microRNA與乙型肝炎病情轉歸的關系
2015-12-14 05:24:33張秀華吳園園
分子診斷與治療雜志 2015年6期
關鍵詞:肝癌
張秀華吳園園
·綜述·
microRNA與乙型肝炎病情轉歸的關系
張秀華★吳園園
微小核糖核酸(microRNA,m iRNA)是一類內源性非編碼調控RNA,由二十幾個核苷酸長度組成,其通過抑制蛋白質翻譯或降解mRNAs調控基因轉錄后的表達,參與調節細胞的分化,增殖及凋亡,并在腫瘤的發生發展中起重要作用。研究表明乙型肝炎病毒和宿主之間通過m iRNA進行相互調節,這在HBV相關肝病的發生發展中起著非常重要的作用。本文就miRNA與HBV感染及其相關肝病的研究現狀加以綜述以探討m iRNA的研究對HBV相關肝病診治的重要意義。
miRNA;急性乙型肝炎;慢性乙型肝炎
乙型肝炎(hepatitis B,HB)是全球廣泛分布的傳染病,是我國突出的公共衛生問題之一。目前世界上約有3.5億慢性乙肝病毒(hepatitis B virus,HBV)感染者,其中中國約有1.2億人[1]。慢性HBV感染是肝癌發生的主要因素之一,我國原發性肝癌患者中HBV感染者高達80%以上,每年近20萬人死于HBV相關疾病[2]。近年來,在多種真核細胞及病毒中發現存在一類來源于內源性染色體上的非編碼單鏈RNA,即微小核糖核酸(microRNA,miRNA)。m iRNA在進化上高度保守,與靶mRNA堿基進行特異性的互補配對,通過引起靶mRNA降解或者抑制蛋白質翻譯,進而調控基因轉錄后過程,在體內發揮重要的生物學功能。多種m iRNA分子通過調控免疫細胞內的信號轉導途徑來影響機體的免疫應答[3-4]。有研究發現,在HBV感染過程中,某些m iRNAs參與了宿主的抗HBV免疫反應,并與細胞因子表達相關,其檢測可為HBV感染的慢性化進展機制提供一個新的研究線索[5-6]。因此m iRNA在HBV相關肝病表達譜的研究中具有重要意義和廣闊的應用前景。
1 miRNA的生物學特征和作用機制
m iRNA是一類由二十幾個堿基長度組成的內源性非編碼RNA分子(small non-coding RNA,sncRNA),目前在人類基因組中發現的成熟m iRNA分子已達2 000多種。m iRNA在基因組中有單拷貝、多拷貝或基因簇等多種存在形式,其位置大多落于基因間隔區,有自身的轉錄調控機制,獨立于其他基因,而且可能從前體到成熟的加工過程中,前體本身滿足了所需的所有順式序列要求。大量的miRNA與其對應的靶基因之間形成了一個龐大而又錯綜復雜的基因表達調控網絡m iRNA的調控網絡如圖1所示[7](以NF-κB信號通路為例)。
miRNA基因在核內由RNA聚合酶Ⅱ轉錄為初級m iRNA(pri-m iRNA),通過Drosha酶及蛋白DGCR8識別并剪切為長約60個~80個核苷酸(nucleotide,nt)的含發夾狀莖環結構的前體m iRNA(pre-m iRNA),再經轉運蛋白(exportin5)轉運至胞質,最后在Dicer酶的作用下,將pre-m iRNA處理成為22 nt的成熟m iRNA。成熟m iRNA 5′端有6個~8個相對恒定的核苷酸,稱為“種子序列”,miRNA通過種子序列與靶mRNA的3′非編碼區(3′-untranslated regions,3′-UTR)域配對結合抑制mRNA的降解或翻譯。根據m iRNA與靶mRNA的配對程度不同,將其作用機制分為2種:靶mRNA的切割以及翻譯抑制。切割抑制是指miRNA與靶mRNA幾乎或完全配對,靶mRNA通過RNA誘導的沉默復合體作用,直接被降解;翻譯抑制是指miRNA通過與靶mRNA的3′-UTR部分互補,同樣通過RNA誘導沉默復合體的形式來抑制轉錄后翻譯水平的基因表達,而不影響mRNA本身[8],許多動物體內miRNA的作用機制屬于后者。病毒利用在核內由RNA聚合酶Ⅱ合成的m iRNA主要有2方面的作用:(1)用于自身基因表達的調控;(2)作用于宿主細胞的基因以便于自身感染。研究表明幾種DNA病毒通過編碼m iRNA進行調節自身或作用于宿主細胞基因,有利于自身的潛伏感染,如Sullivan等發現的SV40編碼的miRNA[9]。此外,每個m iRNA可調控多個位點,而同一位點又對應多個m iRNA。miRNA的調控途徑是一個復雜的網絡,因此,依靠單個m iRNA難以完全阻斷靶mRNA的翻譯,需要若干m iRNA協同表達,聯合調控靶mRNA的翻譯功能。m iRNA的產生與作用機制如圖2所示[10]。
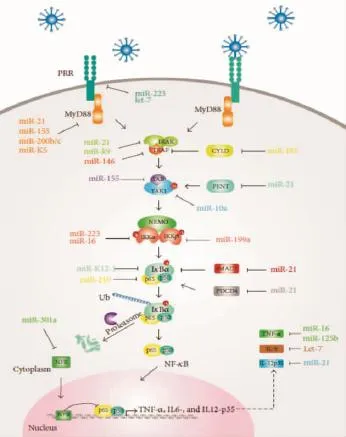
圖1 NF-κB信號通路的m iRNA調控網絡[7]Figure 1 NF-κB signal pathway network regulated by miRNAs[7]
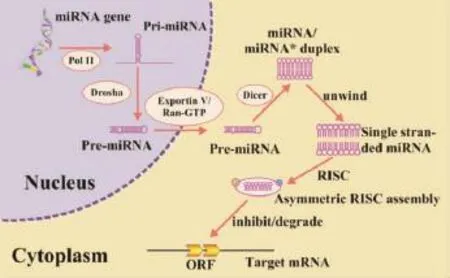
圖2 miRNA的產生與作用機制[10]Figure 2 Schematic overview of miRNA biogenesis and function[10]
2 miRNA與HBV感染及其慢性化
病毒可以利用宿主生物合成途徑,通過復雜
的機制來抵御宿主的細胞反應,在病毒-宿主細胞相互作用過程中miRNA介導了RNA沉默途徑[11]。在此過程中,病毒可以利用宿主細胞的m iRNA,也可以復制自身miRNA[12]。這說明宿主被病毒感染后,不僅宿主自身的m iRNA可作用于病毒,病毒編碼的m iRNA也可作用宿主基因,二者的相互調控對病毒性疾病的發生發展至關重要。Jin等[13]利用計算機分析HBV作為一種DNA病毒能否編碼miRNA,并利用其調節自身或宿主基因的表達,結果發現一種候選的HBV編碼的pre-m iRNA,可以靶向作用于病毒mRNA,提示HBV可能利用編碼miRNA來調節病毒自身的基因表達。來自于宿主編碼的m iRNA可能直接或間接調節病毒基因的表達。一些學者構建多種肝組織特異性m iRNA表達載體,發現宿主編碼的m iRNA可以明顯下調HBsAg和HBeAg的表達,并明顯抑制HBV的復制和表達[14-15]。miRNA與基因調控、細胞分化、疾病的發生和發展密切相關[16],患者感染HBV后,會發生急性乙肝、乙肝病毒攜帶、慢性乙肝、乙肝肝硬化、肝衰竭甚至肝細胞癌等不同階段的病理變化,miRNA在HBV感染及病情發展中發揮重要作用。
急性乙型肝炎(acute hepatitis B,AHB)患者外周血和肝臟內對HBV有強烈的多克隆和多特異性的免疫反應,這種免疫反應最終可清除HBV;而慢性HBV感染者免疫應答功能低下,從而導致HBV持續感染。在不同類型HBV感染者中miRNA分子的表達存在差異性,這可能與其慢性化機制有關。王琳等[5]在對乙型肝炎患者外周血單個核細胞中微小RNA的表達變化的研究中發現,急性乙型肝炎患者外周血單個核細胞(peripheral blood mononuclear cells,PBMC)中m iRNA155的表達水平在急性發作期明顯高于健康對照組,隨疾病進入發作期、病毒清除期、恢復期,其表達逐漸降低,而且miRNA-155與干擾素-α(interferon alpha,IFN-α)、干擾素-β(interferon beta,IFN-β)均具有良好的正相關性。m iRNA181在HBV感染者PBMC中的表達,除急性乙型肝炎恢復期外,均高于健康對照組。說明在HBV感染過程中,m iRNAs參與了宿主的抗HBV免疫應答反應,與AHB免疫發病有關。m iR-155能正性調節炎性因子的釋放[17],此外,m iR-l55還能維持CD4細胞向Thl型細胞分化促使IFN的產生。m iR-181能增強T細胞信號轉導的強度和敏感性[17]。推測在AHB的病毒清除中,m iR-l55與IFN的共同作用發揮了積極作用;而m iR-l81的表達上調,進一步增強了效應T細胞的抗病毒免疫應答作用,從而有助于宿主清除HBV。因此,由于m iR-l55缺陷所致的Th1/Th2失衡及樹突狀細胞功能抑制,可能是HBV持續感染的原因之一。
Ⅰ型干擾素及其信號通路是宿主細胞抵抗病毒的重要防御系統,患者感染HBV時,宿主內源性Ⅰ型干擾素分泌不足,并且干擾素下游的抗病毒信號轉導同樣也發生了異常,這直接導致許多HBV感染患者對于INF-α抗病毒治療應答不良。在李寧等[18]的研究中,發現m iR-548家族成員,特別是m iR-548c5p與CHB患者Ⅰ型干擾素分泌紊亂密切相關。可能是CHB發生發展中m iRNA調控的一個重要環節。此外,Liu等[19]研究結果發現HBV的復制引起肝細胞內m iR-181a的表達顯著上調,m iR-181a可靶向抑制人類白細胞抗原A的表達,從而HBV感染細胞的特異性表型減弱,使細胞毒性T淋巴細胞(cytotoxic lymphocyte,CTL)對被感染細胞的識別和殺傷能力降低,調控被感染細胞逃避宿主的免疫清除,從而參與HBV的持續感染。
在微小RNA在不同時期慢性乙型肝炎病毒感染者的異常表達譜及其臨床意義的研究中[20],與健康對照比較,人m iR-4711-3p(homo sapiens m iR-4711-3p,has-miR-4711-3p)和has-m iR-3191-5p表達水平在慢性乙型肝炎患者和HBV攜帶者明顯下調,has-m iR-5704-5p表達水平明顯上調,has-m iR-146a-5p在HBV攜帶者明顯下調,hasm iR-548ah-5P分子在慢性乙型肝炎患者中明顯上調,has-m iR-29b-3p分子在3個階段的慢性HBV感染者均明顯下調。慢性HBV感染者PBMC中m iRNA分子總體表達下調,可能與高水平的HBV抑制m iRNA分子表達,導致機體特異性免疫功能低下有關;而慢性乙型肝炎患者PBMC中miRNA分子表達明顯上調,也存在部分下調分子,可能與機體免疫功能部分恢復,而其清除功能仍不足有關;非活動性HBV攜帶者因HBV復制被抑制,m iRNA分子總體表達與健康對照差異無統計學意義。據此認為,HBV與機體相互作用導致多種
miRNA分子的異常表達,可能在維持慢性HBV感染者免疫穩態機制中發揮重要作用。
3 miRNA與慢性HBV感染的轉歸
m iRNA異常表達與慢性肝炎、肝硬化及向肝癌演變的過程密切相關。馬兆龍等[21]利用m iRNA芯片技術,分析了在HBV感染、慢性肝炎肝硬化及肝癌演變過程中m iRNA表達譜的變化。結果表明肝硬化和肝癌組織與正常組織相比,均發現有表達上調和下調的m iRNA,并且這些與正常組織有差異表達的m iRNA在肝硬化和肝癌組織中幾乎沒有差別。提示在不同類型肝臟組織中,在HBV感染→肝硬化→肝癌的發生發展過程中差異性表達的m iRNA可能發生在進程早期,起到重要的類似癌基因或抑癌基因的作用,相關m iRNA表達的改變可能促進了該進程的發生發展。其他實驗結果則表明miR-21是抑癌基因PTEN的上游調控因子,調節m iR-21能引起PTEN的改變,繼而調控Akt信號途徑和細胞的增殖活性[22]。有的m iRNA則可能作為肝癌預防和早期診斷的潛在標志物,比如m iR-200a就極大程度地參與了肝癌的發生,在肝癌發生的各個階段均有巨大的指示作用[23]。而miR-148a在體外能抑制肝癌細胞SMMC-7721的侵襲和遷移,進一步明確m iR-148a在肝癌侵襲轉移中的分子機制,從而為肝癌的治療提供實驗依據[24]。miR-122是肝臟中特異性高表達的m iRNA,但在肝癌組織中表達低,且表達水平隨著腫瘤分化程度的降低而下調,這種改變促進肝癌細胞的分裂和增殖,可將其作為未來肝癌診斷和預測的指標[25]。構建miR-122慢病毒表達質粒并感染肝癌細胞,穩定表達的m iR-122a能有效抑制肝癌細胞增殖及克隆增生,證實m iR-122在肝癌生長侵襲方面發揮著重要作用[26]。研究發現血清m iR-222在肝癌患者表達顯著上調,其高表達與肝硬化、腫瘤數目、門靜脈癌栓、TNM分期相關,可能有助于肝癌患者的預后預測[27]。Li等[28]驗證了肝癌患者體內m iRNA-139與臨床特征的相關性,血漿miRNA-139的表達水平可以作為HCC的診斷標志物和預后因素。上述研究表明miRNA在癌旁組織和正常肝組織表達存在差異。但是可能是由于采用的研究方法不同,許多研究發現雖然HCC中miRNA表達失調,但是結論并不一致,應注意因分析方法不同所致的差異。
4 展望
m iRNA在HBV相關肝病的發病機制中的意義重大,隨著研究的不斷深入,特別是m iRNA芯片技術的發展,它為疾病的診治帶來全新的觀念;從診斷方面來看,m iRNA為肝臟疾病的診斷提供了大量信息,m iRNA作為早期診斷HCC及病毒性肝炎感染的新型標志物,將極大程度的提高病毒性肝炎、肝纖維化、肝硬化和HCC等肝臟疾病的診斷水平;從疾病治療的角度分析,隨著對循環m iRNA的深入研究及檢測方法的進步,將更加清晰地闡明許多疾病的發病機制,并為疾病的治療提供新的思路。miRNA介導的RNAi作為一種新型的基因治療技術,具有良好的應用前景。
[1]李晨,萬謨彬,王慧芬.m icroRNA在HBV相關肝病中的研究進展[J].胃腸病學和肝病學雜志,2010,19 (8):774-776.
[2]Neuveut C,Wei Y,Buendia MA.Mechanisms of HBV -related hepatocarcinogenesis[J].J Hepatol,2010,52 (4):594-604.
[3]Li QJ,Chau J,Ebert PJ,et a1.miR-181a is an intrinsic modulator of T cell sensitivity and selection[J].Cell, 2007,129(1):147-161.
[4]Lu LF,Boldin MP,Chaudhry A,et a1.Function of m iR-146a in controlling Treg cell-mediated regulation of Thl responses[J].Cell,2010,142(6):914-929.
[5]王琳,謝青,龔邦東,等.乙型肝炎患者外周血單個核細胞中微小RNA的表達變化[J].中華傳染病雜志, 2010,28(8):473-479.
[6]張靜,史進方,高純,等.乙型肝炎病毒感染不同時期外周血單個核細胞微小RNA表達變化及其意義[J].中華檢驗醫學雜志,2012,35(6):544-549.
[7]Gao Z,Dou Y,Chen Y.M icroRNA roles in the NF-κB signaling pathway during viral infections[J]. Biomed Research International,2014,doi:10.1155/ 2014/436097.
[8]Bartel DP.M icroRNAs:genom ics,biogenesis,mechanism,and function[J].Cell,2004,116:281-297.
[9]Sullivan CS,Grundhoff AT,Tevethia S,et al.SV40-encoded m icroRNAs regulate viral gene expression and reduce susceptibility to cytotoxic T cells[J].Nature, 2005,435(7042):682-686.
[10]Liu B,Fang L,Wang X,et al.Identification of realm icroRNA precursors w ith a pseudo structure status composition approach[J].Plos One,2015,10(3):e0121501.
[11]Gottwein E,Cullen BR.Viral and celIular micmRNAs as detem inants of viral pathogenesis and immunity[J]. Cell HostM icmbe,2008,3(6):375-387.
[12]SkalBky RL,samols MA,Plaisance KB,et a1.Kaposi’s sarcoma-as-soiated herpesvirus encodes an ortholog of miR-155[J].JVim I,2007,8l(23):12836-12845.
[13]Jin WB,Wu FL,Kong D,et a1.HBV-encoded m icroRNA candidate and its target[J].Comput Biol Chem, 2007,31(2):124-126.
[14]Gao YF,Yu L,WeiW,et al.Inhibition of hepatitis B virus gene expression and replication by artificial microRNA[J].World J A stroenterol,2008,14(29):4684-4689.
[15]羅祥基,程慶保,徐峰,等.慢病毒介導基于microrna系統的hbsrnai技術抑制hbv復制[J].第二軍醫大學學報, 2009,30(3):295-299.
[16]Chen XM.M icroRNA signatures in liver diseases[J]. World JGastroenterol,2009,15(14):1665-1672.
[17]Tili E,M ichaille JJ,Cim ino A,etal.Modulation of m iR-155 and miR-125b levels follow ing lipopolysaccharide/TNF-alpha stimulation and their possible roles in regulating the response to endotoxin shock[J].Immunology,2007,179(8):5082-5089.
[18]李寧,陳明泉,程琦,等.調控慢性乙型肝炎患者Ⅰ型干擾素信號通路的m icroRNA的篩選及鑒定[J].中國病毒病雜志,2012,2:107-112.
[19]Liu Y,Zhao JJ,Wang CM,et al.Altered expression profiles of m icroRNAs in a stable hepatitis B virus-expressing cell line[J].Chin Med J,2009,122(1):10-14.
[20]邢同京,徐洪濤,余文慶,等.微小RNA在不同時期慢性乙型肝炎病毒感染者的異常表達譜及其臨床意義[J].中華傳染病雜志,2013,31(10)586-592.
[21]馬兆龍,楊煉,陳立波,等.m iRNA在HBV從感染經由肝硬化到肝癌進程中表達譜的變化[J].世界華人消化雜志,2009,17(20):2112-2116.
[22]Zhang Y,Jia S,Xiang F,et al.Expression ofmiR-122 in hepatocarcinoma cell strains and its relationship w ith PTEN[J].Chinese Clinical Oncology,2014,19(6):490-493.
[23]夏寧,高媛,王興,等.血清及肝組織miR-200a在大鼠肝癌發生過程中的動態表達分析[J].中華肝臟病雜, 2014,22(6):440-444.
[24]賈筱琴,繆俊俊,雍軍,等.miR-148a對肝癌細胞株侵襲和遷移的抑制作用及機制[J].中國癌癥雜志,2014, 24(6):412-417.
[25]張纓,賈紹昌,項方,等.miR-122在肝癌細胞中的表達及其與細胞周期調控的關系[J].臨床腫瘤學雜志, 2013,18(8):691-694.
[26]莊鵬,李志瑩,王湘郴,等.m iR-122慢病毒載體的構建及其在肝癌細胞中的作用[J].2013,29(7):529-531.
[27]占美曉,李勇,胡寶山,等.肝癌患者血清中微小RNA的表達及其臨床意義[J].中華醫學雜志,2013,93(23): 1830-1832.
[28]Li T,Yin J,Yuan L,et a1.Downregulation ofmicroRNA-139 is associated w ith hepatocellular carcinoma risk and short-term survival[J].Oncol Rep,2014,31(4):1699-1706.
The relationship between m icro RNA and prognosis of hepatitis B
ZHANG Xiuhua★,WU Yuanyuan
(Clinical Laboratory of Heze Municipal Hospital,Heze,Shandong,China,274031)
M icroRNA(miRNA)is a type of endogenous regulation of non-coding RNA which is made up of more than 20 nucleotides in length.M iRNA involves in the regulation of cell differentiation, proliferation and apoptosis by inhibiting the expression of protein translation or degrading mRNAs to regulate gene transcription.It plays an important role in the occurrence and development of cancer.Studies have shown that the hepatitis B virus(HBV)and its host carry outmutual adjustment through m iRNA.It is very important in the occurrence and development of HBV-related liver disease.In this paper,the research status of miRNA and HBV infection w ill be reviewed to discuss the importance ofmiRNA on diagnosis and treatment of HBV-related liver disease.
m iRNA;Acute hepatitis B;Chronic hepatitis B
國家科技重大專項(2012ZX10002005)
山東省菏澤市立醫院檢驗科,山東,菏澤274031
★通訊作者:張秀華,E-mail:zxhhzslyy@163.com
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24