GABAA受體基因多態(tài)性與癲癇易感性的關系
2015-12-29 03:05:49張虎,杜欣娜,徐敏等
中國老年學雜志 2015年19期
關鍵詞:癲癇
GABAA受體基因多態(tài)性與癲癇易感性的關系
張虎杜欣娜1徐敏張淑紅王菊莉2張金波朱金玲
(佳木斯大學基礎醫(yī)學院,黑龍江佳木斯154007)
摘要〔〕目的探究γ-氨基丁酸A型(GABAA)受體的亞基基因GABRG2和GABRB2單核苷酸多態(tài)性(SNP)與癲癇(EP)易感性的關系。方法應用限制性片段長度多態(tài)性(RFLP)技術對GABAA受體亞基基因GABRB2(rs12653921、rs2964773)、GABRG2(rs2268581)3個標簽單核苷酸多態(tài)性(Tag SNP)的等位基因和基因型測定分析。結果EP組和對照組等位基因頻率和基因型分布比較,SNP(rs12653921)位點(基因型χ2=8.651,等位基因χ 2=8.649)有統(tǒng)計學意義;SNP(rs2268581)、SNP(rs2964773)位點(rs2268581:基因型χ2=0.354,等位基因χ2=0.294;rs2964773:基因型χ2=1.397,等位基因χ2=0.643)無顯著差異(P>0.05)。結論SNP(rs12653921)可能與EP易感性相關,SNP(rs2268581)、SNP(rs2964773)可能與EP易感性不相關。
關鍵詞〔〕癲癇;GABAA受體;單核苷酸多態(tài)性
中圖分類號〔〕R742.1〔文獻標識碼〕A〔
基金項目:黑龍江省衛(wèi)生廳課題(2012-248)
通訊作者:朱金玲(1967-),女,教授,主要從事癲癇易感性研究。
1佳木斯大學附屬第二醫(yī)院病理科2佳木斯市中心醫(yī)院癲癇科
第一作者:張虎(1981-),男,講師,主要從事癲癇易感性研究。
癲癇(EP)是神經系統(tǒng)內因神經元異常放電而引起短暫的腦功能紊亂的一種多發(fā)病,發(fā)病機制十分復雜,而神經元的超同步放電引發(fā)癇性放電的理論則被一致接受。γ-氨基丁酸(GABA)在中樞神經系統(tǒng)內含量最多的抑制性神經遞質,其抑制作用由GABA受體介導,維持了神經元的興奮與抑制平衡,該平衡被打破導致神經元失去細胞膜穩(wěn)定性而引起EP發(fā)作〔1〕。GABA受體主要包括3個亞型家族:GABAA、GABAB和GABAC受體,其中GABAA受體是目前研究最廣泛、與EP關系最密切的亞型。
1對象與方法
1.1研究對象EP組為2012年3月至2013年9月就診于佳木斯中心醫(yī)院癲癇中心的EP患者198例,其中男102例,女96例,均為漢族,年齡2個月~70歲,均經過臨床確診(依照1981年ILAE分類和診斷標準);對照組為同期于本院體檢中心健康查體的人員186例,其中男90例,女96例,均為漢族,年齡3~75歲。兩組一般資料比較無統(tǒng)計學意義(P>0.05)。
1.2研究方法
1.2.1Tag SNP的選擇在Hapmap中分別載入基因GABRB2和GABRG2基因后,在Haploview 4.2軟件中運行tagger program選取各自的Tag SNP,并且從中挑選具有限制性酶切位點的SNP位點。選取了GABRB2基因的rs12653921、rs2964773和GABRG2基因的rs2268581。
1.2.2基因組DNA的提取取外周靜脈血2 ml,用ACD液(檸檬酸鈉1.32 g,檸檬酸0.48 g,葡萄糖1.47 g,加水至100 ml)抗凝,以低滲溶血,裂解白細胞,用chelex-100提取DNA。
1.2.3PCR-RFLP檢測GABAA受體基因的多態(tài)性①用primer5.0軟件設計PCR引物。20 μl PCR反應體系:2×Power Taq PCR Master Mix 8 μl,上、下游引物各0.5 μl,模板DNA 2 μl,加ddH2O至20 μl。各Tag SNP位點PCR反應條件為:95℃預變性5 min,然后進行35個PCR循環(huán)(95℃變性30 s、相應溫度退火30 s、72℃延伸相應時間),72℃終末延伸5 min。rs12653921上游引物:ATAGACCAAAAGCTGGGCCTC,下游引物:TGGTCCATGGGCTGTAGAATG,退火溫度62℃,延伸時間60 s,片段長度907 bp;rs2268581上游引物:TGGGGTTTGTTTCAGCGTGT,下游引物:CCTAGAACCACATCCCTTGCT,退火溫度60℃,延伸時間30 s,片段長度498 bp;rs2964773上游引物:CCTTGCCCCTGGTTTCATCT,下游引物:TCTTGTGTAACTCAGCATTTCTCA,退火溫度59℃,延伸時間30 s,片段長度353 bp。②RFLP檢測:20 μl的酶切反應體系包括 PCR產物10 μl,限制性內切酶1.5 μl,10×buffer 2.0 μl,ddH20 8.0 μl。其中rs12653921、rs2268581、rs2964773位點的內切酶分別為BglⅡ酶、HincⅡ酶、Taq I酶。 酶切反應體系配好振蕩混勻后,置于37℃水浴箱中,酶切時間分別是16 h、4 h、5 min。
1.2.4PCR產物的測定三種基因型部分產物經純化后進行DNA全自動測序(上海生工),進行基因型驗證。
1.3統(tǒng)計學處理采用SPSS17.0軟件進行χ2檢驗。
2結果
2.1rs12653921結果Rs12653921PCR產物大小為907 bp,其中T/T:498 bp和409 bp;G/G型:907 bp;G/T型:498 bp、409 bp和907 bp。見圖1。兩組基因型和等位基因頻率比較見表1。
2.2rs2268581結果rs2268581 PCR產物大小為498 bp,A/A:388 bp和111 bp;T/T:498 bp;A/T:388 bp、111 bp 和498 bp,見圖2。兩組rs2268581基因型和等位基因比較見表2。
與對照組比較:1)P<0.05
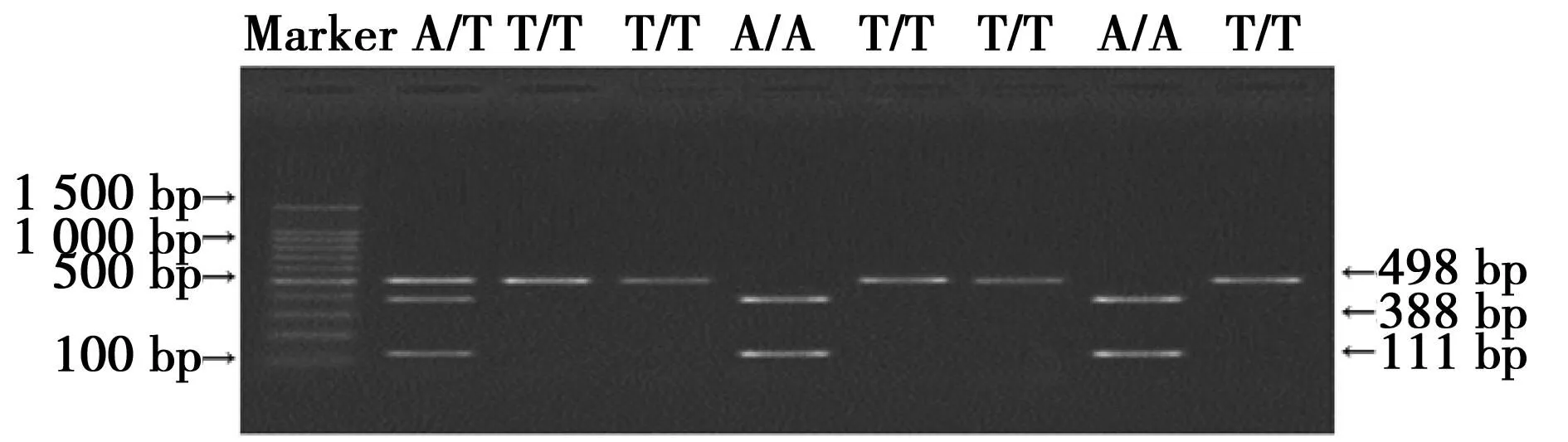
圖2 rs2268581位點酶切圖
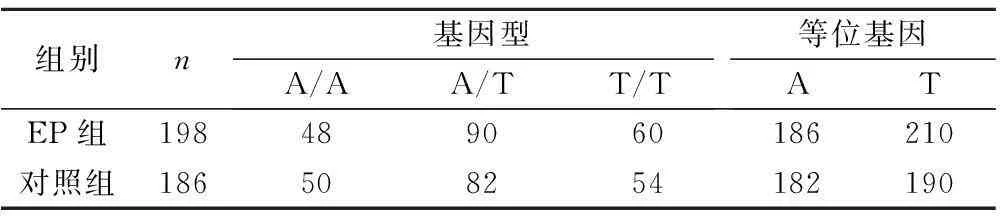
組別n基因型A/AA/TT/T等位基因ATEP組198489060186210對照組186508254182190
2.3rs2964773結果PCR產物大小為353 bp,其中該位點基因型及等位基因頻率比較見表3。C/C:241 bp和112 bp;T/T:353 bp;C/T:241 bp、112 bp和353 bp,見圖3。
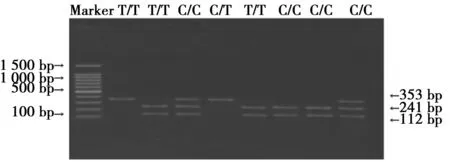
圖3 rs2964773位點酶切圖
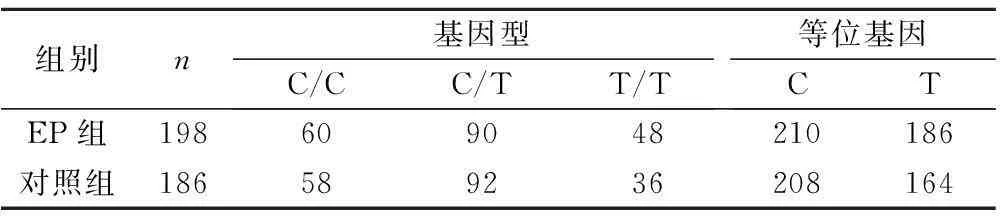
組別n基因型C/CC/TT/T等位基因CTEP組198609048210186對照組186589236208164
3討論
在結構上,GABAA受體是異源五聚體配體門控的氯離子通道,包括α(α1-α6),β(β1-β3),γ(γ1-γ3);δ,ε,θ和ρ(ρ1-ρ3)亞基基因家族編碼的各種蛋白〔2〕,不同亞基在EP發(fā)病中起到不同的作用。最常見的組合是2ɑ1 2β22γ2,是腦組織內含量最豐富的組合。GABA和其重要正性變構劑BD的結合位點在ɑ、β亞基上,介導GABAA受體對神經元的抑制性作用,而γ亞基對其抑制作用有重要輔助作用。因此本實驗選取了GABRG2及GABRB2基因的3個Tag SNP位點進行基因多態(tài)性研究。隨著遺傳技術的快速發(fā)展,GABAA受體亞基基因水平的研究逐漸深入。GABRG2基因第8外顯子K289M突變,是發(fā)現(xiàn)的第一個與EP相關的GABAA受體基因變異〔3〕。Q351X、第6內含子剪接位點(IVS6+2T→G)突變、第2外顯子的R43Q錯義變異,主要分布在CAE和GEFS+家系中〔4,5〕。另外,R177G、p.W390X突變,主要與FS和FS+相關,錯義突變 (c.236A>G:p.N40S)在GTCS患者中被確定〔6~8〕。GABRB2基因在GEFS+家系中發(fā)現(xiàn)第2外顯子的C/G多態(tài)性〔9〕。第11外顯子SNP(1412C>T)導致丙氨酸殘基同義突變〔10〕在對北印度人研究中與癲癇易感性有相關性。
SNP是在人類基因組中分布密集的一種分子標記,大約每1 000 bp有一個SNP位點。由于基因連鎖不平衡,遺傳距離而非物理距離相近的SNP位點的等位基因不是隨意組合而是趨于一種組合,而是形成相對固定的單倍型域。單倍型域可大可小,其中的SNP位點數(shù)目不定。由于單倍型域內的各個SNP是相互關聯(lián)的,它們可以用一個位點(或多個位點的組合)代表另外的位點或單倍型,所選的有代表性的位點即Tag SNP。人類基因組單體型圖計劃中單倍型的圖譜的制作就是Tag SNP的應用的成果。Tag SNP在疾病關聯(lián)分析中被廣泛應用。
rs12653921定位于內含子區(qū),作用機制可能是因為堿基的改變影響了mRNA穩(wěn)定性,不能編碼功能性GABRB2基因蛋白質,β2亞基的表達受到了明顯的影響甚至表達缺失,最終誘發(fā)EP的發(fā)生。證實了β2亞基基因變異在EP發(fā)病機制中的重要作用,堿基的改變最終都是影響了GABAA受體的組裝及其表面表達,增加了癲癇的易感性。SNP(rs12653921)是Tag SNP,也有可能是因其所在的單倍域內其他多態(tài)位點與EP易感性相關,而SNP(rs12653921)只是因與其連鎖不平衡所致基因型分析有差異,因而不起決定作用。
在GABRG2基因SNP(rs2268581)、SNP(rs2964773)的研究中,未發(fā)現(xiàn)與EP易感性有相關性。可能是以下原因所致:(1)EP的發(fā)病機制絕大多數(shù)是多基因遺傳,其發(fā)生是幾個基因共同作用的結果,所以在樣本量難以滿足理想樣本時,很可能難以發(fā)現(xiàn)某一基因的微小作用。(2)在與EP發(fā)作相關的基因中的數(shù)個SNP位點中,可能僅是其中一部分是在癲癇發(fā)作機制中起作用,而另一部分可能僅與其他神經系統(tǒng)疾病相關。(3)種族、地域差異,在以往GABAA受體基因與EP的相關性研究中,同樣存在種族不同而相關性結果分析不一致的情況。EP是一組遺傳異質性的,以后隨著基因工程的技術進展,與EP相關的多態(tài)位點將為新藥物提供有利依據(jù)。
綜上所述,GABAA受體基因變異使介導神經元抑制作用的GABAA受體的亞基的合成、組裝及表面表達受到影響,受體功能最終減弱甚至缺失以致癲癇發(fā)生。越來越多GABAA受體亞基的突變被確定,而且隨著基因克隆技術及動物實驗技術的進步,很多突變的機制在進一步的試驗中得以印證。研究的最終目標是能夠針對突變位點研制新的靶點藥物,而現(xiàn)階段的水平還遠遠不足。需要進一步擴大樣本量,來探索GABAA受體基因在EP的發(fā)病機制中的作用。
4參考文獻
1Treiman DM.GABAergic mechanisms in epilepsy〔J〕.Epilepsia,2001;42(Suppl 3):8-12.
2Fritschy JM.Epilepsy E/I balance and GABA(A) receptor plasticity〔J〕.Front Mol Neurosci,2008;28(1):5.
3李袁華,周春雷,趙德明,等.癲癇持續(xù)狀態(tài)致 MODS 的發(fā)生率及其相關因素分析〔J〕.中華全科醫(yī)學,2008;6(9):925-6.
4Wallace RH,Marini C,Petrou S,etal.Mutant GABAA receptor gamma 2-subunit in childhood absence epilepsy and febrile seizures〔J〕.Nat Genet,2001;28(1):49-52.
5Harkin LA,Bowser DN,Dibbens LM,etal.Truncation of the GABAA receptor gamma2 subunit in a family with generalized epilepsy with febrile seizures plus〔J〕.Am J Hum Genet,2002;70(2):530-6.
6Audenaert D,Schwartz E ,Claeys KG,etal. A novel GABRG2 mutation associated with febrile seizures〔J〕. Neurology,2006;67(4):687-90.
7Sun H, Zhang Y, Liang JM,etal. SCN1A, SCN1B, and GABRG2 gene mutation analysis in Chinese families with generalized epilepsy with febrile seizures plus〔J〕. J Hum Genet ,2008;53(8):769-74.
8Shi X, Huang MC, Ishii A,etal. Mutational analysis of GABRG2 in a Japanese cohort with childhood epilepsies〔J〕. J Hum Genet,2010;55(6):375-8.
9張守山, 黃希順,王家勤,等.全面性癲癇伴熱性驚厥附加癥家系GABRB2基因測序研究〔J〕.實用兒科臨床雜志,2006;21(1):30-1.
10Kumari R,Lakhan R, Kalita J,etal. Potential role of GABAA receptor subunit: GABRA6,GABRB2 and GABRR2 gene polymorphisms in epilepsy susceptibility and pharmacotherapy in North Indian population〔J〕.Clin Chimica Acta, 2011;3(18):1244-8.
〔2014-03-11修回〕
(編輯李相軍)
猜你喜歡
中國民間療法(2021年5期)2021-06-09 09:21:04
中華養(yǎng)生保健(2020年2期)2020-11-16 00:49:00
解放軍醫(yī)學院學報(2020年12期)2020-03-29 05:11:46
中成藥(2017年6期)2017-06-13 07:30:35
飲食科學(2017年5期)2017-05-20 17:11:53
臨床醫(yī)藥文獻雜志(電子版)(2017年11期)2017-05-17 04:48:10
安徽醫(yī)科大學學報(2015年9期)2015-12-16 11:09:44
中國當代醫(yī)藥(2015年7期)2015-03-01 02:01:13
西南軍醫(yī)(2015年4期)2015-01-23 01:19:30
西部中醫(yī)藥(2014年6期)2014-03-11 16:07:47
