基于海馬神經(jīng)元組群的癲癇仿真模型研究進(jìn)展
2016-02-04 01:13:52鄭茜茜陳亮亮葉學(xué)松
溫州醫(yī)科大學(xué)學(xué)報 2016年12期
關(guān)鍵詞:癲癇
鄭茜茜,陳亮亮,葉學(xué)松
(1.溫州醫(yī)科大學(xué) 生物醫(yī)學(xué)工程學(xué)院,浙江 溫州 325035;2.浙江大學(xué) 生物醫(yī)學(xué)工程與儀器科學(xué)學(xué)院,浙江 杭州 310027)
?綜 述?
基于海馬神經(jīng)元組群的癲癇仿真模型研究進(jìn)展
鄭茜茜1,陳亮亮1,葉學(xué)松2
(1.溫州醫(yī)科大學(xué) 生物醫(yī)學(xué)工程學(xué)院,浙江 溫州 325035;2.浙江大學(xué) 生物醫(yī)學(xué)工程與儀器科學(xué)學(xué)院,浙江 杭州 310027)
癲癇是一種嚴(yán)重危害人類健康的神經(jīng)系統(tǒng)疾病,對此研究者們投入了大量的財力、人力和物力研究其發(fā)病機(jī)制。在實驗基礎(chǔ)上建立癲癇仿真模型有利于人們探索癲癇發(fā)病機(jī)制及研制抗癲癇藥物。大量研究表明癲癇的發(fā)生與海馬結(jié)構(gòu)變化有密切關(guān)系,因此本文介紹了近年來國際上廣泛研究的、基于生理解剖的海馬神經(jīng)元組群的癲癇仿真模型的研究工作,并結(jié)合我們目前的工作展望癲癇仿真模型的發(fā)展趨勢。[關(guān)鍵詞]癲癇;仿真模型;海馬CA1區(qū);海馬CA3區(qū);綜述文獻(xiàn)
癲癇是大腦神經(jīng)元高度超同步化造成突發(fā)性異常放電的一種臨床現(xiàn)象,其發(fā)病率在神經(jīng)系統(tǒng)疾病中僅次于腦血管病,是一種較為常見的慢性疾病。癲癇的發(fā)作具有不可預(yù)見性、反復(fù)性和突發(fā)性的特點,發(fā)作時對人體的傷害很大,尤其對腦部的損傷最大。目前臨床上主要通過藥物或者外科手術(shù)來治療癲癇患者。這兩類方法對許多癲癇患者具有良好的治療效果,但仍有約25%的患者不能得到有效治療,稱為難治性癲癇[1]。迄今為止,還不能完全有效地抑制癲癇發(fā)作,主要原因就在于人們對癲癇的發(fā)病機(jī)制尚未完全闡明。
難治性癲癇中絕大部分為顳葉癲癇(temporal lobe epilepsy,TLE)。大量研究表明TLE的發(fā)生可能與海馬和邊緣系統(tǒng)之間復(fù)雜的興奮性連接環(huán)路[2]以及海馬結(jié)構(gòu)變化[3]有關(guān),海馬內(nèi)的神經(jīng)網(wǎng)絡(luò)和神經(jīng)元功能異常是癲癇發(fā)生的重要因素,因此目前對癲癇發(fā)病機(jī)制探索比較多的是以海馬神經(jīng)元組群作為研究對象。但是由于海馬神經(jīng)元系統(tǒng)具有復(fù)雜的形態(tài)結(jié)構(gòu),為直接的實驗研究帶來了不少困難;但是實驗的定性研究與模型的定量分析相結(jié)合,卻為研究癲癇發(fā)病機(jī)制提供了一種行之有效的新方法。據(jù)此,本文就海馬神經(jīng)元組群的解剖結(jié)構(gòu)及癲癇仿真模型的研究現(xiàn)狀作一綜述。
1 海馬與TLE
1.1 海馬神經(jīng)元組群解剖結(jié)構(gòu) 海馬是組成大腦邊緣系統(tǒng)的一個重要部分,包括海馬體(hippocampus,CA)、齒狀回(dentate gyrus,DG)、下托(subiculum)、前下托(presubiculum,PrS)、傍下托(parasubiculum,PaS)、內(nèi)嗅皮層(entorhinal cortex,EC)。海馬CA區(qū)根據(jù)各區(qū)錐體(pyramidal,P)細(xì)胞形態(tài)和纖維排列層次的細(xì)致差異分為4個區(qū):CA1、CA2、CA3和CA4[4]。海馬解剖結(jié)構(gòu)和“三突觸回路”見圖1。三突觸回路是指:神經(jīng)信號通過EC經(jīng)前穿質(zhì)通路(perforant pathway,PP)投射至DG上的顆粒細(xì)胞層;DG顆粒細(xì)胞層上的神經(jīng)信號通過苔狀纖維(mossy fibers,MF)傳遞到位于CA3區(qū)的P細(xì)胞層;CA3區(qū)P細(xì)胞層上的神經(jīng)信號通過Schaffer側(cè)枝(schaffer collaterals,SC)傳向CAl區(qū)。海馬中包含很多類型的神經(jīng)元,主要分為2類:興奮性和抑制性。興奮性神經(jīng)元是海馬中存在的主要細(xì)胞,包括海馬區(qū)的P細(xì)胞、DG層的顆粒細(xì)胞。抑制性神經(jīng)元又叫中間神經(jīng)元[5],主要包括籃狀(basket,B)細(xì)胞、始層分子層(oriens-lacunosum moleculare,OLM)細(xì)胞、枝形細(xì)胞、軸軸(axo-axonic,AA)細(xì)胞、雙尖(bistratified,BS)細(xì)胞等,約占海馬神經(jīng)元總數(shù)的10%。
1.2 海馬與TLE 研究表明:通過對內(nèi)側(cè)TLE患者的海馬組織進(jìn)行磁共振成像(magnetic resonance imaging,MRI)和磁共振波譜(magnetic resonance spectroscopy,MRS)成像分析,發(fā)現(xiàn)其海馬出現(xiàn)硬化現(xiàn)象[6];從TLE患者切除的海馬標(biāo)本中可以發(fā)現(xiàn),海馬出現(xiàn)萎縮現(xiàn)象[7]。普遍認(rèn)為癲癇發(fā)作是由神經(jīng)元集群的放電行為導(dǎo)致的,其誘發(fā)因素有很多,比如神經(jīng)元的放電特性,突觸連接的類型、強(qiáng)度、可塑性以及神經(jīng)元集群連接的拓?fù)浣Y(jié)構(gòu)等。海馬4個區(qū)中CA2只占一個很小的部分,有學(xué)者認(rèn)為CA4區(qū)可能是DG的一部分,因此CA1區(qū)與CA3區(qū)構(gòu)成嚴(yán)格意義上的海馬,多數(shù)關(guān)于癲癇的研究都是以它們作為研究對象[8-12],CA2區(qū)與CA4區(qū)通常忽略不計。
癲癇發(fā)作在腦電圖上比較常見的表現(xiàn)為癲癇樣放電,分為間歇樣放電和發(fā)作樣放電。間歇樣放電表現(xiàn)為時程較短的棘波,一般時程只有幾百毫秒;發(fā)作樣放電表現(xiàn)為長時程(高達(dá)幾十秒)的放電[13]。
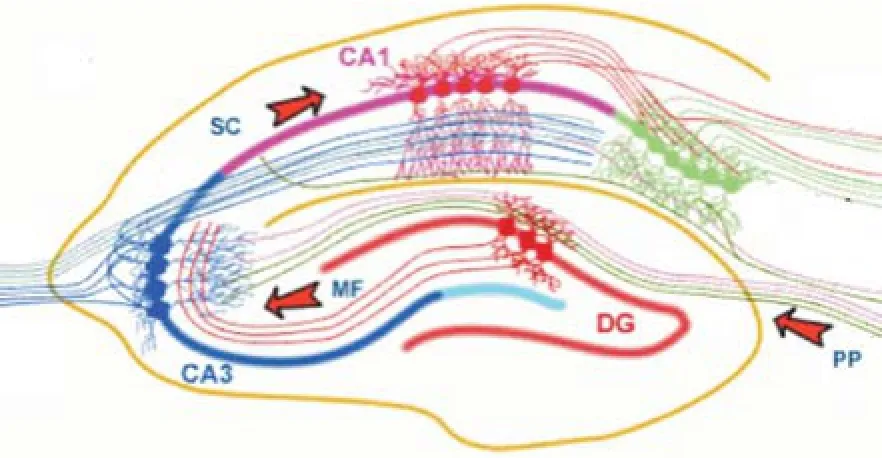
圖1 海馬解剖結(jié)構(gòu)和“三突觸回路”
2 癲癇仿真模型
為了有效、迅速地篩選和研制抗癲癇藥物,首先要建立癲癇模型。目前比較成熟的動物實驗?zāi)P椭饕械玩V和4-氨基吡啶(4-aminopyridine,4-AP)模型,但是這些模型很難解釋癲癇的發(fā)作機(jī)制,極大程度地限制了癲癇預(yù)測及抑制治療的研究進(jìn)展。比如:EC也是大腦邊緣系統(tǒng)一個重要組成部分,研究表明EC與TLE的產(chǎn)生和維持有密切的聯(lián)系。但是動物實驗中,很難只改變網(wǎng)絡(luò)中EC對海馬的作用。因此需要借助計算機(jī)仿真模型,在細(xì)胞水平上闡明癲癇發(fā)作的內(nèi)在機(jī)制。
2.1 海馬CA1區(qū)癲癇仿真模型 為了研究θ節(jié)律(4-7 Hz)在記憶編碼和檢索過程中起的作用,Cutsuridis等[14]構(gòu)建了海馬CA1區(qū)仿真模型,見圖2a。網(wǎng)絡(luò)模型包含P細(xì)胞(100個)和4種類型的中間神經(jīng)元(B、BS、AA和OLM細(xì)胞)。對于P細(xì)胞,輸入來自EC區(qū)和海馬CA3區(qū)的SC。每個中間神經(jīng)元都與P細(xì)胞通過突觸形成抑制性連接。汪洋[15]在此模型基礎(chǔ)上建立了CA1區(qū)癲癇仿真模型,來研究體感刺激對于癲癇發(fā)作時神經(jīng)活動的影響。為了模擬癲癇發(fā)作,通過直接施加脈沖刺激到CA1區(qū)部分P細(xì)胞上獲得仿真癇樣波。改變網(wǎng)絡(luò)中EC輸入突觸連接權(quán)重來模擬真實情況下體感刺激強(qiáng)度的改變。仿真結(jié)果顯示,對于海馬單側(cè)的癲癇活動,體感刺激有抑制作用,但對于雙側(cè)影響不大。
為了模擬癲癇發(fā)作時的間歇樣放電,Ratnadurai-Giridharan等[16]利用H-H方程建立了海馬CA1區(qū)癲癇模型,見圖2b。模型中有225個P細(xì)胞,22 個B細(xì)胞,22個海馬槽(orien alveus,OA)細(xì)胞。AMPA和GABAa電導(dǎo)用來模擬突觸的相互作用。模型很好地模擬了海馬CA1區(qū)自發(fā)的間歇樣放電,即使SC輸入不是完全同步。
Wei等[17]同樣利用H-H方程構(gòu)建了海馬CA1區(qū)生物物理模型,用來仿真氧代謝和癲癇發(fā)作之間的關(guān)系,進(jìn)一步解釋了實驗中所觀察到缺氧可以誘導(dǎo)癲癇發(fā)作這一現(xiàn)象,從而對癲癇發(fā)作時離子動力學(xué)和能量代謝之間復(fù)雜的相互關(guān)系有了一個更全面的了解。
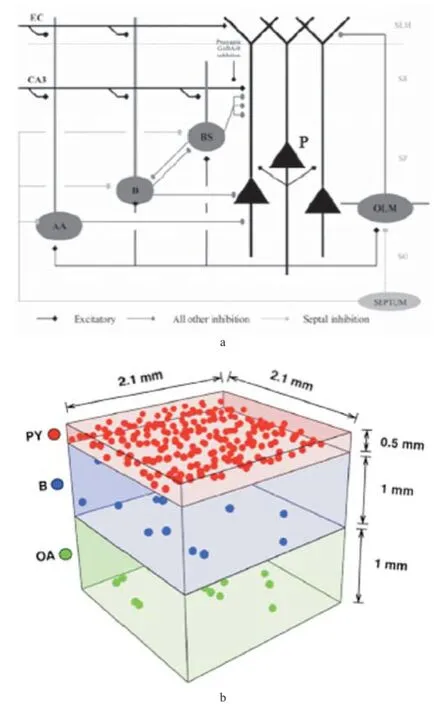
圖2 海馬CA1區(qū)仿真模型
2.2 海馬CA3區(qū)癲癇仿真模型 海馬CA3區(qū)是信號傳遞的樞紐,對于癲癇的產(chǎn)生和擴(kuò)散有著重要的作用。關(guān)于海馬CA3區(qū)仿真模型,也有不少文獻(xiàn)進(jìn)行了報道。Kunec等[18]根據(jù)生理數(shù)據(jù)搭建了如圖3a所示的海馬CA3區(qū)網(wǎng)絡(luò)模型,模型中的主要細(xì)胞包括P細(xì)胞和2種類型的中間神經(jīng)元(B細(xì)胞和OLM細(xì)胞),網(wǎng)絡(luò)的輸入來自鼻中隔和EC。通過改變網(wǎng)絡(luò)中的突觸連接,模擬記憶在編碼和檢索過程中和信號在網(wǎng)絡(luò)中的傳遞。仿真結(jié)果表明:在記憶編碼期間,EC對海馬區(qū)的信號輸入相對較強(qiáng),網(wǎng)絡(luò)中主要細(xì)胞之間的突觸連接變強(qiáng);在記憶檢索期間,來自EC區(qū)的信號通過DG激活之前加強(qiáng)的突觸連接。
Neymotin等[19]建立了如圖3b所示海馬CA3區(qū)網(wǎng)絡(luò)模型,用來研究氯胺酮誘發(fā)精神病的一些可能機(jī)制。模型中包括800個P細(xì)胞、200個B細(xì)胞和200 個OLM細(xì)胞。在模型中,通過阻斷網(wǎng)絡(luò)中不同細(xì)胞上的NMDA受體通道來模擬精神病患者的異常放電,仿真結(jié)果表明氯胺酮可能主要是阻斷了OLM細(xì)胞上的NMDA受體從而誘發(fā)了精神病。這些模型為癲癇仿真模型的建立奠定了良好的理論基礎(chǔ)。
Traub等[20]通過建立海馬CA3區(qū)癲癇仿真模型,研究低鎂人工腦脊液(artificial cerebrospinal fluid,ACSF)誘發(fā)海馬腦片產(chǎn)生癲癇樣放電的可能機(jī)制。該模型包含1 000個P細(xì)胞和100個中間神經(jīng)元,模型沒有考慮中間神經(jīng)元的種類,只建立了一類抑制性細(xì)胞,代表所有中間神經(jīng)元。利用仿真模型研究當(dāng)胞外鎂離子濃度不同時,某一P細(xì)胞的放電情況以及網(wǎng)絡(luò)中同步性放電的神經(jīng)元個數(shù)。研究發(fā)現(xiàn)當(dāng)鎂離子濃度為0 mmol/L時,網(wǎng)絡(luò)中大量神經(jīng)元同步性放電,顯示出癲癇樣放電。
Ren等[21]為了研究EC對海馬癲癇樣放電的影響,簡化了Traub等建立的海馬CA3區(qū)網(wǎng)絡(luò)模型,見圖3c,網(wǎng)絡(luò)中只包括2個P細(xì)胞、1個B細(xì)胞和1個OLM細(xì)胞。根據(jù)生理解剖學(xué)構(gòu)建神經(jīng)元之間的突觸連接,網(wǎng)絡(luò)還接受2個外界輸入:DG和EC的興奮性輸入。仿真結(jié)果表明,改變網(wǎng)絡(luò)中的鎂離子濃度至0 mmol/L,海馬CA3區(qū)產(chǎn)生間歇樣放電,如果給網(wǎng)絡(luò)加入EC信號輸入,網(wǎng)絡(luò)能產(chǎn)生發(fā)作樣放電。同時通過增強(qiáng)抑制性γ-氨基丁酸(γ-aminobutyric acid,GABA)能連接強(qiáng)度和降低興奮性N-甲基-D-天門冬氨酸(N-methyl-D-aspartate,NMDA)能連接強(qiáng)度,來模擬一種抗癲癇藥物—丙戊酸(valproate,VPA)對海馬間歇期放電的影響。結(jié)果顯示可能是由于VPA阻斷了EC的發(fā)作樣放電,從而能夠阻斷EC-海馬聯(lián)合腦片上的發(fā)作樣放電。
乙酰膽堿通過誘導(dǎo)海馬在分離和完成2個模式之間切換來調(diào)節(jié)記憶的編碼和檢索,這2個過程會引起網(wǎng)絡(luò)活動的大量變化,從而可能會導(dǎo)致癲癇樣興奮的傳播。因此在一定范圍內(nèi)保持網(wǎng)絡(luò)的穩(wěn)定性是必須的。Hummos等[22]建立了如圖3d所示的海馬體EC-DG-CA3區(qū)仿真模型,用來研究在記憶的編碼和檢索期間,海馬體CA3區(qū)保持網(wǎng)絡(luò)動態(tài)穩(wěn)定的內(nèi)在機(jī)制,從而抑制癲癇波的傳播。
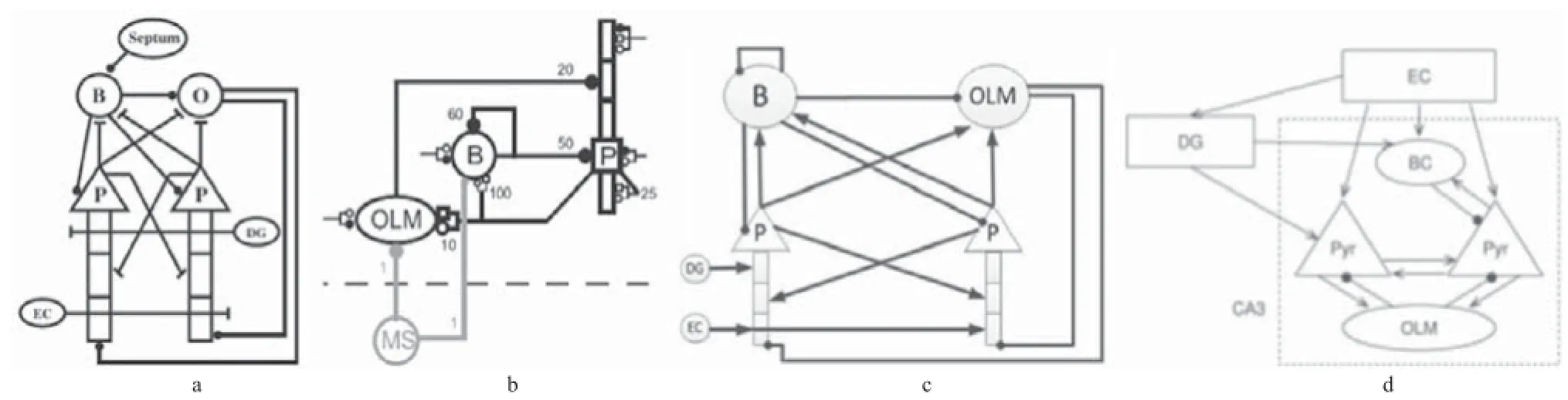
圖3 海馬CA3區(qū)仿真模型
2.3 海馬開、閉環(huán)回路仿真模型 最近的實驗研究[23]表明:EC層和海馬組群之間傳導(dǎo)的時間延遲要比預(yù)期的長,為70~80 ms。Cutsuridis等[24]通過構(gòu)建DG、CA3區(qū)和CA1區(qū)開環(huán)回路仿真模型,研究這種時間延遲的內(nèi)在機(jī)制,見圖4。模型中主要包括顆粒細(xì)胞、CA3區(qū)和CA1區(qū)的P細(xì)胞和6種類型的中間神經(jīng)元(B細(xì)胞、AA細(xì)胞、BS細(xì)胞、OLM細(xì)胞、苔蘚細(xì)胞和前通路門細(xì)胞)。網(wǎng)絡(luò)的輸入來自EC和內(nèi)側(cè)隔。仿真結(jié)果顯示這種延遲主要是由于θ振蕩引起的胞體和突觸抑制性電位的存在。
在此基礎(chǔ)上,Sandler等[25]構(gòu)建了海馬體閉環(huán)回路模型,信號從CA1區(qū)再反饋到EC。利用該模型研究癲癇振蕩的產(chǎn)生和設(shè)計有效的神經(jīng)刺激模式來消除這種振蕩。深部腦刺激(deep brain stimulation,DBS)是一種很有前途的抑制癲癇發(fā)作的治療手段,但是現(xiàn)有的刺激模式多為開環(huán)的,不能根據(jù)患者的實際情況自動改變刺激模式,選擇最佳的DBS頻率參數(shù)。基于該閉環(huán)仿真模型的研究可以被用來作為一個實驗平臺,針對每個患者的實際情況設(shè)計一個最佳的DBS模式。盡管該模型還在驗證階段,但是也解釋了一些在實驗研究中出現(xiàn)、未解決的現(xiàn)象。比如:不同的刺激頻率都可以抑制同一系統(tǒng)內(nèi)的癲癇發(fā)作,這和Kinoshita等[26]發(fā)現(xiàn)的實驗現(xiàn)象(1 Hz和50 Hz的刺激對于TLE患者具有相同的抗驚厥效果)相符合。
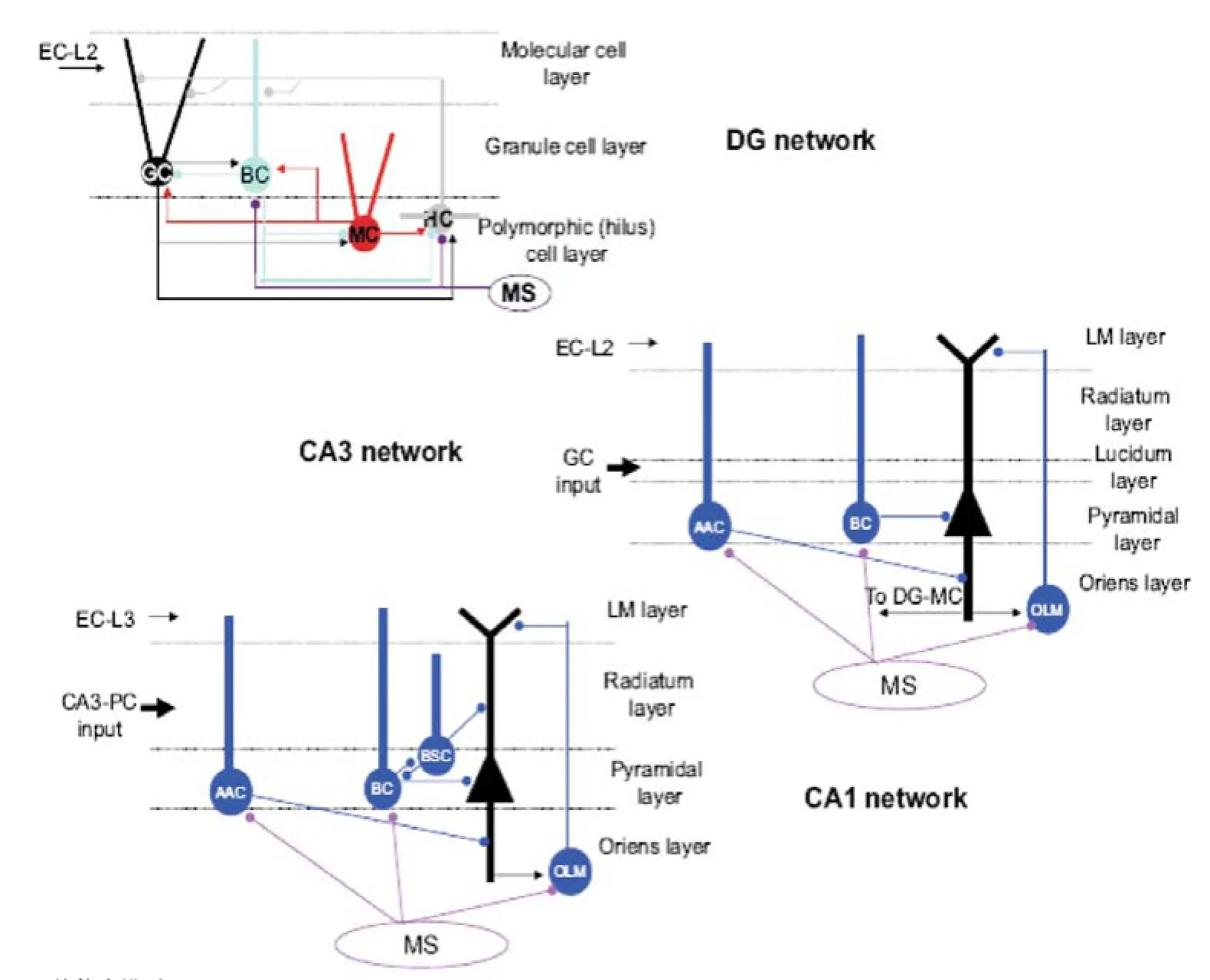
圖4 DG-CA3-CA1網(wǎng)絡(luò)仿真模型
3 展望
根據(jù)目前的研究,人們利用各種癲癇仿真模型,對癲癇的發(fā)病機(jī)制以及癲癇的抑制研究取得了較大的進(jìn)步,但這些研究并沒有以大量的實驗數(shù)據(jù)作為支持。針對以上問題,我們實驗室提出了基于微流控和光解籠鎖谷氨酸的體外神經(jīng)元網(wǎng)絡(luò)癇性模型建立的研究課題。目前認(rèn)為谷氨酸與癲癇的發(fā)作關(guān)系密切[27-28],谷氨酸與其受體結(jié)合會引發(fā)癲癇,導(dǎo)致腦神經(jīng)元受到不同程度的損傷,甚至是不可逆的。其主要工作有:①搭建基于微流控技術(shù)的微電極陣列(micro electrode array,MEA)電信號記錄系統(tǒng):利用微流控技術(shù),得到具有適合海馬細(xì)胞生長的微腔和微通道的微流控芯片。在癲癇的生理解剖基礎(chǔ)上,海馬神經(jīng)元細(xì)胞在MEA芯片上培養(yǎng)和定向生長。陣列式光刺激掃描系統(tǒng)使用特定波長的光照射籠鎖谷氨酸,解除光敏基團(tuán)的籠鎖作用,在光照的特定部位(如神經(jīng)元胞體、神經(jīng)突觸等局部位置)釋放出谷氨酸。研究不同拓?fù)溥B接結(jié)構(gòu)的細(xì)胞網(wǎng)絡(luò)在谷氨酸作用下的放電情況,闡明網(wǎng)絡(luò)結(jié)構(gòu)、突觸連接與癲癇發(fā)作的內(nèi)在機(jī)制。②建立海馬神經(jīng)元網(wǎng)絡(luò)仿真模型:根據(jù)海馬神經(jīng)系統(tǒng)的解剖結(jié)構(gòu),首先建立單個神經(jīng)元模型,包括P細(xì)胞和中間神經(jīng)元;然后建立各個神經(jīng)元之間的突觸連接(興奮性和抑制性突觸),形成神經(jīng)元網(wǎng)絡(luò)仿真模型。通過將仿真結(jié)果與實驗數(shù)據(jù)相比較,優(yōu)化網(wǎng)絡(luò)結(jié)構(gòu)和參數(shù),獲得與實際觀察相符合的網(wǎng)絡(luò)模型,在細(xì)胞水平上闡明癲癇發(fā)作的內(nèi)在機(jī)制。
[1]LITT B, ECHAUZ J.Prediction of epileptic seizures[J].Lancet Neurol, 2002, 1(1): 22-30.
[2]OIKAWA H, SASAKI M, TAMAKAWA Y, et al.The circuit of Papez in mesial temporal sclerosis: MRI[J].Neuroradiology, 2001, 43(3): 205-210.
[3]HANEEF Z, LENARTOWICZ A, YEH H J, et al.Functional connectivity of hippocampal networks in temporal lobe epilepsy[J].Epilepsia, 2014, 55(1): 137-145.
[4]ANDERSON P, MORRIS R, AMARAL D, et al.The hippocampus book[M].New York and London: Oxford University Press, 2007: 37-43.
[5]FREUND T F, BUZSAKI G.Interneurons of the hippocampus[J].Hippocampus, 1996, 6(4): 347-470.
[6]岳偉, 張雅靜, 管雅琳, 等.內(nèi)側(cè)顳葉癲癇患者海馬硬化的影像學(xué)研究[J].中國神經(jīng)精神疾病雜志, 2014, 40(10): 607-611.
[7]BERTRAM E H.Temporal lobe epilepsy: Where do the seizures really begin?[J].Epilepsy Behav, 2009, 14 Suppl 1: 32-37.
[8]YANG F, GONG X W, GONG H Q, et al.Microelectrode array recordings of excitability of low Mg2+-induced acute hippocampal slices[J].Neural Regen Res, 2010, 5(20): 1548-1551.
[9]尤竹燕, 王斌生, 解敏, 等.白藜蘆醇對大鼠海馬CA1區(qū)誘發(fā)癲癇樣放電的影響[J].中國藥理學(xué)通報, 2012, 28(2): 260-265.
[10]HSIAO M C, YU P N, SONG D, et al.An in vitro seizure model from human hippocampal slices using multi-electrode arrays[J].J Neurosci Methods, 2015, 244: 154-163.
[11]ANTONIO L L, ANDERSON M L, ANGAMO E A, et al.In vitro seizure like events and changes in ionic concentration[J].J Neurosci Methods, 2016, 260: 33-44.
[12]SAFFARZADEH F, ESLAMIZADE M J, MOUSAVI S M, et al.TRPV1 receptors augment basal synaptic transmission in CA1 and CA3 pyramidal neurons in epilepsy[J].Neuroscience, 2016, 314: 170-178.
[13]MCCORMICK D A, CONTRERAS D.On the cellular and network bases of epileptic seizures[J].Annu Rev Physiol, 2001, 63: 815-846.
[14]CUTSURIDIS V, COBB S, GRAHAM B P.Encoding and retrieval in a model of the hippocampal CA1 microcircuit[J].Hippocampus, 2010, 20(3): 423-446.
[15]汪洋.大鼠海馬CA1區(qū)神經(jīng)活動對體感刺激響應(yīng)的研究[D].浙江: 浙江大學(xué), 2014.
[16]RATNADURAI-GIRIDHARAN S, STEFANESCU R A, KHARGONEKAR P P, et al.Genesis of interictal spikes in the CA1: A computational investigation[J].Front Neural Circuits, 2014, 8: 2.
[17]WEI Y, ULLAH G, INGRAM J, et al.Oxygen and seizure dynamics: II.Computational modeling[J].J Neurophysiol, 2014, 112 (2): 213-223.
[18]KUNEC S, HASSELMO M E, KOPELL N.Encoding and retrieval in the CA3 region of the hippocampus: a model of theta-phase separation[J].J Neurophysiol, 2005, 94(1): 70-82.
[19]NEYMOTIN S A, LAZAREWICZ M T, SHERIF M, et al.Ketamine disrupts theta modulation of gamma in a computer model of hippocampus[J].J Neurosci, 2011, 31(32): 11733-11743.
[20]TRAUB R D, JEFFERYS J G R, WHITTINGTON M A.Enhanced NMDA conductance can account for epileptiform activity induced by low Mg2+in the rat hippocampal slice [J].J Physiol, 1994, 478Pt3: 379-393.
[21]REN H, SHI Y J, LU Q C, et al.The role of the entorhinal cortex in epileptiform activities of the hippocampus[J].Theor Biol Med Model, 2014, 11: 14.
[22]HUMMOS A, FRANKLIN C C, NAIR S S.Intrinsic mechanisms stabilize encoding and retrieval circuits differentially in a hippocampal network model[J].Hippocampus, 2014, 24 (12): 1430-1448.
[23]MIZUSEKI K, SIROTA A, PASTALKOVA E, et al.Theta oscillations provide temporal windows for local circuit computation in the entorhinal-hippocampal loop[J].Neuron, 2009, 64(2): 267-280.
[24]CUTSURIDIS V, POIRAZI P.A computational study on how theta modulated inhibition can account for the long temporal windows in the entorhinal-hippocampal loop[J].Neurobiol Learn Mem, 2015, 120: 69-83.
[25]SANDLER R A, SONG D, HAMPSON R E, et al.Hippocampal closed-loop modeling and implications for seizure stimulation design[J].J Neural Eng, 2015, 12(5): 056017.
[26]KINOSHITA M, IKEDA A, MATSUHASHI M, et al.Electric cortical stimulation suppresses epileptic and background activities in neocortical epilepsy and mesial temporal lobe epilepsy[J].Clin Neurophysiol, 2005, 116(6): 1291-1299.
[27]MELDRUM B S, AKBAR M T, CHAPMAN A G.Glutamate receptors and transporters in genetic and acquired models of epilepsy[J].Epilepsy Res, 1999, 36(2-3): 189-204.
[28]EID T, LEE T S, WANG Y, et al.Gene expression of glutamate metabolizing enzymes in the hippocampal formation in human temporal lobe epilepsy[J].Epilepsia, 2013, 54(2): 228-238.
(本文編輯:趙翠翠)
R742.1
C
10.3969/j.issn.2095-9400.2016.12.019
2016-04-05
浙江省教育廳訪問學(xué)者教師專業(yè)發(fā)展項目。
鄭茜茜(1978-),女,浙江溫州人,講師,碩士。
猜你喜歡
中國民間療法(2021年5期)2021-06-09 09:21:04
中華養(yǎng)生保健(2020年2期)2020-11-16 00:49:00
解放軍醫(yī)學(xué)院學(xué)報(2020年12期)2020-03-29 05:11:46
中成藥(2017年6期)2017-06-13 07:30:35
飲食科學(xué)(2017年5期)2017-05-20 17:11:53
臨床醫(yī)藥文獻(xiàn)雜志(電子版)(2017年11期)2017-05-17 04:48:10
安徽醫(yī)科大學(xué)學(xué)報(2015年9期)2015-12-16 11:09:44
中國當(dāng)代醫(yī)藥(2015年7期)2015-03-01 02:01:13
西南軍醫(yī)(2015年4期)2015-01-23 01:19:30
西部中醫(yī)藥(2014年6期)2014-03-11 16:07:47
