芹菜素激活c-Src/NADPH氧化酶/ROS通路誘導HO-1表達從而抑制小鼠巨噬細胞過度分泌細胞因子
2015-12-28 00:55:15,,,,,,,,*
中南醫(yī)學科學雜志 2015年4期
, , ,,,,,,*
(1.南華大學附屬第二醫(yī)院呼吸內(nèi)科;2.南華大學附屬第二醫(yī)院藥劑科;3.南華大學病原生物學研究所;湖南 衡陽 421001)
·基礎醫(yī)學·
芹菜素激活c-Src/NADPH氧化酶/ROS通路誘導HO-1表達從而抑制小鼠巨噬細胞過度分泌細胞因子
曾賽麗1,吳廣2,謝莉1,張秀峰1,譚小武1,何振華1,游曉星3,趙蘭華3,曾焱華3*
(1.南華大學附屬第二醫(yī)院呼吸內(nèi)科;2.南華大學附屬第二醫(yī)院藥劑科;3.南華大學病原生物學研究所;湖南 衡陽 421001)
目的觀察芹菜素對脂多糖(LPS)誘導小鼠巨噬細胞分泌細胞因子的影響,并探討其分子機制。方法體外培養(yǎng)小鼠巨噬細胞RAW 264.7,用不同濃度的芹菜素處理后,加入LPS刺激0~16 h。Western blot檢測血紅素氧合酶-1(HO-1)的表達以及c-Src和NADPH氧化酶p47phox亞基的磷酸化;熒光探針H2DCFDA檢測活性氧(ROS)的產(chǎn)生;ELISA檢測TNF-α和IL-6的產(chǎn)生。結(jié)果芹菜素作用RAW 264.7細胞30 min即可誘導c-Src和NADPH氧化酶亞基p47phox磷酸化。采用50 μmol/L c-Src抑制劑PP1預處理細胞后,p47phox磷酸化水平明顯降低。同時,芹菜素能上調(diào)RAW264.7細胞內(nèi)ROS的含量,而NADPH氧化酶抑制劑DPI可抑制其產(chǎn)生。芹菜素也能誘導RAW264.7細胞表達HO-1,而采用PP1、DPI或ROS抑制劑N-乙酰半胱氨酸(NAC)預處理細胞后,HO-1表達水平明顯降低。此外,芹菜素處理能下調(diào)LPS誘導RAW264.7細胞分泌TNF-α和IL-6,而采用siRNA沉默HO-1表達后,能在一定程度上降低芹菜素對細胞因子的抑制效應。結(jié)論芹菜素可能通過激活c-Src/NADPH氧化酶/ROS通路誘導HO-1表達,從而抑制小鼠巨噬細胞過度分泌細胞因子。
芹菜素;血紅素氧合酶-1;小鼠巨噬細胞
膿毒血癥是指各種致病性微生物或其毒性產(chǎn)物持續(xù)存在于血液或組織中,并引起全身炎癥反應和器官功能損害為特征的一種臨床綜合征,全球每年發(fā)病人數(shù)可達1800萬以上。在美國,膿毒血癥在ICU中的發(fā)病率可達30%~50%。革蘭陰性細菌的脂多糖(Lipopolysaccharide,LPS)是引起膿毒血癥最常見的致病物質(zhì)[1]。早期革蘭陰性細菌感染后,可通過固有免疫系統(tǒng)識別LPS,隨后迅速激活單核、巨噬細胞系統(tǒng),并誘導多種促炎細胞因子如TNF-α,IL-1,IL-6以及一氧化氮、巨噬細胞趨化蛋白-1等物質(zhì)的產(chǎn)生,從而促進免疫系統(tǒng)清除病原體[2]。但如果致炎因素持續(xù)存在或炎癥失控,最終可導致嚴重的組織損傷以及多器官功能衰竭[3,4]。傳統(tǒng)治療急性炎癥反應主要采用固醇類和非甾體類藥物治療,但由于這些藥物副作用嚴重而使其應用受限[5],因此尋求新的抗炎輔助藥物對各類炎癥相關疾病的治療具有重要意義。
芹菜素(apigenin,APG)是廣泛分布于水果、蔬菜(尤其是芹菜、大蒜和大白菜)中的一種黃酮類化合物,具有抗腫瘤、抗血小板聚集、抗氧化等多種生物學作用[6]。此外,體內(nèi)外實驗也表明APG具有一定的抗炎效應。如,Wang等研究發(fā)現(xiàn),APG可通過ERK途徑抑制巨噬細胞分泌TNF-α和IL-1β[7]。此外,APG也可誘導小鼠原代肝細胞表達血紅素氧合酶-1(Heme oxygenase-1,HO-1),同時也能抑制尼古丁和LPS誘導牙周韌帶細胞分泌炎癥細胞因子[8]。也有研究表明,APG能抑制NF-κB的活性[6],但APG是否還存在其它的作用機制目前仍不明了。本研究旨在觀察APG對LPS誘導巨噬細胞分泌細胞的影響,并初步探索其分子機制。
1 材料與方法
1.1試劑DMEM培養(yǎng)基、胎牛血清購自Invitrogen;APG、β-actin多克隆抗體為Sigma-Aldrich產(chǎn)品。鼠抗人p-c-Src,p-p47phox以及鼠抗人total-Src和total-p47phox購自Cell Signaling。抗血紅素氧合酶-1(Heme oxygenase-1,HO-1)、以及各類二抗購自Santa Cruz。二亞苯基碘(Diphenyleneiodonium,DPI)、N-乙酰-半胱氨酸(N-acetyl-cysteine,NAC)以及2′,7′-二氯二氫熒光黃二乙酸酯(2′,7′-dichlorodihydrofluorescein diacetate,H2DCFDA)、鈷原卟啉(Cobalt protoporphyrin,CoPP)購自Sigma-Adrich。HO-1 siRNA購自Santa Cruz,TNF-α和IL-1β ELISA試劑盒購自R&D System。磷酸酶、蛋白酶抑制劑購自Roche,siRNA轉(zhuǎn)染試劑盒購自Qiagen。
1.2細胞培養(yǎng)與處理RAW 264.7細胞(ATCC)用含10%胎牛血清,100 U/mL青霉素和100 mg/mL鏈霉素,4.5 mg/mL L-谷氨酰胺以及4.5 g/mL的DMEM培養(yǎng)基中,于37 ℃,5% CO2條件下培養(yǎng)。2~3 d換液一次,待細胞生長至密度為80%時,根據(jù)不同的實驗目的,細胞加入不同濃度的APG作用0~16 h,或隨后加入1 μg/mL LPS刺激8 h。
1.3細胞總蛋白提取細胞處理結(jié)束后的細胞用冰冷PBS(pH7.4)漂洗1次,隨后加入100 μL裂解緩沖液A(10 mmol/L HEPES pH 7.9,1.5 mmol/L MgCl2,0.5 mmol/L DTT,5 μmol/L 亮抑蛋白酶肽,2 μmol/L胃酶抑素A,1 μmol/L 抑肽酶,20 μmol/L PMSF和1×磷酸酶抑制劑混合物),通過反復凍融以充分裂解細胞。隨后1 000×g離心10 min,獲取上清再次離心15 min(10 000×g)后即為胞漿總蛋白。
1.4 Western blot 采用含有蛋白酶抑制劑的RIPA緩沖液(25mmol/L Tris-HCl pH7.4,150mmol/L NaCl,1% NP-40,1% SDS)裂解細胞,并用Bradford法于595nm波長處測定蛋白濃度。取等量蛋白經(jīng)8%~10% SDS-PAGE后,轉(zhuǎn)印至PVDF膜上,用5%脫脂奶粉封閉后,分別加入一抗和HRP標記二抗孵育,ECL顯影。
1.5 ROS測定以H2DCFDA為熒光探針檢測細胞內(nèi)ROS水平。其原理是不發(fā)熒光的H2DCFDA能被過氧化物、氫過氧化物等氧化分解為二氯熒光黃而產(chǎn)生熒光,通過測定熒光的強度來反應ROS的含量。即,細胞中加入終濃度5 μmol/L的H2DCFDA染液,37 ℃避光孵育30 min。PBS洗細胞3次,重懸細胞,熒光分光光度計(SynergyHT,Bio-TeK)測量熒光強度(激發(fā)波長485 nm,發(fā)射波長530 nm),計算熒光的相對值(Relative fluorescence intensity)。
1.6 TNF-α和IL-6測定當生長于6孔板中的細胞長滿80~90%視野時,棄培養(yǎng)基,并加入不同濃度的APG作用1 h后加入LPS作用8 h。操作方法按照試劑盒提供的雙抗體夾心法進行。通過測定450 nm處的吸光度值,并根據(jù)標準曲線計算分泌至細胞外的TNF-α和IL-6含量。
1.7細胞轉(zhuǎn)染約5×105細胞接種于60 mm培養(yǎng)皿中18~24 h。隨后按照Qiagen提供的操作步驟,將100 nmol/L HO-1 siRNA或?qū)φ誷iRNA混合物孵育細胞。孵育4h后,用PBS漂洗細胞,換成完全培養(yǎng)基培養(yǎng)用于下一步研究。

2 結(jié) 果
2.1 APG誘導RAW264.7細胞c-Src磷酸化采用20 μmol/L APG作用RAW264.7細胞30 min即可誘導RAW264.7細胞中c-Src磷酸化,60 min后達到峰值,隨后逐漸降低并持續(xù)至4 h,而細胞內(nèi)總c-Src含量保持恒定(圖1)。
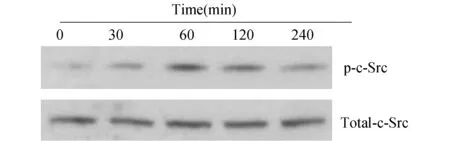
圖1 APG誘導RAW 264.7細胞c-Src磷酸化
2.2 APG誘導RAW264.7細胞中NADPH氧化酶p47phox亞基磷酸化Western blot結(jié)果顯示,空白對照組細胞NADPH氧化酶亞基p47phox磷酸化水平極低,20 μmol/L APG孵育30 min后,磷酸化p47phox有所增加,1 h后達到峰值。而同時采用50 μmol/L c-Src抑制劑PP1處理細胞1 h后,p47phox磷酸化水平明顯降低(圖2)。

圖2 APG經(jīng)c-Src誘導p47phox亞基磷酸化
2.3 APG激活NADPH氧化酶誘導ROS產(chǎn)生20 μmol/L APG處理RAW264.7細胞4 h后,可有效增加細胞內(nèi)ROS的含量。而細胞在APG處理前給予10 μmol/L DPI預處理1 h,結(jié)果顯示ROS含量明顯降低(圖3)。
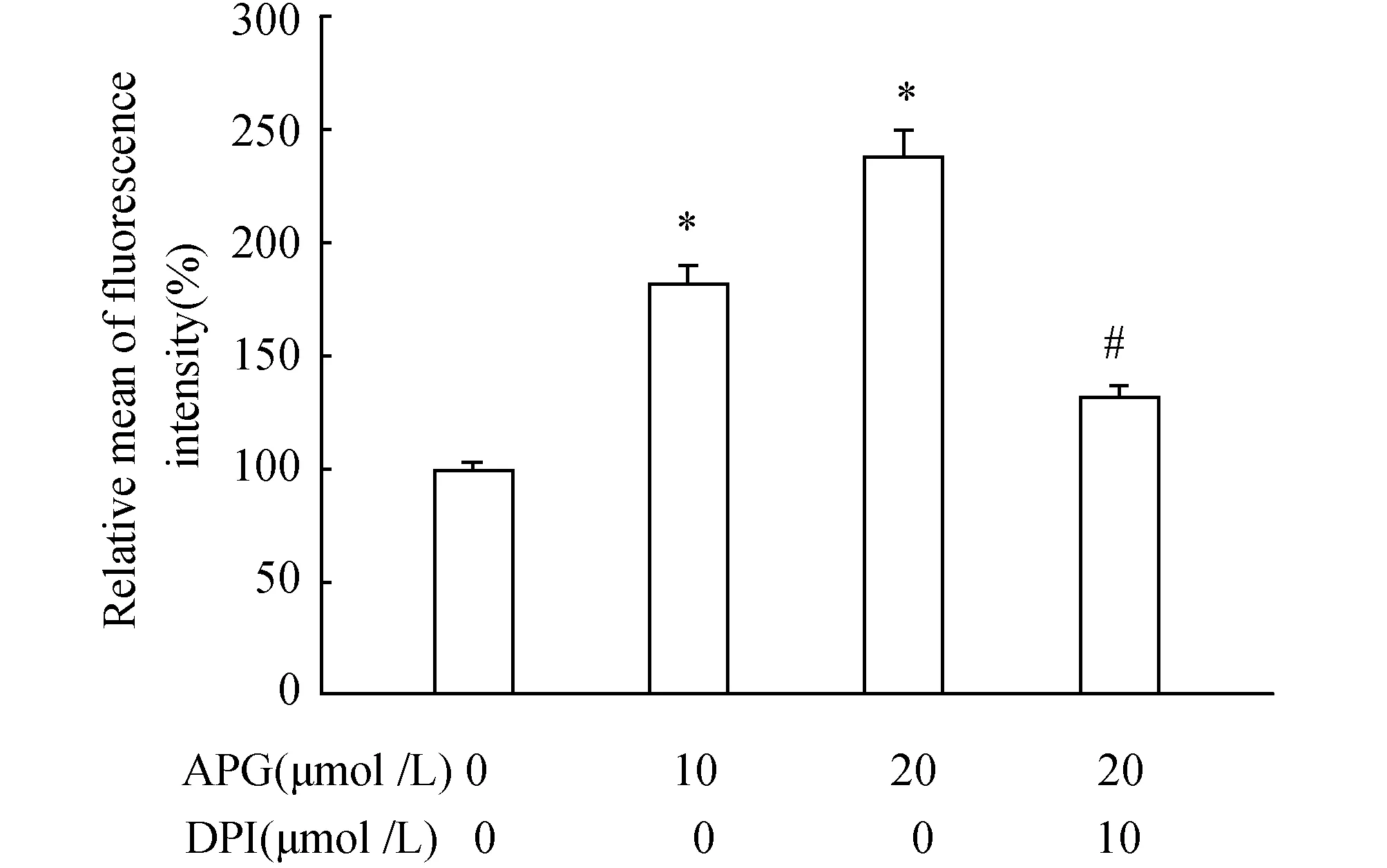
圖3 APG與DPI對ROS產(chǎn)生的影響 與對照組(0 μmol/L APG)相比,*P<0.05;與20 μmol/L APG相比,#P<0.05
2.4 APG對HO-1表達的影響0~20 μmol/L APG作用RAW264.7細胞16 h后,可顯著誘導其表達HO-1,并且呈一定的劑量依賴性(圖4A)。隨后用20 μmol/L APG作用細胞4 h后即可誘導RAW264.7細胞表達HO-1,隨APG作用時間的延長,HO-1表達水平逐漸遞增,持續(xù)到12 h~16 h(圖4B)。

圖4 APG誘導RAW 264.7細胞表達HO-1 A.不同濃度APG誘導RAW264.7細胞表達HO-1;B.20 μmol/L APG在不同時間內(nèi)誘導RAW264.7細胞表達HO-1
2.5 APG經(jīng)c-Src/NADPH氧化酶/ROS誘導HO-1表達RAW264.7細胞在APG處理前,分別用50 μmol/L c-Src抑制劑PP1、10 μmol/LNADPH氧化酶抑制劑DPI和10 mmol/L ROS抑制劑NAC預處理1 h,隨后再用20 μmol/L APG孵育16 h。結(jié)果顯示,PP1、DPI和NAC處理后,HO-1蛋白表達水平顯著降低(圖5)。

圖5 PP1、DPI、NAC對APG誘導HO-1表達的影響
2.6 APG上調(diào)HO-1表達從而負調(diào)控TNF-α和IL-6過度分泌RAW264.7細胞經(jīng)APG處理后,可顯著下調(diào)LPS誘導其分泌TNF-α和IL-6。而RAW264.7細胞采用siRNA干擾HO-1表達后,與未轉(zhuǎn)染組相比,TNF-α和IL-6含量明顯增高。此外,LPS刺激前采用10 μmol/L HO-1激動劑CoPP誘導4 h也得到了類似結(jié)果(圖6)。

圖6 HO-1對TNF-α和IL-6分泌的影響 與LPS相比,*P<0.05
3 討 論
巨噬細胞在各種病原微生物引起的炎癥反應中發(fā)揮重要作用。各種細菌和病毒均可活化巨噬細胞并誘導其分泌一系列炎癥細胞因子和介質(zhì)。盡管在急性期這些物質(zhì)對清除病原體感染、維持機體的穩(wěn)態(tài)方面具有積極作用,但巨噬細胞的持續(xù)活化也是引起各種病理性損傷的重要原因[3]。因此采用藥物干預炎癥反應發(fā)生的強度,是有效預防各類病理損傷的重要途徑。HO-1是催化血紅素降解為膽紅素、Fe2+和CO的限速酶,又稱為熱休克蛋白32。HO-1為誘導型,多種病理生理狀態(tài)可誘導其表達等[9]。Stuart等研究表明,LPS通過上調(diào)HO-1的表達,從而抑制單核細胞系THP-1過度分泌細胞因子[10]。此外,在動物模型中,誘導HO-1表達可調(diào)節(jié)流感病毒引起的免疫反應[11]。而HO-1基因敲出動物對氧化應激以及多器官功能衰竭的敏感性增加[12],因此HO-1的表達可能是宿主細胞免受氧化應激與炎癥損傷的一種獲得性保護機制。在本研究當中,我們證實APG能上調(diào)HO-1的表達。但其調(diào)控機制尚不明了。一般認為,HO-1的表達主要受核轉(zhuǎn)錄因子Nrf2的調(diào)控,此外,PKC,PI3K/Akt和NAPDH氧化酶也參與了HO-1的表達[13]。為了進一步探討APG的調(diào)控機制,本研究對HO-1可能的調(diào)控分子進行了觀察。c-Src是一種非受體型酪氨酸蛋白激酶,c-Src未激活時,其527位酪氨酸殘基(Tyr527)處于磷酸化狀態(tài)而使c-Src呈空間卷曲。各種因素導致c-Src激活后,Tyr527發(fā)生去磷酸化,同時誘導其Tyr416殘基磷酸化而使c-Src處于開放構(gòu)象而激活[14]。有研究顯示,多種病原體致病物質(zhì)可通過激活c-Src而誘導HO-1表達[15]。本研究也證實,巨噬細胞經(jīng)APG處理30min后即可檢測出磷酸化c-Src。c-Src激活后能發(fā)揮廣泛的酪氨酸激酶作用,如激活NAPDH氧化酶/ROS信號通路而誘導多種抗氧化、抗炎癥相關蛋白的表達[15]。NADPH氧化酶是一個多組分活化的酶復合體,主要由質(zhì)膜結(jié)合成分(包括gp91phox,p22phox,小GTP酶結(jié)合蛋白Rap1A)及胞漿成分組成(p47phox,p67phox,p40phox,小GTP酶結(jié)合蛋白 Rac2,Cdc42等)組成。外源性因素刺激下可誘導NADPH 氧化酶胞漿亞基p47phox磷酸化,隨后轉(zhuǎn)位到質(zhì)膜或顆粒體膜,從而組裝成有功能的NADPH氧化酶。因此p47phox的磷酸化是NAPDH氧化酶激活的標志[16]。在人體單核、巨噬細胞或中性粒細胞中,NADPH 氧化酶可通過一系列電子傳遞催化ROS的生成(氧化爆發(fā)),這是氧化應激或炎癥條件下ROS的主要來源。ROS是生物體內(nèi)產(chǎn)生的活性含氧化合物的總稱,包括超氧陰離子(O2·)、過氧化氫(H2O2)、羥自由基(HO·)、一氧化氮(NO·)等。過量的ROS可引起細胞大分子的氧化損傷,適量的ROS是調(diào)節(jié)細胞生理功能狀態(tài)的重要信號分子。本研究結(jié)果顯示,APG處理后可誘導p47phox磷酸化,并能誘導細胞內(nèi)ROS產(chǎn)生。且采用c-Src抑制劑PP1處理后,p47phox磷酸化水平明顯降低。此外,采用NADPH氧化酶抑制劑DPI處理后,ROS含量也明顯減少,表明APG處理后可激活c-Src/NADPH氧化酶/ROS通路。最后,分別采用c-Src、NAPDH氧化酶、ROS抑制劑PP1、DPI和NAC處理細胞后,HO-1的表達顯著降低。這表明APG誘導HO-1的表達受c-Src/NADPH氧化酶/ROS調(diào)控。
HO-1可通過多種機制發(fā)揮對細胞的保護作用及免疫調(diào)控作用。為了進一步探討為了明確HO-1在負調(diào)控炎癥反應中的作用,本研究通過靶向沉默HO-1表達以及藥物誘導的方式對TNF-α和IL-6的分泌進行了觀察。RNA干擾HO-1表達后,與單純LPS處理相比,TNF-α和IL-6分泌顯著增高。同時采用CoPP誘導HO-1表達后,TNF-α和IL-1β水平再次降低。以上結(jié)果表明HO-1表達能在一定程度上調(diào)控炎癥反應的強度,從而發(fā)揮對免疫功能的調(diào)控。
總之,本研究證實APG對LPS誘導巨噬細胞分泌細胞因子具有抑制作用。APG通過激活c-Src/NAPDH氧化酶/ROS信號通路,從而誘導HO-1的表達。當然,除了NADPH氧化酶以及ROS外,HO-1的表達還受PI3K/Akt以及蛋白激酶C的調(diào)控,在隨后的研究當種我們將會對其開展更深一步探討。
[1] Park BS,Lee JO.Recognition of lipopolysaccharide pattern by TLR4 complexes[J].Exp Mol Med,2013,45:e66.
[2] Roger T,Delaloye J,Chanson AL,et al.Macrophage migration inhibitory factor deficiency is associated with impaired killing of gram-negative bacteria by macrophages and increased susceptibility to Klebsiella pneumoniae sepsis[J].J Infect Dis,2013,207(2):331-339.
[3] Hotamisligil GS.Inflammation and metabolic disorders[J].Nature,2006,444(7121):860-867.
[4] Rosenblat JD,Cha DS,Mansur RB,et al.Inflamed moods:a review of the interactions between inflammation and mood disorders[J].Prog Neuropsychopharmacol Biol Psychiatry,2014,53:23-34.
[5] Walker JS,Nguyen TV,Day RO.Clinical response to non-steroidal anti-inflammatory drugs in urate-crystal induced inflammation:a simultaneous study of intersubject and intrasubject variability[J].Br J Clin Pharmacol,1994,38(4):341-347.
[6] Lefort EC,Blay J.Apigenin and its impact on gastrointestinal cancers[J].Mol Nutr Food Res,2013,57(1):126-144.
[7] Wang J,Liao Y,Fan J,et al.Apigenin inhibits the expression of IL-6,IL-8,and ICAM-1 in DEHP-stimulated human umbilical vein endothelial cells and in vivo[J].Inflammation,2012,35(4):1466-1476.
[8] Huang CS,Lii CK,Lin AH,et al.Protection by chrysin,apigenin,and luteolin against oxidative stress is mediated by the Nrf2-dependent up-regulation of heme oxygenase 1 and glutamate cysteine ligase in rat primary hepatocytes[J].Arch Toxicol,2013,87(1):167-178.
[9] 游曉星,馬小華,劉良專,等.支原體脂肽經(jīng)TLR2,6/c-Src/PI3K通路誘導單核細胞表達血紅素氧合酶-1[J].中國免疫學雜志,2014,30(5):587-590.
[10] Rushworth SA,MacEwan DJ,O'Connell MA.Lipopolysaccharide-induced expression of NAD(P)H:quinone oxidoreductase 1 and heme oxygenase-1 protects against excessive inflammatory responses in human monocytes[J].J Immunol,2008,181(10):6730-6737.
[11] Cummins NW,Weaver EA,May SM,et al.Heme oxygenase-1 regulates the immune response to influenza virus infection and vaccination in aged mice[J].FASEB J,2012,26(7):2911-2918.
[12] Gozzelino R,Jeney V,Soares MP.Mechanisms of cell protection by heme oxygenase-1[J].Annu Rev Pharmacol Toxicol,2010,50:323-354.
[13] Zou W,Chen C,Zhong Y,et al.PI3K/Akt pathway mediates Nrf2/ARE activation in human L02 hepatocytes exposed to low-concentration HBCDs[J].Environ Sci Technol,2013,47(21):12434-12440.
[14] Byeon SE,Yi YS,Oh J,et al.The role of Src kinase in macrophage-mediated inflammatory responses[J].Mediators Inflamm,2012,2012:512926.
[15] Lee IT,Wang SW,Lee CW,et al.Lipoteichoic acid induces HO-1 expression via the TLR2/MyD88/c-Src/NADPH oxidase pathway and Nrf2 in human tracheal smooth muscle cells[J].J Immunol,2008,181(7):5098-5110.
[16] Pick E.Role of the Rho GTPase Rac in the activation of the phagocyte NADPH oxidase:outsourcing a key task[J].Small GTPases,2014,5:e27952.
ApigeninInducesHO-1Expressionbyc-Src/NADPHOxidase/ROStoProtectAgainstExcessiveCytokinesSecretioninMurineMacrophages
ZENG Saili,WU Guang,XIE Li,et al
(DepartmentofRespiratoryMedicine,TheSecondAffiliatedHospital,UniversityofSouthChina,Hengyang421001,Hunan,China)
ObjectiveTo investigate the effect and the mechanism of Apigenin on lipopolysaccharides (LPS)-induced cytokines production in murine macrophages.MethodsThe murine macrophage cell line RAW 264.7 cells were cultured in vitro,and were treated with different concentration of Apigenin followed by LPS stimulation.Expression of heme oxygenase-1 (HO-1),phosphorylation of c-Src and p47phox,the subunit of NADPH oxidase were detected by Western blot;The intracellular formation of reactive oxygen species (ROS) was detected by using the fluorescent probe H2DCFDA;Secretion of TNF-α and IL-6 were detected by ELISA.ResultsWestern blot indicated that phosphorylated c-Src and p47phoxwere induced by apigenin after 30min of incubation.While 50 μmol/L of PP1,an inhibitor of c-Src,could significantly inhibit p47phoxphosphorylation.In addition,Apigenin could also induce RAW264.7 cells accumulation of ROS,and this effect could be abrogated by the NAPDH oxidase inhibitor,DPI.HO-1 could be induced by Apigenin,and pretreatment of PP1,DPI and the ROS scavenger N-acetyl-cysteine (NAC) could block HO-1 production.Furthermore,apigenin could inhibit LPS-induced secretion of TNF-α and IL-6,and transfection of HO-1 siRNA could antagonize this action.ConclusionApigenin can induce HO-1 expression by c-Src/NADPH oxidase/ROS to protect against excessive cytokines secretion in murine macrophages.
Apigenin; Heme oxygenase-1; Murine macrophages
10.15972/j.cnki.43-1509/r.2015.04.003
2015-04-22;
2015-05-23
國家自然科學基金(31370207).
*通訊作者,E-mail:zengyihua21cn@126.com.
R967
A
(此文編輯:秦旭平)
