日本囊對(duì)蝦(Marsupenaeus japonicus)秋繁仔蝦形態(tài)表型與抗流性能間的相關(guān)性*
2016-01-15 06:46:39王志錚朱海軍任夙藝斯烈鋼祝本強(qiáng)
海洋與湖沼 2016年2期
關(guān)鍵詞:差異實(shí)驗(yàn)
王志錚 朱海軍 任夙藝 斯烈鋼, 祝本強(qiáng)
(1. 浙江海洋大學(xué) 舟山 316022; 2. 寧波市海洋與漁業(yè)研究院 寧波 315000)
水流作為水域生態(tài)系物質(zhì)和能量輸運(yùn)的重要載體, 直接或間接地決定著對(duì)蝦的區(qū)域分布, 是影響對(duì)蝦存活與生長(zhǎng)的重要環(huán)境因子(Dall et al, 1992; 戴習(xí)林等, 2008)。日本囊對(duì)蝦屬沿岸定居性蝦種(王克行,1997), 系近年來我國東南沿海重要的增殖放流對(duì)象之一。雖Dall 等(1992)認(rèn)為長(zhǎng)期生活于沿岸水域的對(duì)蝦為適應(yīng)潮汐運(yùn)動(dòng)形成了對(duì)水平流的適應(yīng)性, 但自主運(yùn)動(dòng)能力較弱的仔蝦, 仍極易受潮流作用的影響而被沖離增殖定居區(qū)。因此, 日本囊對(duì)蝦仔蝦的增殖放流效果勢(shì)必與其抗流能力有著極為密切的關(guān)系,開展該蝦仔蝦抗流性能的等級(jí)篩選與定向選擇研究無疑對(duì)于促進(jìn)其增殖漁業(yè)的健康發(fā)展具重要現(xiàn)實(shí)意義。
水生動(dòng)物大多具有逆水頂流的生活習(xí)性。已有研究表明, 水生動(dòng)物在靜水狀況下游泳時(shí)的耗氧率明顯低于流水狀況, 且隨流速的提高, 游泳速度的加快, 耗氧率也相應(yīng)增加(Wardle et al, 1996;Herskin et al, 1998; Lee et al, 2003); 形態(tài)表型不僅會(huì)改變水生動(dòng)物的游泳能力, 而且還受到不同流速生境的影響(Langerhans et al, 2003; Haas et al,2010)。王吉橋等(1999)指出, 蝦體不同部位異速生長(zhǎng)的階段性和時(shí)序性與其器官結(jié)構(gòu)和功能完善及強(qiáng)化的順序有關(guān), 是機(jī)體內(nèi)新陳代謝相輔相成反應(yīng)過程中起主導(dǎo)地位的方面發(fā)生變化所致。無疑, 對(duì)蝦的頂流運(yùn)動(dòng)能力與形態(tài)表型及其所映射的能耗效率間均有著極為密切的關(guān)系, 這就為從表型水平劃分仔蝦的抗流性能等級(jí)提供了啟示。但迄今國內(nèi)外尚未見有關(guān)日本囊對(duì)蝦形態(tài)表型對(duì)其抗流性能影響的研究報(bào)道。鑒于此, 本文作者于 2014年 10月以日本囊對(duì)蝦秋繁同生群仔蝦為研究對(duì)象, 借助多元分析方法, 較系統(tǒng)研究了不同抗流性能群體間的形態(tài)表型差異, 旨為該蝦秋繁同生群仔蝦抗流性能評(píng)價(jià)方法的構(gòu)建和逆境生態(tài)學(xué)研究提供基礎(chǔ)資料。
1 材料與方法
1.1 日本囊對(duì)蝦仔蝦
由寧波市海洋與漁業(yè)研究院提供, 總長(zhǎng)(7.643±0.639)mm。運(yùn)回浙江海洋學(xué)院水產(chǎn)安全養(yǎng)殖實(shí)驗(yàn)室后,即刻分置于若干規(guī)格為 60cm×80cm×60cm 的小型實(shí)驗(yàn)水槽內(nèi), 靜水停飼暫養(yǎng)24h后備用。
1.2 理化條件
實(shí)驗(yàn)用水為經(jīng)暗沉淀處理24h的過濾海水, 水溫(18.0±1.0)°C、鹽度 20、pH 8.1±0.2, 水質(zhì)符合 NY 5052-2001無公害食品 海水養(yǎng)殖用水水質(zhì)(中華人民共和國農(nóng)業(yè)部, 2001)要求。水流測(cè)定裝置見圖1。
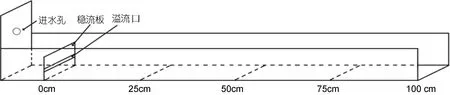
圖1 水流測(cè)定裝置示意圖(水流方向自左至右)Fig.1 The flume test
1.3 實(shí)驗(yàn)方法
自制總流長(zhǎng)為1m的水流測(cè)定裝置, 以溢流口底部為起點(diǎn), 按等距離間距法將該裝置依次劃分為A[0—25cm]、B(25—50cm)、C(50—75cm)、D(75—100cm)和 E(>100cm)等五個(gè)實(shí)驗(yàn)區(qū)段(圖 1)。任選 3000尾仔蝦, 于實(shí)驗(yàn)時(shí)將它們均勻分置于 30個(gè)上述水流測(cè)定裝置的溢水口底部, 以 1min為流速處理時(shí)限,以經(jīng)預(yù)實(shí)驗(yàn)確認(rèn)在實(shí)驗(yàn)終了時(shí)刻仍分布于 A區(qū)段內(nèi)的仔蝦占實(shí)驗(yàn)仔蝦總數(shù) 5%的流速 0.823cm/s為實(shí)驗(yàn)流速, 于實(shí)驗(yàn)終了時(shí)刻隨機(jī)選取分布于A、B、C、D、E區(qū)段內(nèi)的仔蝦群體(依次定義為A、B、C、D、E實(shí)驗(yàn)群體) 各 100尾作為測(cè)量樣本, 借助研究體視鏡(Lecia S8APO型)并采用掃描像素法(楊勁峰等, 2002),逐尾測(cè)量其總長(zhǎng)(X1, 額角前緣至尾節(jié)末端的直線長(zhǎng)度)、第一觸角柄長(zhǎng)(X2)、額劍長(zhǎng)(X3)、眼徑(X4)、頭胸甲長(zhǎng)(X5)、頭胸甲高(X6)、第一腹節(jié)長(zhǎng)(X7)、第二腹節(jié)長(zhǎng)(X8)、第三腹節(jié)長(zhǎng)(X9)、第四腹節(jié)長(zhǎng)(X10)、第五腹節(jié)長(zhǎng)(X11)、尾節(jié)長(zhǎng)(X12)、尾扇長(zhǎng)(X13)、腹節(jié)高(X14)和尾節(jié)高(X15)等15項(xiàng)形態(tài)學(xué)指標(biāo)。
1.4 數(shù)據(jù)處理
根據(jù)所測(cè)結(jié)果, 分別計(jì)算A、B、C、D、E實(shí)驗(yàn)群體的各項(xiàng)形態(tài)性狀與形態(tài)比例性狀的均值和標(biāo)準(zhǔn)差, 并采用 LSD多重比較法檢驗(yàn)組間差異顯著性(P<0.05視為顯著水平); 根據(jù)上述5個(gè)不同抗流性能實(shí)驗(yàn)群體各形態(tài)比例性狀的均值, 計(jì)算它們間經(jīng)標(biāo)準(zhǔn)化處理的歐氏距離(全距為 1.00), 并以此度量它們?cè)谛螒B(tài)體型比例上的相似性程度(P<0.05視為顯著水平); 對(duì)所有測(cè)量樣本的形態(tài)比例性狀進(jìn)行主成分分析, 以特征值大于1且累計(jì)貢獻(xiàn)率大于80%為原則確定主成分(PC)的提取個(gè)數(shù); 將A實(shí)驗(yàn)群體定義為水流脅迫處理選留群, B、C、D、E實(shí)驗(yàn)群體統(tǒng)歸為水流脅迫處理淘汰群。采用逐步導(dǎo)入剔除法對(duì)上述兩類群進(jìn)行判別分析, 并計(jì)算判別準(zhǔn)確率(P1為某類群實(shí)驗(yàn)對(duì)象判別正確的個(gè)體數(shù)占該類群實(shí)驗(yàn)對(duì)象實(shí)際總數(shù)的百分比; P2為諸實(shí)驗(yàn)類群在等樣本容量條件下, 某類群實(shí)驗(yàn)對(duì)象判別正確的個(gè)體數(shù)占判入該類群實(shí)驗(yàn)對(duì)象的總個(gè)體數(shù)的百分比)和綜合判別率(P為諸實(shí)驗(yàn)類群在等樣本容量條件下, 各類群實(shí)驗(yàn)對(duì)象判別正確的個(gè)體數(shù)之和占實(shí)驗(yàn)總個(gè)體數(shù)的百分比)以驗(yàn)證所建判別方程組的可靠性。上述計(jì)算分析均借助 SPSS 17.0軟件來完成。
2 結(jié)果
2.1 抗流特征
觀察發(fā)現(xiàn), 實(shí)驗(yàn)期間絕大多數(shù)仔蝦一直處于頂流運(yùn)動(dòng)狀態(tài)。實(shí)驗(yàn)結(jié)束時(shí)刻, 分布于A、B、C、D、E區(qū)段內(nèi)的實(shí)驗(yàn)仔蝦占比均值分別為 5%、11.33%、18.67%、25.33%和39.67% (圖2), 呈單調(diào)增加趨勢(shì)。根據(jù)統(tǒng)計(jì)學(xué)中的小概率事件實(shí)際不可能性原理(蓋均鎰, 2000), 可將流速0.823cm/s確定為實(shí)驗(yàn)處理時(shí)長(zhǎng)1min條件下實(shí)驗(yàn)仔蝦[總長(zhǎng)(7.643±0.639)mm]的抗流極限值。
2.2 各實(shí)驗(yàn)群體形態(tài)性狀測(cè)量值和形態(tài)比例指標(biāo)值間的差異比較
由表1可見, 本研究所涉5個(gè)不同抗流性能實(shí)驗(yàn)群體在所測(cè)15項(xiàng)形態(tài)性狀中, 除X1、X2、X3、X11、X15均呈 A>E>D>C>B (P<0.05),X12呈 A>E≈D>C≈B,X13呈 A>E≈D>C>B外, 其余 8項(xiàng)形態(tài)性狀測(cè)量指標(biāo)值均無組間差異(P>0.05); 由表2可見, 在所涉17項(xiàng)形態(tài)比例指標(biāo)中,C1僅C、D間具顯著差異,C3、C4、C6、C7、C8均呈 B>C>D>E>A (P<0.05),C12、C13均呈A>E>D>C>B (P<0.05),C14僅 C 分別與 D、E 具顯著差異, 其余 7項(xiàng)形態(tài)比例指標(biāo)值均無組間差異(P>0.05)。由此可見, 實(shí)驗(yàn)群體間抗流能力的分化源自它們間形態(tài)表型的差異, 在表征實(shí)驗(yàn)仔蝦抗流能力的差異上形態(tài)比例性狀較形態(tài)性狀本身更具豐盛度和有序性。
2.3 各實(shí)驗(yàn)群體形態(tài)比例特征間的相似度比較
根據(jù)本研究所涉 5個(gè)不同抗流性能實(shí)驗(yàn)群體各項(xiàng)形態(tài)比例性狀的均值(表 2), 分別計(jì)算它們間的歐氏距離(表 3)。由表 3可見, 各實(shí)驗(yàn)群體間的歐氏距離均達(dá)到顯著水平(P<0.01), 且均有隨實(shí)驗(yàn)群體間抗流性能差異的增大而呈顯著增大的趨勢(shì), 進(jìn)一步表明可用形態(tài)比例性狀來表征實(shí)驗(yàn)群體間抗流性能的差異。
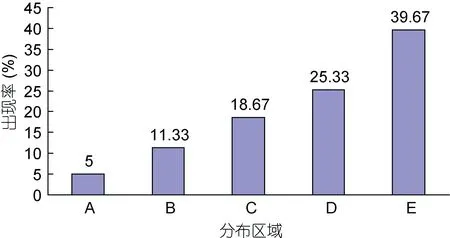
圖2 實(shí)驗(yàn)終了時(shí)刻仔蝦在A、B、C、D、E區(qū)段的出現(xiàn)率Fig.2 The frequency of appearance at the end of experiment at sections A, B, C, D, and E
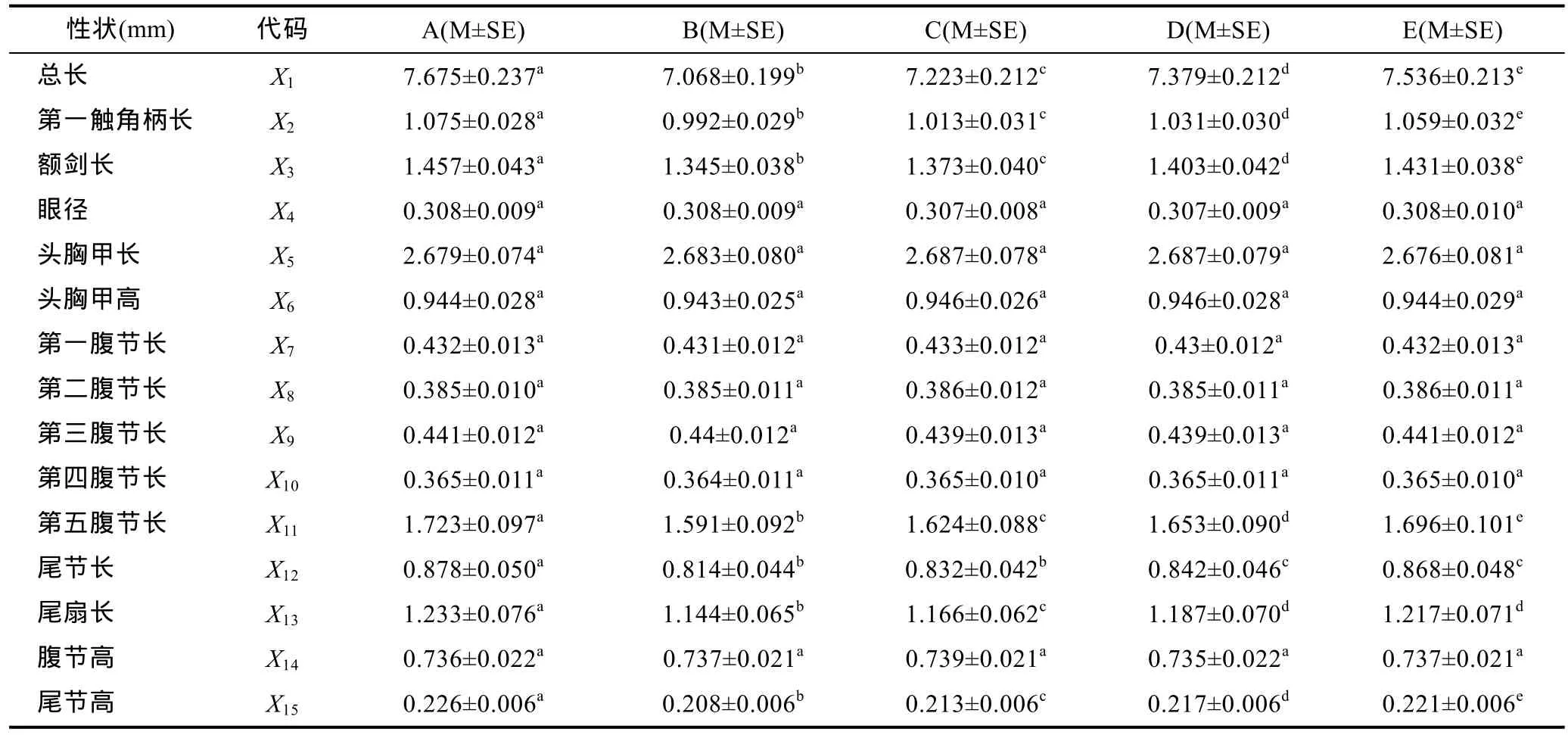
表1 實(shí)驗(yàn)群體形態(tài)性狀測(cè)量值的參數(shù)統(tǒng)計(jì)(n=100)Tab.1 Statistics of morphological measurements in all experimental groups
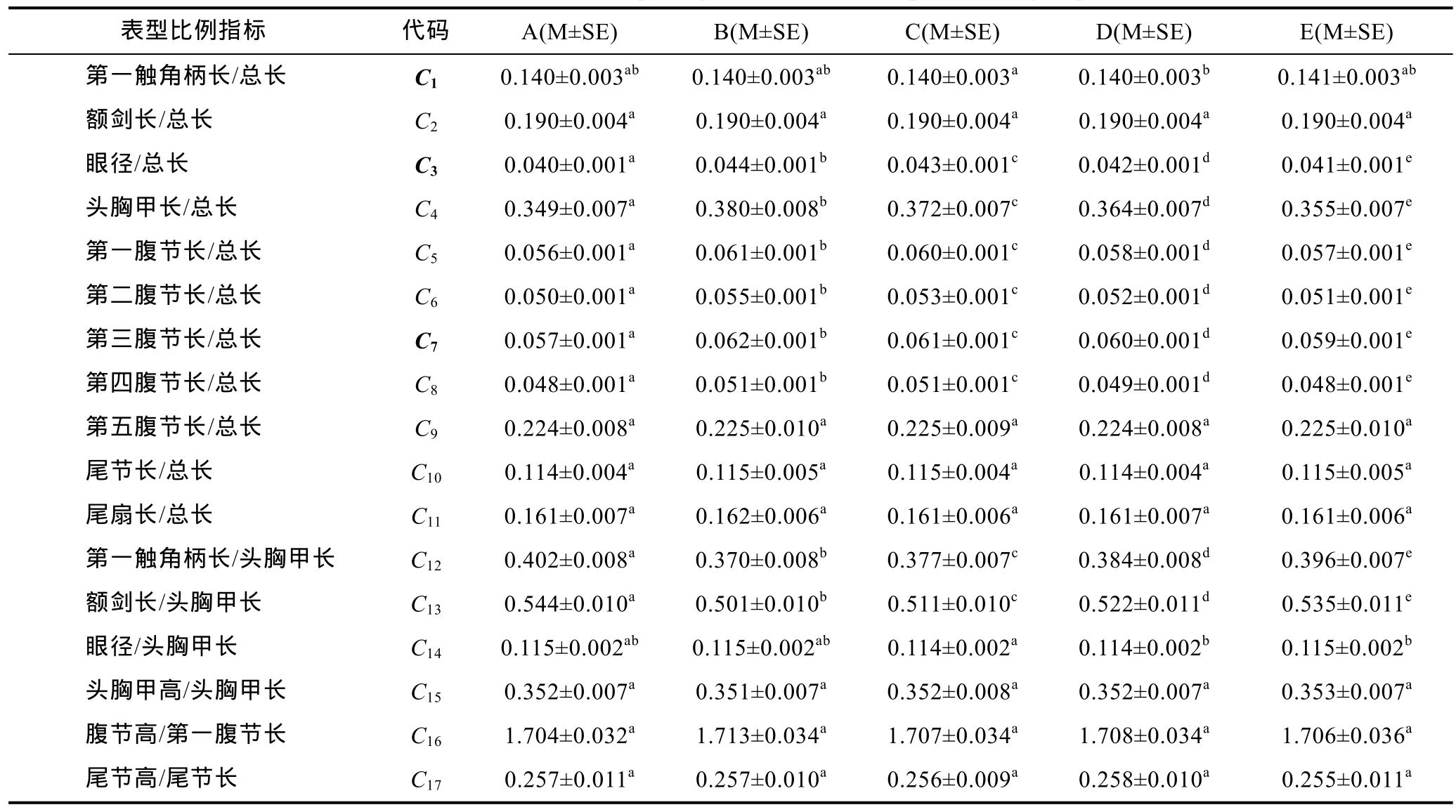
表2 實(shí)驗(yàn)群體形態(tài)比例特征間的差異(n=100)Tab.2 Differences in morphological traits for all experimental groups
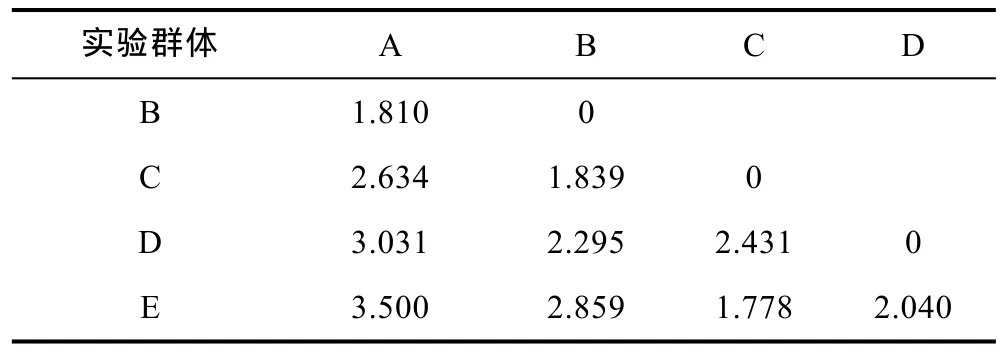
表3 實(shí)驗(yàn)群體形態(tài)比例特征間的歐氏距離(n=100)Tab.3 The Euclidean distance of morphological traits for all experimental groups
2.4 實(shí)驗(yàn)測(cè)定對(duì)象形態(tài)比例特征的主成分分析
經(jīng)Bartlett球形檢驗(yàn)和KMO適合度檢驗(yàn)發(fā)現(xiàn), 本研究所涉形態(tài)比例性狀相關(guān)系數(shù)矩陣與單位陣具顯著差異(P<0.05)且適合度尚可(KMO=0.767>0.700),表明有做主成分分析的必要性。
由表4可見, 所列5個(gè)主成分的特征值均大于1且方差累計(jì)貢獻(xiàn)率達(dá)80.795%, 故可基本認(rèn)定它們?yōu)槟芨爬ū狙芯克嫒毡灸覍?duì)蝦秋繁同生群仔蝦不同抗水流性能實(shí)驗(yàn)群體間形態(tài)比例特征差異的公共因子。其中, PC1的方差貢獻(xiàn)率最大(39.561%)且遠(yuǎn)高于其它各主成分。將載荷絕對(duì)值P>0.5的變量確定為主要影響變量, 則 PC1、PC2、PC3、PC4、PC5所含主要變量的個(gè)數(shù)依次為8、4、3、3和1, 除PC5僅有的1個(gè)主要變量載荷高達(dá)0.957外, 其余8個(gè)載荷絕對(duì)值達(dá)到0.80以上的主要變量均存在于PC1中。無疑, PC1在區(qū)分的不同抗水流性能實(shí)驗(yàn)群體間的體型差異上具重要作用。繪制PC1與其它各主成分間的得分散布圖, 得圖3。由圖3可見, 雖然A、B、C、D、E實(shí)驗(yàn)測(cè)定樣本沿FAC1軸呈由右向左依次排列, 但抗流性能相鄰及相間的兩實(shí)驗(yàn)群體之間均存在較大程度的重疊, 表明PC1在反映本研究所涉全部實(shí)驗(yàn)個(gè)體的類群歸屬問題上受到了來自自身及其它主成分主要影響變量的干擾。
2.5 水流脅迫處理淘汰群體和選留群體實(shí)驗(yàn)對(duì)象間形態(tài)比例特征的判別分析
采用逐步導(dǎo)入剔除法, 從表2所列形態(tài)比例性狀中篩選出對(duì)判別貢獻(xiàn)較大的C1、C3、C7和C13進(jìn)行判別分析, F檢驗(yàn)表明這些性狀均達(dá)到極顯著水平(P<0.01)。根據(jù)上述4個(gè)性狀建立本研究各區(qū)段秋繁仔蝦實(shí)驗(yàn)樣本的Fisher分類函數(shù)方程組于表5。經(jīng)驗(yàn)證, 選留群體實(shí)驗(yàn)樣本的判別準(zhǔn)確率P1、P2分別為98%和 84.85%, 淘汰群體實(shí)驗(yàn)樣本的判別準(zhǔn)確率P1、P2分別為 82.25%和 97.63%, 綜合判別準(zhǔn)確率為90.12% (表6)。另, 所繪制的典型判別函數(shù)判別得分散布圖(圖 4), 更直觀地印證了上述判別結(jié)果的可靠性。
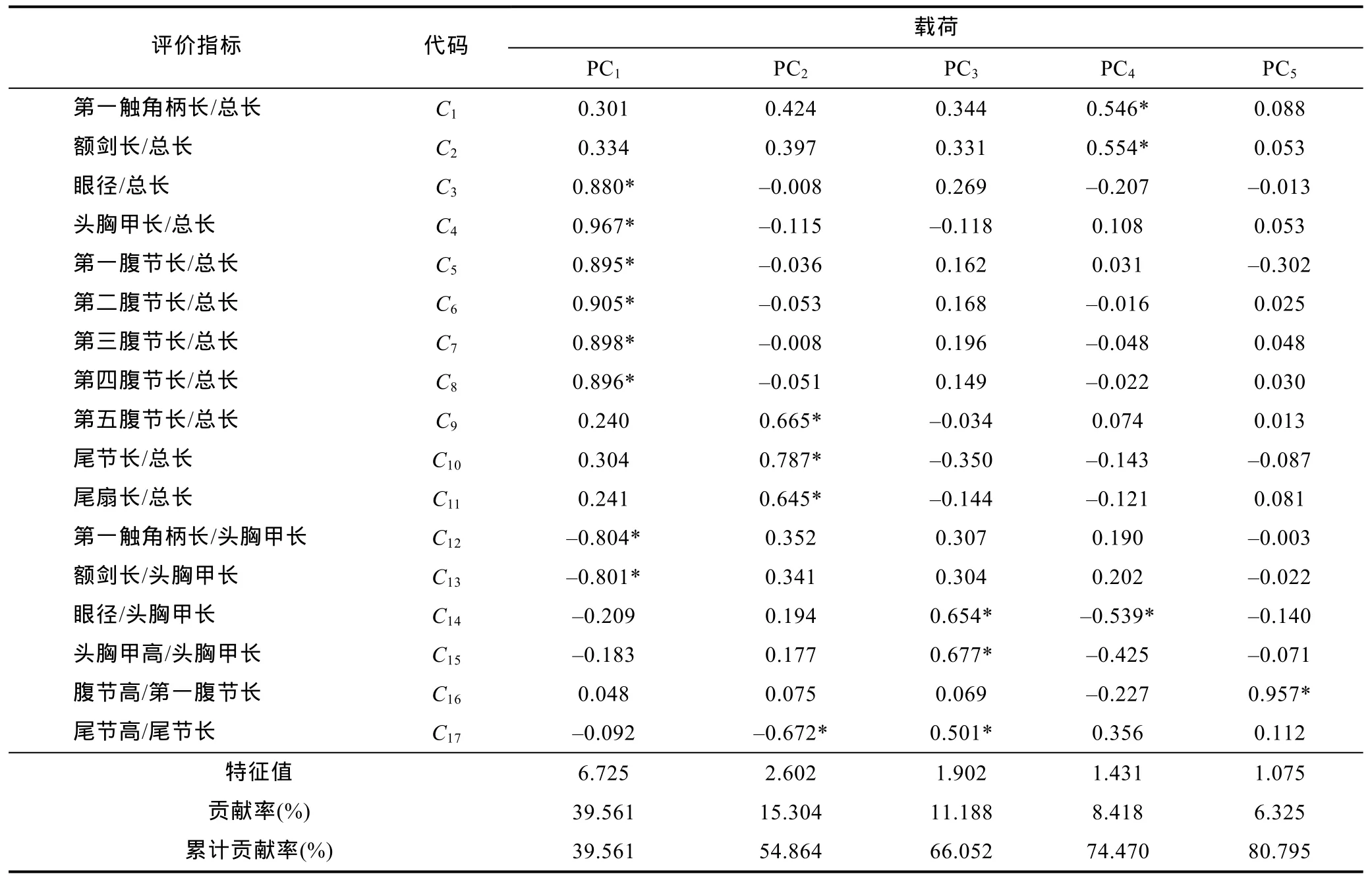
表4 實(shí)驗(yàn)群體形態(tài)比例特征的主成分分析Tab.4 The principal component analysis on morphological traits for all experimental groups
3 討論
本研究所涉各測(cè)定群體形態(tài)比例指標(biāo)均值間的歐式距離均有隨抗流性能差異的增大而呈顯著增大的趨勢(shì)(表 3), 表明形態(tài)比例特征與其抗流性能間具強(qiáng)相關(guān)性。由表4可見, 雖PC1的主要影響變量個(gè)數(shù)遠(yuǎn)多于其它主成分, 方差貢獻(xiàn)率略低于其它主成分之和, 但就FAC1得分軸上各相鄰及相間實(shí)驗(yàn)群體測(cè)定樣本間均存在較大程度的重疊(圖 2), 以及水流脅迫處理選留群體和淘汰群體分別有2%和17.75%的個(gè)體被誤判(表6, 圖3)的結(jié)果, 表明被列入諸主成分的主要影響變量和經(jīng)判別分析被選入Fisher分類函數(shù)方程組的自變量中共同包含了對(duì)區(qū)分抗流性能具高分辨作用和較強(qiáng)干擾作用的兩類關(guān)鍵變量。結(jié)合表5所列 4 個(gè)判別變量中,C3、C7均呈 B>C>D>E>A(P<0.05),C13呈 A>E>D>C>B (P<0.05), 而C1僅 C、D實(shí)驗(yàn)群體間具顯著差異(P<0.05) 的結(jié)果(表 2), 可判定上述具強(qiáng)干擾作用的關(guān)鍵變量應(yīng)為C1, 而其余3個(gè)判別變量則均為具高分辨作用的關(guān)鍵變量。
已有研究表明, 頂流運(yùn)動(dòng)會(huì)顯著增加水生動(dòng)物的能耗水平并提高機(jī)體耗氧率(Stevenet al, 1982;Wardleet al, 1996; Herskinet al, 1998; Leeet al, 2003);較為激烈的運(yùn)動(dòng)會(huì)導(dǎo)致水生動(dòng)物因氧需求量超過其供應(yīng)量而表露“氧債”, 并使其機(jī)體能量供不應(yīng)求(林浩然, 1999)。因此, 較高的流速會(huì)使頂流運(yùn)動(dòng)中供氧或供能不足的實(shí)驗(yàn)個(gè)體首先被沖離, 即實(shí)驗(yàn)個(gè)體抗流能力的強(qiáng)弱與其運(yùn)動(dòng)能量代謝能力有著極為密切的關(guān)系。無疑, 本研究中A實(shí)驗(yàn)群體不僅抗流性能與B最為接近, 與E差距最大(表3), 且在FAC1得分軸上與 B重疊程度最大, 與 E完全不重疊(圖 2), 而在判別表型上卻表露為C3、C7均呈 B>C>D>E>A(P<0.05),C13呈 A>E>D>C>B (P<0.05)的結(jié)果(表 2,圖3), 表明實(shí)驗(yàn)仔蝦存在以A、B實(shí)驗(yàn)群體為代表的兩種截然不同的運(yùn)動(dòng)能量代謝機(jī)制。
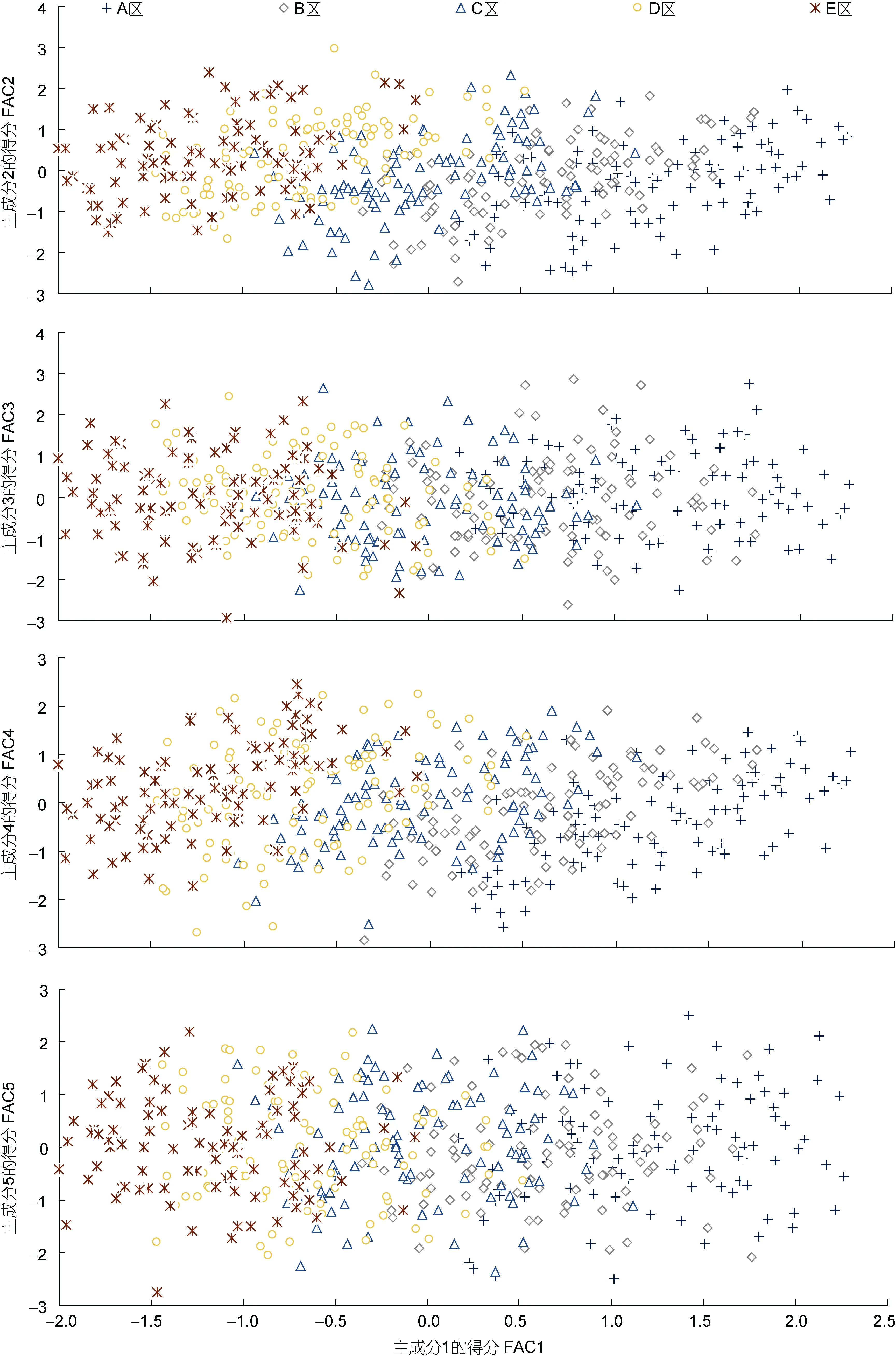
圖3 實(shí)驗(yàn)測(cè)定樣本形態(tài)比例指標(biāo)的主成分散布圖Fig.3 The scatter diagram of principal component of morphological traits for all experimental groups
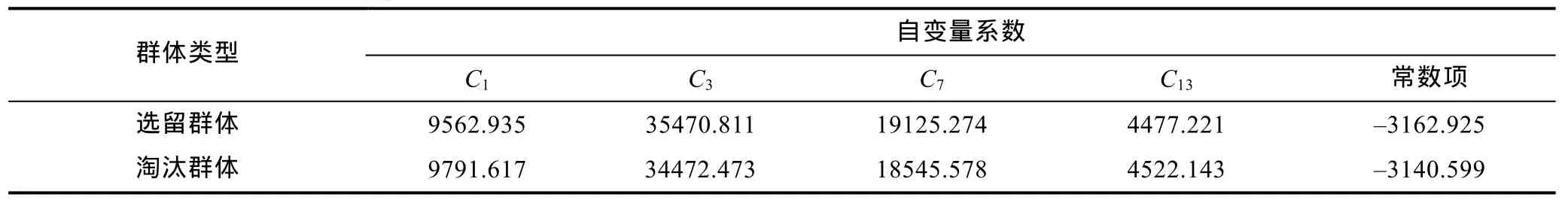
表5 選留群體和淘汰群體表型比例特征Fisher分類函數(shù)方程組自變量系數(shù)及常數(shù)項(xiàng)Tab.5 The independent variable coefficient and the constant terms in the Fisher classification function

表6 選留群體和淘汰群體實(shí)驗(yàn)樣本的判別分類結(jié)果Tab.6 The discrimination for all experimental groups
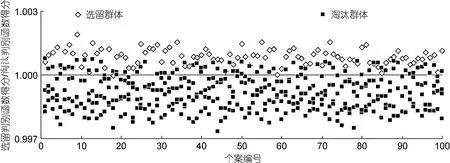
圖4 淘汰群體與選留群體實(shí)驗(yàn)樣本的典型判別函數(shù)判別得分散布圖Fig.4 The scatter diagram of typical discriminant function for all experimental groups
仔蝦處于以體型增長(zhǎng)為主和體型增粗為輔的異速生長(zhǎng)階段。據(jù)報(bào)道, 對(duì)蝦科種類的主要臟器集中于頭胸部(王吉橋等, 1999); 額劍系表征對(duì)蝦體質(zhì)和生長(zhǎng)代謝旺盛程度的重要體征(王志錚等, 2012), 具御敵攻擊和保障運(yùn)動(dòng)平衡的作用(王安利等, 1993; 沈輝等, 2010)。因此, 本研究所涉各實(shí)驗(yàn)群體的X1和C13均呈 A>E>D>C>B (P<0.05)的結(jié)果(表 1, 表 2), 表明額劍長(zhǎng)與頭胸甲之比的C13可作為表征仔蝦生長(zhǎng)代謝強(qiáng)度的重要指標(biāo)。因此, 可用于表征生長(zhǎng)速度的X1和生長(zhǎng)代謝強(qiáng)度的C13均呈A實(shí)驗(yàn)群體顯著大于B實(shí)驗(yàn)群體(P<0.05), 而可用于表征尋食視野的C3和腹部運(yùn)動(dòng)幅度的C7則均呈A實(shí)驗(yàn)群體顯著小于B實(shí)驗(yàn)群體(P<0.05)的結(jié)果(表1, 表2), 也就揭示了A實(shí)驗(yàn)群體具采取顯著提高運(yùn)動(dòng)代謝強(qiáng)度和增加捕食運(yùn)動(dòng)頻率, 以獲得快速生長(zhǎng)為特征的“高能耗速生型”運(yùn)動(dòng)能量代謝機(jī)制, B實(shí)驗(yàn)群體具采取顯著降低運(yùn)動(dòng)代謝水平以減少運(yùn)動(dòng)頻率, 和顯著擴(kuò)大尋食視野與腹部運(yùn)動(dòng)幅度以提高捕食成功率為特征的“低能耗慢長(zhǎng)型”運(yùn)動(dòng)能量代謝機(jī)制, 故前者較后者更易在較激烈的頂流運(yùn)動(dòng)中表露“氧債”。無疑, 戴習(xí)林等(2008)報(bào)道的養(yǎng)殖于水平水流 0.63—2.78mm/s范圍內(nèi)的凡納濱對(duì)蝦幼蝦體長(zhǎng)整齊度顯著優(yōu)于養(yǎng)殖于靜水狀態(tài)下的結(jié)果, 也充分支持了上述判斷的可靠性, 即水流作用使采取“高能耗速生型”運(yùn)動(dòng)能量代謝機(jī)制的幼蝦, 因能耗明顯增加致使其體長(zhǎng)增速有所減緩, 而采取“低能耗慢長(zhǎng)型”運(yùn)動(dòng)能量代謝機(jī)制的幼蝦, 則因供餌條件明顯改善致使其體長(zhǎng)增速有所提高, 從而導(dǎo)致上述兩類運(yùn)動(dòng)能量代謝機(jī)制的幼蝦的體長(zhǎng)更加逼近,整齊度獲得顯著提高。顯然, A實(shí)驗(yàn)群體被選留的原因, 系本研究中所設(shè)實(shí)驗(yàn)周期下的實(shí)驗(yàn)流速接近或低于 A實(shí)驗(yàn)群體頂流運(yùn)動(dòng)的能耗可容忍臨界所致,而B、C、D、E實(shí)驗(yàn)群體, 則因該實(shí)驗(yàn)流速已超過它們頂流運(yùn)動(dòng)的能耗可容忍臨界, 致使它們隨生長(zhǎng)速度和頂流能耗的增大而首先被沖離。綜上可知,C3、C7、C13成為對(duì)區(qū)分抗流性能具高分辨作用的關(guān)鍵變量的主因, 為這些關(guān)鍵判別變量組合可真實(shí)反映所涉實(shí)驗(yàn)群體間運(yùn)動(dòng)能量代謝能力的差異; 仔蝦第一觸角柄第一節(jié)基部具平衡囊, 囊內(nèi)有砂粒, 具司體位及姿態(tài)平衡的作用, 第三節(jié)末端的觸鞭被認(rèn)為是觸覺感受器官(王克行, 1997), 故C17成為對(duì)區(qū)分抗流性能具較強(qiáng)干擾作用的關(guān)鍵變量的主因, 無疑與其含形態(tài)性狀信息在維系仔蝦運(yùn)動(dòng)平衡中所起的具重要作用有關(guān)。
王吉橋, 靳翠麗, 姜靜穎等, 1999. 中國對(duì)蝦和日本對(duì)蝦身體不同部位生長(zhǎng)的研究. 見: 中國動(dòng)物科學(xué)研究——中國動(dòng)物學(xué)會(huì)第十四屆會(huì)員代表大會(huì)及中國動(dòng)物學(xué)會(huì)65周年年會(huì)論文集. 鄭州: 中國動(dòng)物學(xué)會(huì), 92—98
王安利, 母學(xué)全, 周世梅等, 1993. 中國對(duì)蝦行為的觀察. 海洋科學(xué), 17(4): 16—18
王志錚, 楊 磊, 施建軍等, 2012. 日本沼蝦(Macrobrachium nipponensis)池養(yǎng)越冬抱卵親蝦表型性狀對(duì)個(gè)體繁殖力的影響效應(yīng). 海洋與湖沼, 43(1): 166—173
王克行, 1997. 蝦蟹類增養(yǎng)殖學(xué). 北京: 中國農(nóng)業(yè)出版社, 7—8,31
中華人民共和國農(nóng)業(yè)部, 2001. NY 5052-2001無公害食品海水養(yǎng)殖用水水質(zhì). 北京: 中國農(nóng)業(yè)出版社, 1—5
楊勁峰, 陳 清, 韓曉日等, 2002. 數(shù)字圖像處理技術(shù)在蔬菜葉面積測(cè)量中的應(yīng)用. 農(nóng)業(yè)工程學(xué)報(bào), 18(4): 155—158
沈 輝, 萬夕和, 許 璞等, 2010. 脊尾白蝦的行為學(xué)觀察研究. 海洋科學(xué), 34(10): 53—56
林浩然, 1999. 魚類生理學(xué). 廣州: 廣東高等教育出版社, 57
蓋均鎰, 2000. 試驗(yàn)統(tǒng)計(jì)方法. 北京: 中國農(nóng)業(yè)出版社, 49
戴習(xí)林, 臧維玲, 張 韜, 2008. 水流對(duì)凡納濱對(duì)蝦幼蝦生長(zhǎng)與存活的影響. 上海海洋大學(xué)學(xué)報(bào), 17(1): 52—57
Dall W, Hill B J, Rothlisberg P C等編著, 陳楠生, 李新正, 劉恒等譯, 1992. 對(duì)蝦生物學(xué). 青島: 青島海洋大學(xué)出版社,358—379
Haas T C, Blum M J, Heins D C, 2010. Morphological responses of a stream fish to water impoundment. Biology Letters, 6(6):803—806
Herskin J, Steffensen J F, 1998. Energy saving in sea bass swimming in a school: measurements of tail beat frequency and oxygen consumption at different swimming speeds.Journal of Fish Biology 53(2): 366—376
Langerhans R B, Layman C A, Langerhans A K et al, 2003.Habitat-associated morphological divergence in two neotropical fish species. Biological Journal of the Linnean Society, 80(4): 689—698
Lee C G, Devlin R H, Farrell A P, 2003. Swimming performance,oxygen consumption and excess post-exercise oxygen consumption in adult transgenic and ocean-ranched coho salmon. Journal of Fish Biology, 62(4): 753—766
Li S F, Wang C H, Cheng Q Q, 2005. Morphological variations and phylogenesis of four strains in Cyprinus carpio. Journal of Fisheries of China, 29(5): 606—611
Stevens D E, Dizon A E, 1982. Energetic of locomotion in warm-bodied fish. Annual Review of Physiology, 44:121—131
Wardle C S, Soofiani N M, O’Neill F G et al, 1996.Measurements of aerobic metabolism of a school of horse mackerel at different swimming speeds. Journal of Fish Biology, 49(5): 854—862
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
音樂探索(2022年2期)2022-05-30 21:01:37
中學(xué)生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
小天使·一年級(jí)語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
鄭州大學(xué)學(xué)報(bào)(醫(yī)學(xué)版)(2015年2期)2015-02-27 14:50:46

- 海洋與湖沼的其它文章
- 烏賊墨黑色素對(duì)鉛中毒小鼠的排鉛作用及其對(duì)體內(nèi)鈣、鋅和銅含量的影響*
- 感染溶藻弧菌及白斑綜合癥病毒后凡納濱對(duì)蝦不同組織的Toll樣受體基因表達(dá)變化研究*
- 南沙群島西南部陸架區(qū)底層魚類營(yíng)養(yǎng)結(jié)構(gòu)研究*
- 無機(jī)砷As(V)對(duì)牟氏角毛藻(Chaetoceros mulleri)生長(zhǎng)、葉綠素a含量及抗氧化活力的影響*
- 海捕大管鞭蝦(Solenocera melantho)調(diào)理食品在常溫保藏中的貨架期預(yù)測(cè)及產(chǎn)品質(zhì)量分析*
- 十溴聯(lián)苯醚(BDE-209)脅迫對(duì)褶皺臂尾輪蟲(Brachionus plicatilis)攝食與選食的影響*