七姊妹山保護區紅椿種群動態與譜分析
2015-12-18 19:27:35汪洋張敏陳霞周火明操英南肖創偉梁國章李新枝
湖北農業科學 2015年22期
汪洋 張敏 陳霞 周火明 操英南 肖創偉 梁國章 李新枝
摘要:為研究紅椿(Toona ciliata Roem)種群動態,將林木徑級代表年齡結構,分析了齡級與種群數量動態,編制了靜態生命表, 繪制并分析了存活曲線、死亡率曲線和種群損失度曲線,對種群進行了譜分析。結果表明,七姊妹山紅椿種群衰退動態指數Vpi=32.32%,穩定動態指數Vpi′=4.039%,屬于增長型穩定種群,風險敏感度較高。存活曲線接近DeeveyⅡ型,各齡級個體具有大體相等的死亡率。死亡率qx與損失率Kx均在1、4、6級出現高峰,說明不同階段紅椿生理特性、環境篩與人為干擾因素對種群的影響較大。波譜分析基波A1=0.752 8,A3出現明顯小周期,表明紅椿種群天然更新存在著周期性,且有小周期的多諧波疊加。
關鍵詞:紅椿(Toona ciliata Roem);種群動態;譜分析;七姊妹山
中圖分類號:Q949.753.1;S758.5 文獻標識碼:A 文章編號:0439-8114(2015)22-5632-05
Abstract: To research the dynamics of Toona ciliata Roem population in Qizimeishan Mountain, diameter classes of T. ciliata plants were transformed into according age classes; numeral dynamics of the plants in different age classes were analyzed; a static life table was made; curves of survival rate, death rate and vanishing index were drawn; and a spectral analysis of the population was conducted. The research indicates that the population is in growth form, as dynamics index Vpi was 32.32%; and Vpi′ was 4.039% when stress considered; and the sensitiveness value was comparatively high. The survival curve conforms approximately to that of Deevey Ⅱ, illustrating a general equivalence of death rate in different age classes. qx and Kx surge to peaks in 1, 4 and 6 age classes simultaneously, showing that the physiological features at different growing stages, environmental sieves and human disturbances all influence the T. ciliata population. In spectral analysis of the population, the fundamental wave A1 is 0.752 8, while A3 has an obvious minor cycle deviating from that of A1, demonstrating a periodicity in natural regeneration of T. ciliata population, with multi-harmonic wave superposition of minor cycles.
Key words: Toona ciliata Roem; population dynamics; spectral analysis; Qizimenshan Mountains
植物種群動態是一定時間和空間范圍內種群的大小與數量的變化規律,是植物個體生存能力與環境相互作用的結果,現已成為植物生態學研究的熱點之一[1]。生命表可以預測出該物種在某些特定條件下存活與繁殖的可能性,了解種群的現存狀態,分析過去種群的結構與受干擾狀況,并可預測未來的種群動態[2]。Fourier譜分析是探討種群天然分布的波動性和年齡更替過程周期性的數學工具,種群天然更新動態可通過種群不同齡級的株數分布波動來表現[3,4]。伍業鋼等[3]認為紅松(Pinus koraiensis Siebold et Zuccarini)天然更新過程呈周期波浪式發展,這也是其穩定的一個特點。波動可出現于所有的植被中,而譜分析方法可以揭示種群數量變動的周期波動[5]。因此,植物種群的動態研究和譜分析對于研究種群,尤其是瀕危優勢種的生命過程就具有現實意義。
紅椿(Toona ciliata Roem)為楝科(Meliaceae)落葉大喬木。素有“中國桃花心木”之稱,被列為Ⅱ級國家重點保護野生植物[6]。湖北省的恩施地區有零星分布,由于開發過度、自然環境變化和天然更新緩慢等原因,分布區逐漸縮小。課題組通過對湖北省內的紅椿種群實地考察,探討分布地七姊妹山紅椿種群的生長狀況和動態規律,旨在為紅椿種群的保護與管理提供理論依據。
1 研究區概況
研究區位于七姊妹山國家級自然保護區。地處鄂西南武陵山區的宣恩縣境內。該地位于北緯29°39′30″-30°05′15″,東經109°38′30″-109°47′00″,總面積34 550 hm2,由七姊妹山、秦家大山和八大公山3個大的山脊構成。地表的巖層主要由石英砂頁巖、頁質層巖、沙質層巖組成[7];土壤分布類型隨海拔高度變化而不同,海拔1 500 m以下的區域為黃棕壤。在氣候條件方面屬中亞熱帶季風濕潤型氣候;海拔800~1 200 m的亞高山地帶年平均氣溫13.7 ℃,年無霜期263 d,年降水量1 635.3 mm[8],年日照時間1 212.4 h。植被以常綠、落葉闊葉混交林為主,具有亞熱帶山地的典型植被類型[9]。
雖然紅椿在保護區范圍外西部與沙道溝鎮相連地段有零星分布,且具有極少數胸徑達到100 cm 以上的古樹,但沒有完整的種群格局,不便于取樣研究。樣地位于長潭鄉后河村賀家灣,該村地處北緯30°02′34″,東經109°41′30″,是宣恩縣發現最完整、面積最大的種群,其紅椿為群落優勢種。種群位于山溝槽,土壤為山地黃棕壤,陽光充足。由于人為干擾嚴重,物種豐富度很低。主要喬木包含棕櫚[Trachycarpus fortunei(Hook. f.)H.Wendl.]、櫻桃[Cerasus pseudocerasus (Lindl.)G. Don]等7種。灌木藤本有八角楓[Alangium platanifolium(Siebold et Zucc.)Harms]、柔毛繡球(Hydrangea villosa Rehd)、中國旌節花(Stachyurus chinensis Franch.)等11種。草本蕨類豐富,有大葉水龍骨[Goniophlebium niponicum(Mett.)Bedd. var. wattii(Bedd.)Bedd.]、卵心葉虎耳草(Saxifraga aculeate Baif f.)、管莖鳳仙花(Impatiens tubulosa Hemsl.)等36種。
2 研究方法
2.1 野外調查方法
受環境因素限制,七姊妹山紅椿種群是課題組在宣恩縣發現的最大且較完整的天然種群。試驗選擇的樣地面積均為40 m×10 m,能完全覆蓋全部種群。在每塊樣地內設5 m×5 m的樣方,共計16個。對樣方內紅椿進行每木調查,統計數據。對胸徑(DBH)≤2.5 cm的植株測定地徑、高度;對胸徑≥2.5 cm的植株記錄樹高、胸徑、冠幅、枝下高、分枝角等指標。記錄環境因子,對林下的喬灌草藤本植物進行調查統計,具體見表1。
2.2 齡級劃分方法及動態分析
為維護保護區生態完整性,研究參照戴其生等[10]對紅椿用材林的解析數據和龍漢利等[11]對紅椿胸徑與年齡的擬合公式。通過紅椿平均胸徑與連年生長過程分析,采用“空間替代時間法”,即以立木級結構代替種群年齡結構分析種群動態[12,13],以每4年為1個齡級,將該種群劃分為8個齡級。即幼苗級Ⅰ(DBH<2.5 cm);從第二齡級開始,以胸徑5 cm為步長增加一級幼樹級Ⅱ(2.5 cm≤DBH<7.5 cm)、Ⅲ(7.5 cm≤DBH<12.5 cm)、Ⅳ(12.5 cm≤DBH<17.5 cm);中樹級Ⅴ(17.5≤DBH<22.5 cm),Ⅵ(22.5 cm≤DBH<27.5 cm);大樹級Ⅶ(27.5 cm≤DBH<32.5 cm),Ⅷ(DBH≥32.5 cm)。
采用陳曉德[14]的數學模型,推導種群年齡結構的動態指數(V),對紅椿種群動態進行定量描述,Vn、Vpi、Vpi′分別取正、負、零值的意義,分別反映種群或相鄰年齡級個體數量的增長、衰退、穩定的動態關系。
2.3 編制生命表
編制紅椿種群靜態(特定時間內)生命表。靜態生命表包含以下項目:x為齡級內徑級值;ax為在x級現有的個體數;lx為在x級開始時標準化存活的個體數(一般轉換為1 000);dx為從x到x+1齡級間隔期內標準化死亡的個體數;qx為從x到x+1齡級間隔期間的死亡率;Lx為從x到x+1齡級間隔期間還存活的個體數;Tx為從x齡級到超過x齡級的個體總數;ex為進入x齡級個體的生命期望或平均期望壽命;Kx為損失度[12,15,16]。
2.4 種群譜分析
譜分析是以Fourier級數展開的,寫成正弦波形式:
式中,A0為周期變化的平均數,Ak為各諧波振幅,標志其起作用的大小;ωk和θk分別為諧波率及相角;Nt為t時刻種群的大小。將種群各年齡個體數分布視為一時間系列t,以xt表示為t年齡序列時個體數。試驗采用前面劃分的徑級序列作為其年齡序列。n為系列總長度,P=2/n為諧波的總個數。T為正弦波的基本周期,即時間系列t的最長周期,也就是資料總長度,這里T=n是已知的。則可利用下式來估計Fourier分解后的各個參數[3,4]:
3 結果與分析
3.1 齡級結構與動態
試驗在16個樣方中共記錄到大小植株41株。以8個年齡級為橫坐標,以各齡級個體存活數量為縱坐標,繪制紅椿種群徑級結構的分布圖,結果見圖1。由圖1可以看出,Ⅰ齡級的幼苗級紅椿植株為15株,占比達36.59%,中間經歷了Ⅳ、Ⅴ、Ⅵ 3個齡級,個體數量形成了一個谷區。最小存活數反映在Ⅴ齡級上,只占2.44%。Ⅶ齡級占12.20%。顯而易見,圖1直觀表現出了種群中不同齡級種群數量的大小。
對紅椿種群各齡級數量變化動態進行定量分析,發現Ⅰ、Ⅱ、Ⅲ、Ⅳ齡級的個體數量變化動態指數分別為40.00%、55.56%、50.00%、50.00%,處于正增長。Ⅴ、Ⅵ、Ⅶ齡級的個體數量變化動態指數分別為-50.00%、-33.33%、-40.00%,為負增長。反映到齡級結構圖上一致。整個種群年齡結構的數量變化動態指數Vpi=32.32%,>0,種群屬于增長型[17];Vpi′=4.039%,>0,說明種群仍穩定;P=0.125,較大,表明種群的抗風險敏感性很高。
3.2 生命表與存活曲線
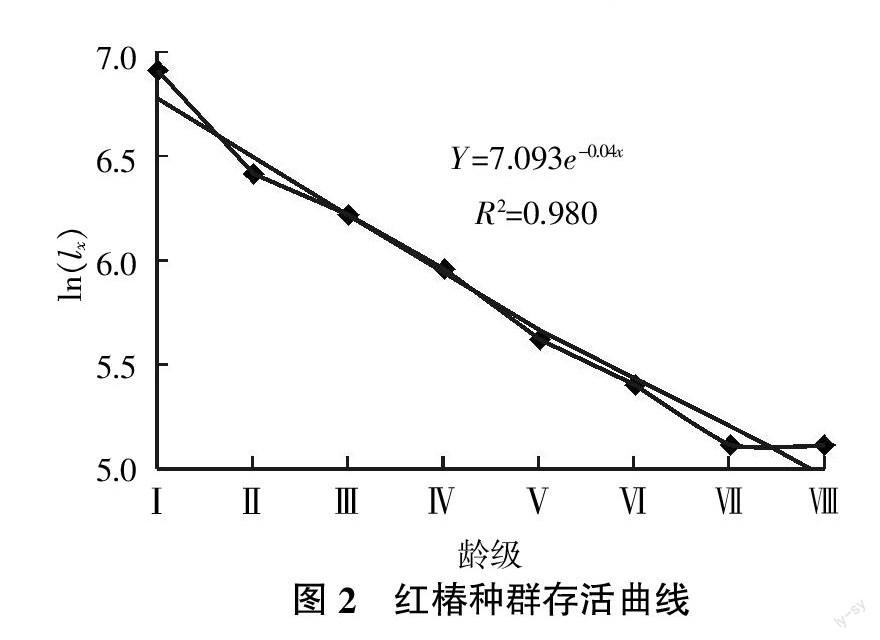
年齡結構與靜態生命表以及種群動態預測都是基于實際數據而做出的,能夠反映種群的基本屬性,結果也符合種群的自然動態規律[5,18]。試驗做出的七姊妹山保護區紅椿種群生命表見表2。在該生命表中,Ⅴ~Ⅵ齡級出現負的dx值,與生命表數學假設技術不符。因此,在生命表的編制中,許多學者采用了Lowe對赤鹿種群表使用的勻滑技術(Smooth out)[12]進行處理,本研究也采用此技術對紅椿種群進行勻滑處理,處理中的ax為ax′。
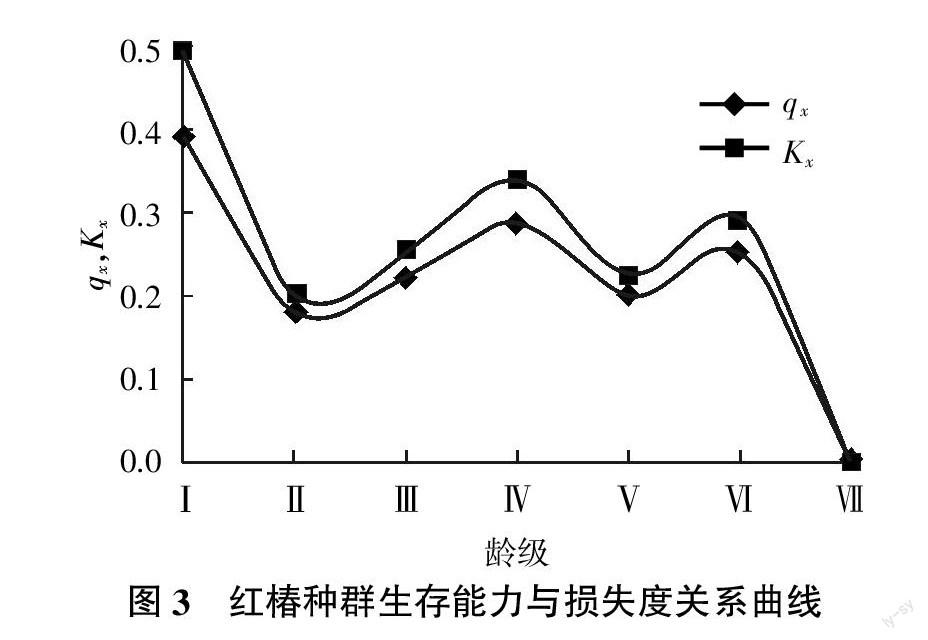
種群生命表和存活曲線從整體上反映了種群的數量動態變化趨勢[15]。從表2可知,qx的Ⅰ級與Ⅳ級最高,相對幼苗數量大,Ⅶ級為0,與Kx的解釋一致。ex在Ⅱ齡級最高,相應的,前面5個齡級(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ)的值高于后面的3個齡級,說明生命活力在幼苗到成樹初期較高,種群生命力較強。Kx直觀體現在Ⅰ和Ⅳ 2個齡級,最大值在Ⅰ齡級,為0.492。相應幼苗級到幼樹級損失度最大。Ⅶ齡級損失為0,表明種群進入動態穩定期。
存活曲線是反映種群個體在各齡級的存活狀況曲線,是借助于存活個體數量來描述特定年齡死亡率的[19]。紅椿種群存活曲線見圖2。由圖2可知,在Ⅰ~Ⅱ齡級與Ⅵ~Ⅶ齡級區間種群死亡率最高,幼苗數量最多,種群發育過程中逐漸受到環境篩的限制。經過兩個波動的高死亡率,形成數量較為穩定的高齡級成樹。這種現象的成因與紅椿的生理特性有關,除幼苗時期外,整個發育過程必須保證強光才能實現正常發育。因此種群發育過程必然缺失中間齡級。存活曲線檢驗符合DeeveyⅡ型曲線[20]。
3.3 死亡率與損失度分析
種群死亡率曲線qx反映種群死亡率的動態變化過程。消失率曲線Kx反映各齡級的致死力或損失度。紅椿種群生存能力與損失度關系曲線見圖3。從圖3可知,紅椿種群死亡率曲線與消失率曲線基本一致,即在Ⅰ~Ⅱ齡級區間、Ⅵ~Ⅶ齡級區間下降趨勢一致。Ⅰ齡級死亡率最大,其次Ⅳ齡級經歷第二次高峰。表明此時環境篩選強度很高,主要體現在光因子和種間種內競爭方面,人為干擾也是重要因素。相應Kx的變化也在Ⅰ、Ⅳ、Ⅵ齡級出現三次高峰。Ⅱ、Ⅴ齡級種群表現相對穩定,死亡率和消失率較低。Ⅰ齡級的高qx與Kx說明可能是物種的生理特性,而Ⅳ、Ⅵ齡級的小波峰說明種群與樹齡,土壤、空間和水分等生態因子對其生存的限制有關外,更多與人類的干擾關系緊密。
3.4 紅椿種群譜分析
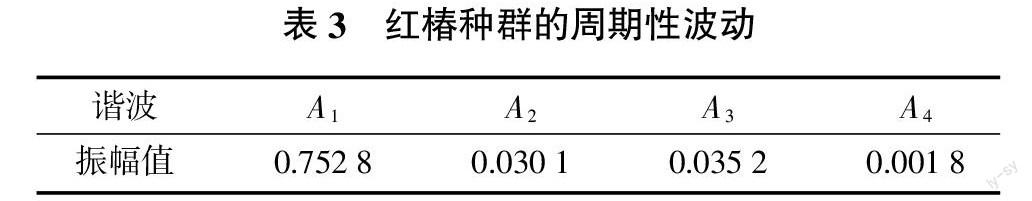
伍業鋼等[3]研究紅松種群后認為,各世代所形成的一定時間間隔的規律性就是紅松種群天然更新過程的周期性反映,群落優勢種存在一定的共性。本研究通過不同齡級與存活數量分布波動來探討紅椿種群的動態,以生命表的齡級作為波譜分析的級差,共有8級;采用未經勻滑處理的實際數據,總波序K=N/2=4。利用波譜分析公式計算各個波形的振幅Ax值(K=1,2,……,P),結果見表3。在表3中, A1為基波,A2~A4為各個諧波,Ax的值大小差異反映了各周期作用大小的差別。基波A1=0.752 8,為種群本身所固有,反映種群波動特性。紅椿種群數量動態除受基波影響外,還顯示出明顯的小周期波動。A3=0.035 2,出現明顯小周期,反映在8×5/3=13 cm徑級,即Ⅳ齡級。前面生命表里的Ⅳ齡級的高死亡率和高損失度也與這一現象契合。
紅椿種群受調查面積、年齡的限制,不能完整反映種群的全部生命周期。但譜分析表明,紅椿種群天然更新存在著周期性,而且是非單一周期,大周期內有小周期的多諧波迭加特征。小周期波動約為12~16 a(A3),是理論上的最小周期,其形成機制有待研究。這種波動與生命表特征吻合,小的周期波動可使種群的自我穩定性得以維持與延續[5]。
4 小結與討論
紅椿是強陽性植物,因此紅椿的分布多在陽坡或靠近河道光照條件好的地方。在幼苗發育成幼樹的過程中受到環境篩的強烈篩選,達到Ⅳ齡級的植株往往缺乏光照,不能維持自身的碳素平衡,在進入主層林之前通常死亡。生命表分析表明,七姊妹山紅椿種群存活曲線屬于DeeveyⅡ型,生命過程各階段具有相同死亡率。但對恩施市盛家壩紅椿種群研究得出的結論卻是符合Deevey Ⅲ型。胡喜生等[18]通過木荷(Schima superba Gardn. et Champ.)天然種群的研究發現珍稀瀕危植物的存活曲線不是單一的,說明引起瀕危的原因很復雜。如木荷、米櫧[Castanopsis carlesii(Hemsl.)Hay.]、南方紅豆杉[Taxus chinensis(Pilger)Rehd. var. mairei (Lemee et Levl.)Cheng et L. K. Fu]和冷杉[Abies fabri(Mast.)Craib]等屬于Deevey Ⅲ型;而絲栗栲(Castanopsis fargesii Franch.)、南方鐵杉[Tsuga chinensis(Franch.)Pritz. var. tchekiangensis(Flous)Cheng et L.K.Fu]和長苞鐵杉(Tsuga longibracteata W. C. Cheng)等存活曲線屬于DeeveyⅡ型[18]。
種群的死亡和生存率是種群內在的變化與極端環境條件相互作用的結果[21]。七姊妹山種群位于天然林與農田的交界地帶,可能是由于人為干擾因素過多、特定環境因子影響了種群結構造成的。紅椿不同種源地出現兩種類型存活曲線其原因是復雜的,有待進一步探討。
紅椿種群出現三次死亡率和損失度峰值,說明了紅椿幼苗到幼樹只有部分獲得“林窗機遇”,其以高死亡率為代價進入幼樹期。第二階段的高峰來自林冠郁閉、種內種間競爭加劇,出現新死亡率峰值。第三次高峰值可能來自于年齡或生理等因素。
波動出現于所有植被中,周期性波動可成為種群穩定性維持的一個機制,而譜分析方法可用于表述這種機制[4]。受調查資源的限制,群落環境、生理特性和人為干擾等因素會對周期長短產生影響,紅椿種群天然更新存在著周期性是肯定的,而且是非單一性的周期,并且具有大周期內有小周期的多諧波迭加特征。小周期波動為12~16 a,說明紅椿種群天然更新過程的周期波動與生長動態是密切相關的。可能在同一周期內,紅椿的聚集分布(另文研究)將加劇種內競爭或抑制光環境,而Ⅳ齡級的高死亡率又騰出了空間,促進了優勢者生長;另有可能是同一周期內個體生長受到前后不同周期波動的影響,其中原因需要進一步探討。
參考文獻:
[1] 李 妍,李登武,韓東辰.賀蘭山東坡青海云杉種群動態[J].浙江農林大學學報,2014,31(1):50-56.
[2] STEWART G H, ROSE A B. The significance of life history strategies in the developmental history of mixed beech(Notho fagus) forest, New Zealand[J]. Vegetation,1990,87(2):101-114.
[3] 伍業鋼,薛進軒.紅松林紅松種群動態的譜分析[J].生態學雜志,1988,7(1):19-23.
[4] 劉金福,洪 偉,吳則焰,等.孑遺植物水松(Glyptostrobus pensilis)種群生命表和譜分析[J].武漢植物學研究,2008,26(3):259-263.
[5] 閆淑君,洪 偉,吳承禎,等.絲栗栲種群生命過程及譜分析[J].應用與環境生物學報,2002,8(4):351-355.
[6] 丁永福.中國野生植物保護工作的里程碑——國家重點保護野生植物名錄(第一批)出臺[J].植物雜志,1999(5):5-11.
[7] 劉勝祥,瞿建平.湖北七姊妹山自然保護區科學考察與研究報告[M].武漢:湖北科學技術出版社,2006.l-22.
[8] 馬宇飛,李俊清.湖北七姊妹山珙桐種群結構研究[J].北京林業大學學報,2005,27(3):12-16.
[9] 班繼德,漆根深,李 博,等.鄂西植被研究[M].武漢:華中理工大學出版社,1995.30-138.
[10] 戴其生,張梅林,徐玉偉,等.紅楝子人工造林試驗初報[J].安徽林業科技,1997,34(1):33-34.
[11] 龍漢利,馮 毅,向 青,等.四川盆周山地紅椿生長特性研究[J].四川林業科技,2011,32(3):37-41.
[12] 江 洪.云杉種群生態學[M].北京:中國林業出版社,1992.8-26.
[13] 呂海英,王孝安,李 進,等.珍稀植物銀砂槐中國分布區的種群結構與動態分析[J].西北植物學報,2014,34(1):177-183.
[14] 陳曉德.植物種群與群落結構動態量化分析方法研究[J].生態學報,1998,18(2):214-217.
[15] 洪 偉,王新功,吳承禎,等.瀕危植物南方紅豆杉種群生命表及譜分析[J].應用生態學報,2004,15(6):1109-1112.
[16] 吳承禎,洪 偉,謝金壽.珍稀瀕危植物長苞鐵杉種群生命表分析[J].應用生態學報,2000,11(3):333-336.
[17] 楊 寧,鄒冬生,楊滿元,等.貴州雷公山禿杉的種群結構和空間分布格局[J].西北植物學報,2011,31(10):2100-2105.
[18] 胡喜生,洪 偉,吳承禎,等.木荷天然種群生命表分析[J].廣西植物,2007,27(3):469-474.
[19] 劉 洋.小興安嶺山楊種群生命表及生存分析[J].林業科技,2010,35(2):15-18.
[20] 黃 剛,杜珍珠,孫 肖,等.準噶爾盆地南緣小葉樺種群生命表與生存分析[J].江蘇農業科學,2012,40(9):310-313.
[21] 吳承禎,洪 偉.杉木數量經營學引論[M].北京:中國林業出版社,2000.129-132.
(責任編輯 王珞)
