日本晴/R1126水稻重組自交系群體粒形性狀QTL定位
2016-03-18 07:58:57葉乃忠肖應輝
華北農學報 2016年1期
關鍵詞:水稻
葉乃忠,曾 蓋,唐 偉,肖應輝
(1.湖南農業大學 農學院,湖南 長沙 410128;2.南方糧油作物協同創新中心,湖南 長沙 410128)
?
日本晴/R1126水稻重組自交系群體粒形性狀QTL定位
葉乃忠1,2,曾蓋1,2,唐偉1,2,肖應輝1,2
(1.湖南農業大學 農學院,湖南 長沙410128;2.南方糧油作物協同創新中心,湖南 長沙410128)
摘要:為闡明大穗型秈稻品系R1126稻谷籽粒性狀的遺傳機制,以其與粳稻日本晴構建的重組自交系F10為材料,通過連續2年田間種植并測定各株系的籽粒粒長、粒寬和粒厚等表型性狀,結合利用SSR和SFP等分子標記構建的遺傳圖譜,對控制該群體的稻谷粒形性狀進行了QTL分析。2年試驗共檢測到10個控制粒長、粒寬和粒厚性狀的QTL,其中qGL3-1、qGL3-2和qGL9這3個粒長QTL,以及qGW2和qGW5這2個粒寬QTL在2年試驗中能被重復檢測;而粒厚性狀在2年試驗中檢測到5個QTL,但均只在1年試驗中出現。根據連鎖的分子標記信息,qGL3-2、qGW2和qGW5可能分別與已報道的主要粒形基因GS3、GW2和GW5等位;而qGL3-1和qGL9可能為新的粒長QTL,且在2年試驗中具有很好的重演性和穩定性,兩者的加性效應均能使粒長增加0.2 mm以上,對于改善稻谷外觀品質性狀具有較好的潛在應用價值。
關鍵詞:水稻;粒長;粒寬;粒厚;QTL定位
水稻粒形包括粒長、粒寬、長寬比以及粒厚等性狀,是稻谷(米)外觀品質性狀的重要表現形式[1],同時也是決定粒重進而影響水稻產量的重要因素[2]。然而,稻谷籽粒體積特別是粒寬和粒厚增大,往往伴隨著稻米外觀品質和加工品質變劣,進而影響稻米的商用和市場價值[3]。另一方面,父母本種子粒形差異可通過機械加工分離,從而使得水稻混播雜交制種成為可能,由于該技術可以大幅降低種子生產成本而備受種子企業關注[4]。因此,剖析水稻籽粒粒形性狀的遺傳調控機制,是實現水稻籽粒(種子)形態精細調控并通過分子設計塑造不同粒形水稻品種的前提。
鑒于粒形性狀在水稻產量和品質形成中的重要作用,粒長、粒寬、粒厚和粒重等粒形相關性狀的遺傳機理一直是研究的重點。近20年來,科學家們利用不同類型的遺傳群體,累計報道了400多個與水稻籽粒大小和形態有關的QTL[5],根據連鎖標記信息歸類整合發現至少有286個與粒形相關的QTL。其中,與粒長性狀相關的QTL 75個,與粒寬相關的QTL 66個,另有92個QTL控制水稻籽粒粒重,這些QTL在12條染色體上均有分布。迄今已有8個與籽粒大小有關的QTL被分離克隆,分別是GW2[6]、GS3[7]、GW5[8]、GS5[9]、GW8[10]、qGL3(qGL3.1)[11]、GIF1[12]、TGW6[13];此外,另有gw3.1[14]、gw8.1[15]、gw9.1[16]、qGL3-3a[17]、GW3[18]、GW6[19]、qGL4b[20]、qGL7[21]、GS7[22]、qGRLl[23]、qSS7[24]、GW1-1[24]、GW1-2[24]、Lk-4(t)[25]、qTGW3.2[26]、GS2[27]16個粒形相關基因/QTL被精細定位。這些與粒形性狀相關基因的分離與克隆為水稻外觀品質和產量的遺傳改良提供了理論基礎。
目前,針對這些粒形相關基因,育種家就其在水稻育種應用方面展開了進一步探索。張曉軍[28]發現,以水稻品種9311為受體的近等基因系9311NIL-qgl3的株高、分蘗數、穂粒數、穗長等性狀與受體親本保持一致,而籽粒粒長和粒重分別比受體親本增加約20%和37%,小區產量增產16.2%。楊梯豐等[29]以華粳秈74為遺傳背景且攜帶谷粒長基因GS3的單片段代換系與攜帶其他優良基因的單片段代換系雜交,分子聚合育成了谷粒長為 9.51~9.67 mm 的長粒形聚合品系,有效地改良了華粳秈 74的外觀品質。Wang等[30]通過分子標記輔助選擇聚合稻谷粒形基因qgw8與qgs3,創制的新品系 Huabiao1與受體親本HJX74產量相當,而稻谷粒形性狀得以明顯改良。這些粒形基因的成功應用,為粒形基因在水稻育種實踐上提供了成功先例。
R1126是國家雜交水稻工程技術研究中心選育的大穗型水稻品系,其平均穗粒數一般在400粒以上。筆者利用該大穗品系與粳稻品系日本晴構建的重組自交系群體開展了粒形性狀的遺傳模型分析[31],本研究擬進一步對粒長、粒寬、粒厚和粒重等粒形相關性狀進行QTL定位研究,旨在為大穗型水稻品系R1126在水稻粒形性狀改良中提供遺傳理論基礎。
1材料和方法
1.1試驗材料
以大穗秈稻品系R1126與粳稻品系日本晴(Nipponbare,NPB)為親本,在雜種F1自交分離獲得F2的基礎上通過單粒傳法構建包含185個株系的日本晴/R1126重組自交系群體(F10)。所有試材種子均由湖南農業大學水稻科學研究所提供。
1.2材料種植及表型鑒定方法
田間試驗分別于2012,2013年在湖南農業大學耘園試驗基地進行。2年均于當年5月25日播種所有試材,秧齡25 d左右移栽。每個株系種植1行,每行10株,株行距為20 cm×20 cm。按照當地水稻栽培技術要求進行田間管理與病蟲害防治,保證水稻在整個生長過程中無病蟲、鳥及鼠的危害。
于各株系成熟期,選取每株系除邊行外的中間8個單株收種,每一單株只收取最大穗,自然曬干后貯存用于性狀調查。每個株系隨機選取10粒飽滿種子,用電子游標卡尺(精確到0.01 mm)分別測定籽粒粒長、粒寬和粒厚,取10粒平均值作為該株系相應性狀的表型值。
1.3基因型分析方法
選取在親本R1126和日本晴間呈多態性且在12條染色體上基本均勻分布的122個SSR和SFP標記,用于基因型分析。SSR和SFP分析方法參照Chen等[32]的方法,10 μL反應體系包括:10 mmol/L Tris-HCl pH值8.3,50 mmol/L KCl,1.5 mmol/L MgCl2,50 μmol/L dNTPs,0.2 μmol/L引物,0.5 UTaqpolymerase和20 ng DNA 模板。擴增反應在ABI PCR system 2700上進行:94 ℃預變性5 min;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 1 min,共35個循環;最后,72 ℃延伸7 min。擴增產物用8%的非變性PAGE膠分離,銀染顯色。
1.4數據分析方法
采用MapMaker3.0軟件[33]計算分子標記間的遺傳距離,并繪制分子標記遺傳連鎖圖譜。使用WinQTL Cartographer 2.5軟件[34]用復合區間作圖法(CIM)進行QTL分析,以LOD值2.5作為檢測QTL的閾值,某一標記區間QTL的LOD值高于臨界閾值則認為存在一個QTL,同時估算QTL效應及其對性狀的貢獻率。
2結果與分析
2.1親本與群體性狀的表型分析
2012,2013年的試驗數據均表明,對于粒長、粒寬和粒厚性狀,在親本R1126和NPB間均表現出顯著性差異(表1)。粒長和粒厚性狀值均是親本R1126大于NPB,而粒寬性狀值則是NBP大于R1126。 在重組自交系群體中,粒長平均值介于雙親性狀值之間,而粒厚和粒寬的平均值低于雙親性狀值。重組自交系群體各粒形性狀的變異幅度較大,在2012,2013年不同年份多出現了超親現象。
在NPB/R1126重組自交系群體中,3個性狀的分布特征均表現為連續變異(圖1),分布的偏度和峰度系數的絕對值均小于1,基本上符合正態分布,暗示在該群體中所有粒形性狀均表現為多基因控制的數量性狀遺傳模式。同一性狀2年間的頻率分布特征基本一致,但2012年各性狀值多表現為大于2013年,可能因不同年份氣候不同導致水稻籽粒灌漿差異。
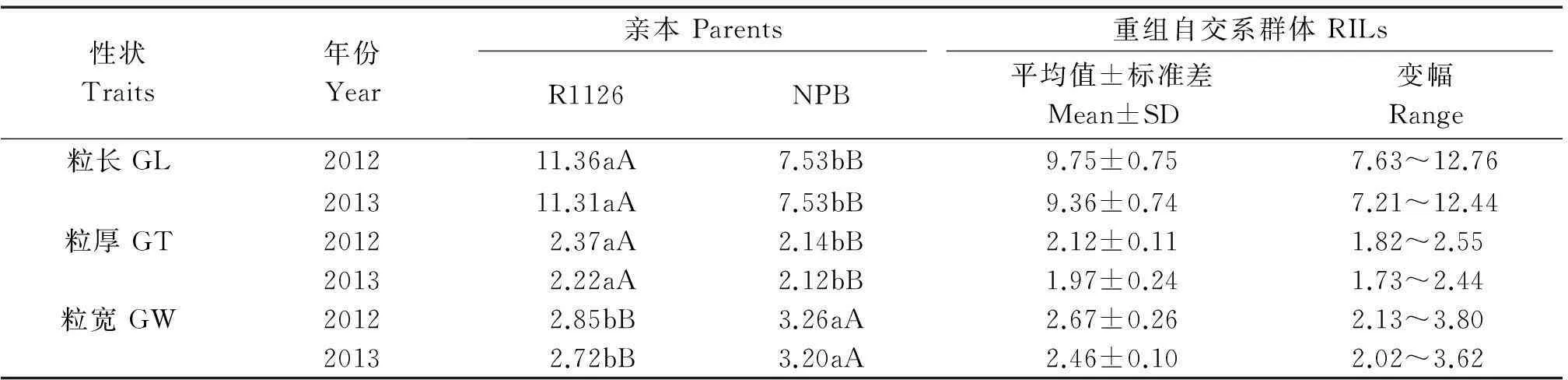
表1 水稻重組自交系及親本2年各粒形性狀表型
注:數字后的大、小寫字母分別代表在1%和5%水平上差異顯著。
Note:The capital and lowercase indicated significant difference at level ofP=0.01 andP=0.05,respectively.

圖1 日本晴/R1126重組自交系群體粒形相關性狀分布特征
2.2粒形性狀QTL定位
采用SSR和SFP標記構建了日本晴/R1126重組自交系群體遺傳連鎖圖譜,該圖譜包含覆蓋水稻全基因組12條染色體的122個標記,連鎖圖譜全長1 205.6 cM,標記間平均距離約為9.9 cM。該圖譜共包含14個連鎖群,其中第3,4染色體各包含2個連鎖群。依據該遺傳連鎖圖譜,結合稻谷粒形表型數據,對該群體粒形性狀進行了QTL分析。
2.2.1粒長性狀QTL檢測2年試驗重復檢測到3個稻谷粒長相關的QTL(表2、圖2),其中第3染色體上的2個粒長QTL,qGL3-1和qGL3-2,分別位于RM5474-RM4992和RM6080-RM5626標記區間;第9染色體的qGL9位于短臂端SFP9_2附近。粒長相關QTL的遺傳效應在2年試驗中基本一致,其LOD值為2.73~5.01,貢獻率為5.41%~11.76%。3個粒長QTL的加性效應均表現為正效應,其增效等位基因均來自R1126,可增加稻谷粒長0.20~0.31 mm。在3個粒長QTL中,位于第3 染色體RM6080-RM5626區間的qGL3-2貢獻率最大,能解釋表型變異的10.56%~11.76%。
2.2.2粒寬性狀QTL檢測粒寬性狀的QTL分析表明,位于第2染色體RM7082-RM6853區間的qGW2和第5染色體RM5874-RM405區間的qGW5,在2年試驗中均能重復檢測(表2、圖2)。qGW5在2年試驗中的LOD值分別為9.21和8.56,貢獻率分別達17.93%和17.44%;而qGW2的貢獻率相對較低,為8.26%和8.65%。此外,2個與粒寬性狀相關QTL的加性效應均為負加性效應,來自R1126的等位基因分別能使粒寬減小0.08~0.09,0.12~0.13 mm。

表2 粒形相關性狀QTL
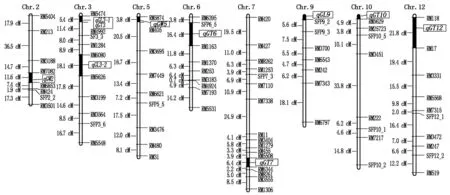
圖2 水稻粒形相關性狀QTL在染色體上的分布
3討論
本研究采用粳稻日本晴和大穗型秈稻品系R1126構建的重組自交系群體對該群體粒形性狀進行了QTL分析,2年的田間重復試驗共檢測到10個粒形相關QTL。其中,3個粒長QTL和2個粒寬QTL均在2年試驗中能重復檢測到,表明粒長和粒寬的遺傳機制相對簡單,受環境因素影響相對較小。在2012,2013年試驗中分別檢測到2,3個粒厚QTL,這些QTL都只能在其中1年被檢測到,暗示粒厚性狀受環境因素影響較大,其遺傳相對復雜。這一現象與水稻籽粒的發育特征是吻合的,粒長和粒寬主要由穎殼長度和寬度決定,而粒厚則主要與籽粒充實度有關,不同環境條件下籽粒灌漿程度差異會造成籽粒厚度變化。
比較已報道的水稻粒形性狀基因/QTL[5-27],發現本研究檢測到的粒長QTLqGL3-2所在染色體區段RM6080-RM5626與Fan等[7]檢測的GS3位置相近;粒寬qGW2所在的RM7082-RM6853和qGW5所在的RM5874-RM405分別與粒寬基因GW2、GW5所在染色體區段一致,推斷這3個QTL可能與已報道的粒長、粒寬基因等位。本研究檢測的2個粒長QTL,即位于第3染色體RM5474-RM4992區間的qGL3-1和第9染色體短臂端SFP9_2附近的qGL9,在2年試驗中具有很好的重演性和穩定性,兩者的加性效應均能使粒長增加0.2 mm以上,對于改善稻谷外觀品質性狀具有較好的潛在應用價值,值得進一步深入研究。
本研究中重組自交系群體親本之一R1126是一個平均穗粒數400粒以上的大穗型水稻品系,已用于配制雜交稻組合或改良其他水稻品系[35]。本研究發掘的3個粒長QTL使粒長增加的等位基因均來自于R1126,而該品系中的2個粒寬等位QTL卻使粒寬變小。通過合理利用R1126的稻谷粒形基因/QTL,可以增大稻谷的長寬比,改善稻谷(稻米)的外觀品質性狀。因此,本研究所獲得的與稻谷粒長、粒寬QTL連鎖的分子標記,可望為R1126應用于稻谷粒形性狀改良提供技術支持。
參考文獻:
[1]徐正進,陳溫福,馬殿榮,等.稻谷粒形與稻米主要品質性狀的關系[J].作物學報,2004,30(9):894-900.
[2]石春海,申宗坦.秈稻粒形及產量性狀的加性相關和顯性相關分析[J].作物學報,1996,22(1):36-42.
[3]林荔輝,吳為人.水稻粒型和粒重的QTL定位分析[J].分子植物育種,2003,1(3):337-342.
[4]湯國華,謝紅軍,余應弘.雜交水稻機械化制種研究的現狀、問題與對策[J].湖南農業科學,2012(3):133-136.
[5]Database G./[Z],2015.http://www.gramene.org/.
[6]Song X J,Huang W,Shi M,et al.A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase[J].Nature Genetics,2007,39(5):623-630.
[7]Fan C,Xing Y,Mao H,et al.GS3,a major QTL for grain length and weight and minor QTL for grain width and thickness in rice,encodes a putative transmembrane protein[J].Theoretical and Applied Genetics,2006,112(6):1164-1171.
[8]Weng J E,Gu S H,Wan X Y,et al,Isolation and initial characterization of GW5,A major QTL associated with rice grain width and weight[J].Cell Research,2008,18(12):1199-1209.
[9]Li Y,Fan C,Xing Y,et al.Natural variation in GS5 plays an important role in regulating grain size and yield in rice[J].Nature Genetics,2011,43(12):1266-1269.
[10]Wang S,Wu K,Sun X,et al.The regulation of OsSPL16 promotes grain yield in rice[J].Nature Genetics,2012,44(8):950-954.
[11]Hu Z,He H,Zhang S,et al.A kelch motif-containing serine/threonine protein phosphatase determines the large grain QTL trait in rice[J].Journal of Integrative Plant Biology,2012,54(12):979-990.
[12]Wang E,Wang J,Zhu X,et al.Control of rice grain-filling and yield by a gene with a potential signature of domestication[J].Nature Genetics,2008,40(11):1370-1374.
[13]Ishimaru K,Hirotsu N,Madoka Y,et al.Loss of function of the IAA-glucose hydrolase geneTGW6 enhances rice grain weight and increases yield[J].Nature Genetics,2013,45(6):707-711.
[14]Li J,Thomson M,Mccouch S R.Fine mapping of a grain-weight quantitative trait locus in the pericentromeric region of rice chromosome 3[J].Genetics,2004,168(4):2187-2195.
[15]Xie X,Song M H,Jin F,et al.Fine mapping of a grain weight quantitative trait locus on rice chromosome 8 using near-isogenic lines derived from a cross betweenOryzasativaandOryzarufipogon[J].Theoretical and Applied Genetics,2006,113(5):885-894.
[16]Jin F X,Ji S D,Xie X B,et al.Detection of epistatic interaction of two QTLs,gw8.1 and gw9.1,underlying grain weight using nearly isogenic lines in rice[J].Breeding Science,2011,61(1):69-75.
[17]Wan X Y,Wan J M,Jiang L,et al.QTL analysis for rice grain length and fine mapping of an identified QTL with stable and major effects[J].Theoretical and Applied Genetics,2006,112(7):1258-1270.
[18]Guo L,Ma L,Jiang H,et al.Genetic analysis and fine mapping of two genes for grain shape and weight in rice[J].Journal of Integrative Plant Biology,2009,51(1):45-51.
[19]Kato T,Segami S,Toriyama M,et al.Detection of QTLs for grain length from large grain rice (OryzasativaL.)[J].Breeding Science,2011,61(3):269-274.
[20]Bai X,Luo L,Yan W,et al.Genetic dissection of rice grain shape using a recombinant inbred line population derived from two contrasting parents and fine mapping a pleiotropic quantitative trait locus qGL7[J].BMC Genetics,2010,11(1):16.
[21]Shao G,Wei X,Chen M,et al.Allelic variation for a candidate gene for GS7,responsible for grain shape in rice[J].Theoretical and Applied Genetics,2012,125(6):1303-1312.
[22]Singh R,Singh A K,Sharma T R,et al.Fine mapping of grain length QTLs on chromosomes 1 and 7 in Basmati rice (OryzasativaL.)[J].Journal of Plant Biochemistry and Biotechnology,2012,21(2):157-166.
[23]Qiu X,Gong R,Tan Y,et al.Mapping and characterization of the major quantitative trait locus qSS7 associated with increased length and decreased width of rice seeds[J].Theoretical and Applied Genetics,2012,125(8):1717-1726.
[24]Yu S W.Fine mapping of quantitative trait loci for grain length and grain width on the short arm of rice chromosome 1[J].Chinese Journal of Rice Science,2008,22(5):465-471.
[25]Zhou L Q,Wang Y P,Li S G.Genetic analysis and physical mapping ofLk-4(t),a major gene controlling grain length in rice,with a BC2F2 population[J].Acta genetica Sinica,2006,33(1):72-79.
[26]Tang S Q,Shao G N,Wei X J,et al.QTL mapping of grain weight in rice and the validation of the QTL qTGW3.2[J].Gene,2013,527(1):201-206.
[27]Zhang W H,Sun P Y,He Q,et al.Fine mapping ofGS2,a dominant gene for big grain rice[J].The Crop Journal,2013,1(2):160-165.
[28]張曉軍.水稻粒長基因qGL3的定位克隆,功能分析及育種利用研究[D].南京:南京農業大學,2012.
[29]楊梯豐,曾瑞珍,朱海濤,等.水稻粒長基因GS3在聚合育種中的效應[J].分子植物育種,2010,8(1):59-66.
[30]Wang S,Wu K,Yuan Q,et al.Control of grain size,shape and quality by OsSPL16 in rice[J].Nature Genetics,2012,44(8):950-954.
[31]葉乃忠,曾蓋,唐偉,等.2套半同胞秈粳交重組自交系粒形性狀的相關和遺傳分析[J].作物研究,2015(2):111-116.
[32]Chen X,Temnykh S,Xu Y,et al.Development of a microsatellite framework map providing genome-wide coverage in rice (OryzasativaL.)[J].Theoretical and Applied Genetics,1997,95(4):553-567.
[33]Lincoln S,Daley M,Lander E.Constructing genetic maps with MAPMAKER/EXP 3.0[M].3rd edn.Cambridge:Whitehead Institute Technical Report,1992.
[34]Zeng Z B.Precision mapping of quantitative trait loci[J].Genetics,1994,136:1457-1468.
[35]周紅英,張桂蓮,肖應輝,等.超大穗型水稻R1126及其組合穗頸輸導組織與籽粒灌漿結實的關系[J].中國水稻科學,2014,28(4):411-418.
Mapping of QTL Associated with Grain Shape Traits in Nipponbare/R1126 Recombinant Inbred Lines Population of Rice
YE Naizhong1,2,ZENG Gai1,2,TANG Wei1,2,XIAO Yinghui1,2
(1.College of Agronomy,Hunan Agricultural University,Changsha410128,China;2.Southern Regional Collaborative Innovation Center for Grain and Oil Crops in China,Changsha410128,China)
Abstract:In order to clarify the genetic mechanism of grain shape traits for large panicle indica rice line,R1126,a set of recombinant inbred lines,which contain 185 lines,was developed by using R1126 crossed to the japonica rice line,Nipponbare.By measuring the grain shape traits include grain length (GL),grain width (GW) and grain thickness (GT) in two years field experiment,combining with the genetic map constructed with SSR and SFP molecular markers,QTL associated with grain shape traits were analyzed.Total ten QTL associated with GL,GW and GT,were detected in two years experiment.Among them,three QTL related to GL including qGL3-1,qGL3-2 and qGL9,and two QTL related to GW including qGW5 and qGW2,were be detected repeatedly in two years,respectively.However,no one QTL related to GT could be detected repeatedly.By comparing the molecular marker information,qGL3-2,qGW2 and qGW5 might be allele with the reported genes of GS3,GW2 and GW5,respectively.The two QTL associated with grain length,qGL3-1 and qGL9,which were expressed stably and repeated in two years experiment,exhibiting the additive effect of increasing the grain length of 0.2 mm,could be used for improving rice appearance quality.
Key words:Rice;Grain length;Grain width;Grain thickness;QTL mapping
doi:10.7668/hbnxb.2016.01.015
中圖分類號:S511.03
文獻標識碼:A
文章編號:1000-7091(2016)01-0090-06
作者簡介:葉乃忠(1989-),男,福建武夷山人,在讀碩士,主要從事水稻遺傳育種研究。通訊作者:肖應輝(1970-),男,湖南漣源人,研究員,博士,主要從事水稻遺傳育種研究。
基金項目:教育部“創新團隊發展計劃”項目(IRT1239);湖南省高校科技創新團隊項目
收稿日期:2015-12-02
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
