升高,之后“隴薯3號”的4CL活性持續降低,并低于未接種的對照;而“青薯168”的4CL活性則持續增高至第3天,之后便持續降低并低于未接種的對照,但始終顯著高于接種的“隴薯3號”,在接種后第3天和第4天的活性分別高于“隴薯3號”3.14 倍和7.12 倍(圖2B)。
2個品種塊莖切片中的C4H活性同樣呈單峰形變化,“青薯168”和“隴薯3號”的活性分別于第3天和第4天達到最大,但“青薯168”中的C4H活性始終顯著高于“隴薯3號”。接種F. sulphureum后,2個品種塊莖切片組織中的C4H活性在接種前期(3 d)也有所升高但始終低于未接種的對照,之后“隴薯3號”的C4H活性降低;而“青薯168”中的C4H活性持續升高至第4天后降低,雖低于未接種的對照,但顯著高于接種的“隴薯3號”,接種后第3天和第4天的活性分別高于“隴薯3號”26.12%和45.56%(圖2C)。
2個品種塊莖切片中總酚含量呈先上升后下降的趨勢,均在第4天達到峰值,在前3 d,兩品種中總酚含量無顯著差異,但從第4天開始,“青薯168”中總酚含量高于“隴薯3號”,在第5天時高出24.48%。接種F. sulphureum后,2個品種塊莖切片組織中總酚含量均顯著增高,于第3天達到最大值后急劇降低,“青薯168”中總酚含量上升的幅度大于“隴薯3號”,且總酚含量始終高于接種的“隴薯3號”,在接種后第3天和第4天分別高于“隴薯3號”55.01%和26.25%(圖2D)。
2個品種塊莖切片組織中類黃酮含量在前期(3 d)顯著增高,之后“隴薯3號”中類黃酮含量持續降低,而“青薯168”中類黃酮含量則持續增高至第4天,之后也略有降低,但顯著高于“隴薯3號”,在第4天和第5天分別高于“隴薯3號”34.61%和40.54%。接種F. sulphureum后,2個品種塊莖切片組織中類黃酮含量均顯著增高,于第3天達到最大值,之后迅速下降并低于未接種的對照,但“青薯168”中類黃酮含量的下降速度明顯低于接種的“隴薯3號”,類黃酮含量顯著高于接種的“隴薯3號”,在接種后第5天和第6天分別高于“隴薯3號”32.41%和32.25%(圖2E)。
2個品種塊莖切片中木質素含量呈持續下降趨勢,“青薯168”中木質素含量的下降幅度低于“隴薯3號”,且木質素含量顯著高于“隴薯3號”,在第3天和第5天分別高于“隴薯3號”36.51%和32.21%。接種F. sulphureum后,2個品種塊莖切片組織中木質素含量的下降加劇,從第1天開始,木質素含量低于對照,但“青薯168”中木質素含量的下降速度低于接種的“隴薯3號”,木質素含量顯著高于“隴薯3號”,在接種后第3天的第4天分別高于“隴薯3號”55.00%和26.15%(圖2F)。
3 討 論
本研究結果表明,“青薯168”塊莖和切片接種F. sulphureum后病斑直徑均顯著小于“隴薯3號”,表明抗病品種“青薯168”對馬鈴薯干腐病具有更強的抗性,該結果與陳紅梅等[21]的研究基本一致。接種病原物后抗病品種具有更高的抗性酶活性和更多的抗性物質積累,表明苯丙烷代謝在馬鈴薯塊莖的抗病中發揮了重要作用。
苯丙烷代謝在植物抗病防衛反應中扮演很重要的角色[7-8]。PAL是苯丙烷代謝中的關鍵酶和限速酶,其活性通常被用來衡量植物抗病性的強弱[7]。C4H是苯丙烷代謝中繼PAL之后又一關鍵酶,與香豆酸的合成密切相關,而香豆酸是合成咖啡酸、阿魏酸等物質的前體,這些物質對病原物具有直接毒殺作用[22-23]。4CL位于苯丙烷代謝的分支點處,在此酶的催化下,形成總酚、類黃酮、木質素等代謝產物[21′24]。總酚、類黃酮和木質素是植物體內最主要的抗菌物質,侵染組織周圍酚類、類黃酮和木質素的積累能有效抑制病原菌的擴展[25]。植物受到病原物侵染后苯丙烷代謝的酶類會被激活,從而積累抗性物質來抵御病原物的侵染。本研究發現,塊莖組織接種F. sulphureum后,在早期PAL和4CL活性顯著增加,該結果與Slatner等[16]的研究結果相一致。隨著酶活性的增加,總酚、類黃酮含量在接種前期也顯著高于未接種的對照,該結果與龍書生等[26]的研究一致。說明在接種前期,馬鈴薯也在一定程度上啟動了體內的抗病防衛反應。一般情況下,經物理刺激及愈傷過程中,馬鈴薯苯丙烷代謝會被刺激增強,各接菌組馬鈴薯切片苯丙烷代謝活性的增強是是愈傷過程與病原菌侵染復合影響的結果。隨著病斑的擴展,在中后期馬鈴薯組織中PAL、4CL、C4H活性迅速降低并顯著低于未接種的對照,這些酶活性的降低使合成的抗菌物質減少。接種后期總酚、類黃酮含量顯著降低并低于對照,木質素的含量一直低于未接種的對照。Saidi等[27]研究發現,海棗葉片被脆葉病菌侵染后木質素含量顯著降低,且受侵染葉片中木質素含量顯著小于健康葉片,本實驗結果與此相符。說明在接種后期或病斑的擴展期,馬鈴薯塊莖的苯丙烷代謝被抑制。
苯丙烷代謝中一些產物的含量和酶的活性均與植物的抗病性密切相關。一般認為植物體內抗菌物質含量和酶活性與抗病性呈正相關。水稻受R. solani侵染后抗病品種中木質素含量顯著高于感病品種[16]。煙草接種Alternaria alternate后,抗病品種中總酚、類黃酮含量顯著高于感病品種[28]。本研究同樣發現抗病品種“青薯168”塊莖切片組織的PAL、C4H和4CL活性在接種前期上升的幅度和持續時間顯著大于感病品種“隴薯3號”,總酚、木質素和類黃酮含量也顯著高于“隴薯3號”,說明抗病品種在受到病原物侵染后能更好地激發和維持苯丙烷代謝的活性。此外,苯丙烷代謝所產生的酚類物質及其氧化產物醌類物質可鈍化真菌的酶和毒素[31],抗病品種“青薯168”中較高的酚類物質能更好地發揮抵制病原物致病因子的毒害作用。
本課題組的前期研究表明,F. sulphureum接種馬鈴薯抗病品種后細胞壁降解酶的活性和單端孢霉烯族毒素的含量均顯著低于感病品種[29-30],這進一步驗證了上述觀點。Pseudoperonospora cubensis侵染不同品種的甜瓜以后,抗病品種中積累的木質素和胼胝質較多[31],本研究也發現接種F. sulphureum后抗病品種中木質素含量顯著高于感病品種,木質素的沉積可起到物理屏障作用。馬鈴薯愈傷過程中經常伴隨著苯丙烷代謝的增強[9],抗病品種“青薯168”較強的苯丙烷代謝的活性顯著有利于細胞表面栓質化或木質化形成愈傷周皮,從而對F. sulphureum具有較強的抵抗力。此外,研究發現“青薯168”接種F. sulphureum后活性氧清除酶類的活性高于“隴薯3號”,積累的活性氧較少,由活性氧造成的氧化損傷較少[32]。抗病品種和感病品種在組織結構上也有很大的差別。如抗病棉花品種均具有堅實的木質部,并且木質部的間隙較小,細胞壁比較厚,而感病品種的組織結構則相反[33]。植物對病原物的抗性還與體內預存抗菌物質的含量和受侵染后合成病程相關蛋白的能力有關[10]。由此推測,馬鈴薯對鐮刀菌的抗性可能還與其組織結構、預存抗菌物質和病程相關蛋白的含量相關,但還需實驗證明。
綜上所述,馬鈴薯接種F. sulphureum后,前期PAL和4CL活性以及總酚和類黃酮含量有所增加,但是隨著病原物侵染和病害的加重,罹病組織中這些酶的活性和物質的含量迅速下降并低于對照,C4H活性和木質素含量在整個培養期間均低于對照。說明苯丙烷代謝在病原物與寄主互作的早期發揮了重要作用,一旦病斑開始擴展,苯丙烷代謝的抗病作用將顯著降低。與易感病品種“隴薯3號”相比,抗病品種“青薯168”的苯丙烷代謝相關酶的活性和產物的含量顯著較高,說明抗病品種在受到病原物侵染后能更好地激發和維持苯丙烷代謝的活性,產生較多的抗菌物質。
[1] LI Yongcai BI Yang GE Yonghong et al Antifungal activity of sodium silicate on Fusarium sulphureum and its effect on dry rot of potato tubers[J]. Journal of Food Science′2009′74(5): 213-218.
[2] 李金花′柴兆祥′王蒂′等. 甘肅馬鈴薯貯藏期真菌性病害病原菌的分離鑒定[J]. 蘭州大學學報: 自然科學版′2007′43(2): 39-42.
[3] 魏周全′張廷義′杜璽. 馬鈴薯塊莖干腐病發生危害及防治[J]. 植物保護′2006′32(2): 103-105.
[4] 孫小娟′李永才′畢陽′等. 西北地區馬鈴薯貯藏期病害調查分析[J]. 中國馬鈴薯′2009′23(6): 364-365.
[5] 何蘇琴′金秀琳′魏周全′等. 甘肅省定西地區馬鈴薯塊莖干腐病病原真菌的分離鑒定[J]. 云南農業學報′2004′19(5): 550-552.
[6] 徐曉梅′楊署光. 苯丙氨酸解氨酶研究進展[J]. 安徽農業科學′2009′37(31): 15115-15119.
[7] STADNIK M J BUCHENAUER H Inhibition of phenylalanine ammonia-lyase suppresses the resistance induced by benzothiadiazole in wheat to Blumeria graminisf. sp tritici[J]. Physiological and Molecular Plant Pathology′2000′57(1): 25-34.
[8] BOURBONNAIS R PAICE M G Oxidation of non-phenolic substrates an expanded role for laccase in lignin biodegradation[J]. Febs Letters′2003′267(1): 99-102.
[9] RAMAMURTHY M S MAITI B THOMAS P High-performance liquid chromatography determination of phenolic acids in potato tubers (Solanum tuberosum during wound healing[J]. Journal of Agricultural and Food Chemistry′1992′40(4): 569-572.
[10] 夏啟中′張明菊. 植物抗病的物質代謝基礎[J]. 黃岡職業技術學院學報′2004′6(3): 38-41.
[11] KOSTYN K CZEMPLIK M KULMA A et al Genes of phenylpropanoid pathway are activated in early response to Fusarium attack in flax plants[J]. Plant Science′2012′190: 103-115.
[12] WEN K SEGUI P St-ARNAUD M et al Real-time quantitative RTPCR of defense-associated gene transcripts of Rhytophthora infestans isolates representing US-1 and US-8 genotypes[J]. Plant Pathology′2005′95(4): 1026-1037.
[13] ZHAO Changjiang WANG Airong SHI Yujun et al Identification of defense-related genes in rice responding to challenge by Rhizoctonia solani[J]. Theoretical Applied Geneicst′2008′116(4): 501-516.
[14] LEHTONEN M J SOMERVUO P VALKONEN J P T Infection with Rhizoctonia solani induces defense genes and systemic resistance in potato sprouts grown without light[J]. Phytopathology′2008′98(12): 1190-1198.
[15] 王敬文′薛應龍. 植物苯丙氨酸解氨酶的研究Ⅱ苯丙氨酸解氨酶在抗馬鈴薯晚疫病中的作用[J]. 植物生理學報′1982′8(1): 35-43.
[16] SLATNAR A MIKULIC P M HALBWIRT H et al Enzyme activity of the phenylpropanoid pathway as a response to apple scab infection[J]. Annals of Applied Biology′2010′156(3): 449-456.
[17] 龔得明′陳如凱′林彥栓. 甘蔗受黑穗病菌侵染后苯丙烷類代謝變化及與其抗性的關系[J]. 福建農業大學學報′1995′24(4): 394-398. [18] 楊志敏′畢陽′李永才′等. 馬鈴薯干腐病菌侵染過程中切片組織細胞壁降解酶的變化[J]. 中國農業科學′2012′45(1): 127-134.
[19] YIN Yan LI Yongcai BI Yang et al Postharvest treatment with β-aminobutyric acid induces resistance against dry rot caused by Fusarium sulphureum in potato tuber[J]. Agricultural Sciences in China′2010′9(9): 1372-1380.
[20] 范存斐′畢陽′王云飛′ 等. 水楊酸對厚皮甜瓜采后病害及苯丙烷代謝的影響[J]. 中國農業科學′2012′45(3): 584-589.
[21] 陳紅梅′李金花′柴兆祥′等. 35個馬鈴薯品種對鐮刀菌干腐病優勢病原的抗病性評價[J]. 植物保護學報′2012′39(4): 308-314.
[22] 李莉′趙越′馬君蘭. 苯丙氨酸代謝途徑關鍵酶: PAL、C4H、4CL研究新進展[J]. 生物信息學′2007′5(4): 187-189.
[23] 張維一′畢陽. 果蔬采后病害與控制[M]. 北京: 中國農業出版社′1996.
[24] 趙淑娟′劉滌′胡之璧. 植物4-香豆酸: 輔酶A連接酶[J]. 植物生理學通訊′2006′42(3): 529-538.
[25] REIMERS P J LEACH I E Race-specific resistance to Xanthomonas oryzae pv oryzae conferred by bacterial blight resistance gene Xa-10 in rice (Oryzae sativa involves accumulation of a lignin-like substance in host tissues[J]. Molecular Plant Pathology′1999′38: 39-55.
[26] 龍書生′李亞玲′段雙科′等. 玉米苯丙烷類次生代謝物與玉米對莖腐病抗性的關系[J]. 西北農林科技大學學報′2004′32(9): 93-96.
[27] SAIDI M N BOUAZUZ D HAMMAMI I et al Alterations in lignin content and phenylpropanoids pathway in date palm (Phoenix dactylifera L.) tissues affected by brittle leaf disease[J]. Plant Science′2013′211: 8-16.
[28] 劉敬業′陳惠明′黃學躍′等. 煙草催赤星病后苯丙烷類代謝途徑有關酶及物質的動態研究[J]. 昆明師專學報′1997′12(2): 10-14.
[29] 楊志敏. 馬鈴薯干腐病菌生物學特性及其侵染過程中塊莖細胞壁降解酶的活性變化[D]. 蘭州: 甘肅農業大學′2012.
[30] XUE Huali BI Yang TANG Yamei et al Effect of cultivars Fusarium strains and storage temperature on trichothecenes production in inoculated potato tubers[J]. Food Chemistry′2013′http://dx.doi. org/10.1016/j.foodchem.2013.11.060.
[31] COHEN Y EYAL H HANANIA J Ultrastructure autofluorescenee callose deposition and lignification in susceptible and resistant muskmelon leaves infected with the powdery mildew fungus Sphaerotheca fuliginea[J]. Physiological and Molecular Plant Pathology′1990′36: 191-204.
[32] BAO Gaihong BI Yang LI Yongcai et al Overproduction of reactive oxygen species involved in the pathogenicity of Fusarium in potato tubers[J]. Physiological and Molecular Plant Pathology′2014′86: 35-42.
[33] 陳捷胤′戴小楓. 棉花對黃萎病的抗病機制研究進展[J]. 分子植物育種′2005′3(3): 427-430.
Comparison of Phenylpropanoid Pathway Metabolism in Slices of Susceptible and Resistant Potato Cultivars Inoculated with Fusarium sulphureum
BAO Gaihong BI Yang*′LI Yongcai WANG Yi WANG Ting TANG Ying MA Chaoling BAI Xiaodong
(College of Food Science and Engineering Gansu Agricultural University Lanzhou 730070′China)
Comparative analysis between two potato cultivars was carried out to detect the key enzymes activities and metabolite accumulation in phenylpropanoid pathway in whole and sliced potatoes inoculated with Fusarium sulphureum The results showed that lesion diameters on tubers and slices of the resistant cultivar “Qingshu No.168” inoculated with F. sulphureum were significantly smaller than those of the susceptible cultivar “Longshu No.3”. The activities of phenylalanine ammonia-lyase (PAL and 4-coumarate A ligase (4CL)′and the contents of total phenolics and flavonoids in inoculated slices of both potato cultivars increased at early stages of infection but decreased sharply with the enlargement of lesions reaching levels lower than in the control at later stages of infection However the activity of cinnamate 4-hydroxylase (C4H and the content of lignin were lower than in the control over the entire culture period The activities of PAL′4CL and C4H and the contents of total phenolics flavonoids and lignin in inoculated slices of “Qingshu No.168” were higher than those of “Longshu No.3”. These findings suggested that phenylpropanoid metabolism played a positive role at early stages of F. sulphureumpotato interaction however the resistance role of phenylpropanoid metabolism decreased with the expansion of dry rot and the activities of phenylpropanoid metabolism in slices of the resistant potato cultivar were significantly higher than in the susceptible one indicating that phenylaprapanoid metabolism plays an important role in resistance of potato tubers against F. sulphureum.
potato; Fusarium sulphureum inoculation phenylpropanoid metabolism
TS201.3
A
1002-6630(2015)06-0251-06
10.7506/spkx1002-6630-201506048
2014-07-20
甘肅省科技支撐計劃項目(1011JKCA179);甘肅省自然科學基金項目(1107RJZA232);甘肅省干旱生境作物學重點實驗室開放基金項目(GSCS-2010-08)
包改紅(1987—),女,碩士研究生,研究方向為采后生物學與技術。E-mail:baogaihong@163.com
*通信作者:畢陽(1962—),男,教授,博士,研究方向為采后生物學與技術。E-mail:beyang62@163.com
