不同密度雜草稻脅迫對栽培稻光合作用及生理特性的影響
2016-03-18 07:59:04王曉琳張曉昉張卓亞
華北農學報 2016年1期
王曉琳,張曉昉,李 可,張卓亞,李 貴
(1.江蘇省農業科學院 植物保護研究所,江蘇 南京 210014;2.南京農業大學 生命科學學院,江蘇 南京 210095)
?
不同密度雜草稻脅迫對栽培稻光合作用及生理特性的影響
王曉琳1,張曉昉2,李可2,張卓亞2,李貴1
(1.江蘇省農業科學院 植物保護研究所,江蘇 南京210014;2.南京農業大學 生命科學學院,江蘇 南京210095)
摘要:為了探明雜草稻脅迫對栽培稻產量的影響及其生理原因,以雜草稻JS-Y1和栽培稻南粳44為材料,通過田間試驗研究不同密度雜草稻(0,4,8,12,16株/m2)脅迫對栽培稻拔節期、抽穗期和灌漿中期光合性能、葉綠素熒光參數、透光率、膜脂過氧化和保護酶活性及產量的影響。結果表明:與對照相比,隨著雜草稻發生密度的增加,雜草稻脅迫不同程度抑制了栽培稻不同生育期生理指標,影響栽培稻正常的生理功能,雜草稻的存在對栽培稻和雜草稻的共生群體的透光率造成了影響,在距地面20,50,80 cm和穗下10 cm,較對照分別降低54.03%~72.49%,39.68%~61.57%,48.98%~54.40%,38.07%~56.76%,達顯著水平,葉片葉綠素含量、光合參數、PSⅡ的實際光化學效率(ФPSⅡ)和光化學淬滅系數(qP),以及膜脂過氧化酶活性,均隨著栽培稻生育進程和雜草稻密度的增加而呈逐漸降低的趨勢,在灌漿中期尤為明顯;與對照(雜草稻發生密度為0)相比,雜草稻發生密度為4,8,12,16株/m2時,栽培稻產量分別下降了16.73%,43.01%,66.61%,83.24%,達顯著差異。說明不同密度雜草稻脅迫時,影響了栽培稻群體的透光率,造成地上光合能力下降,栽培稻葉片細胞膜系統受到破壞是造成栽培稻產量降低的重要生理原因。
關鍵詞:雜草稻;栽培稻;光合作用;生理特性
稻田雜草不僅與水稻競爭自然資源,還是許多病蟲害的中間寄主,一直是影響水稻高產穩產的限制因素。近年來,隨著輕簡化栽培技術的發展,水稻田雜草群落發生很大變化,其中雜草稻(Oryzasativaf.spontanea)的不斷蔓延嚴重影響水稻產量及稻米品質,正成為輕簡化稻作技術的關鍵問題[1-2]。雖然在水稻田雜草防治方面積累了豐富的經驗和技術,形成了以化學除草為核心的雜草防除技術體系,但是由于雜草稻與水稻同為稻屬植物,水稻生長過程中還缺少有效的除草劑可以防除雜草稻[3-4]。目前,國內外有關雜草稻的研究主要集中在雜草稻生物學生態學特性、對栽培稻生長發育以及生態安全的影響、起源與分類、遺傳多樣性、抗逆特性及其利用等方面[5-9],其中與栽培稻競爭方面的研究主要集中在雜草稻對栽培稻生長發育、根系形態、產量以及產量構成因素結實率等影響,當雜草稻密度為1~9 株/m2,栽培稻實際產量減少5.62%~33.81%[10];另外馬殿榮等[11]通過研究發現,雜草稻對栽培稻群體生長環境影響顯著,當栽培田中雜草稻密度超過3株/m2時,移栽稻群體照度和透光率降低,日平均氣溫和日最高氣溫極顯著降低等,最終雜草稻密度增大導致稻田群體環境惡化。
以往有關雜草稻對栽培稻的研究多集中在雜草稻密度對栽培稻表型特征及產量的影響,對于栽培稻的防御機制以及造成栽培稻產量降低的生理原因缺乏深入系統的研究。為此,本研究從光合特性、葉綠素熒光特性、抗氧化酶系統等方面觀察不同密度雜草稻脅迫對栽培稻生理特性的影響,為防治雜草稻的危害提供一定的理論依據。
1材料和方法
1.1供試材料
水稻:南粳44(江蘇省農業科學院糧食作物研究所提供);雜草稻:JS-Y1,秈型(采集于江蘇省揚中市西來橋鎮直播稻田)。
1.2試驗設計
2014年6月21日將水稻種子和雜草稻種子催芽后播種于江蘇省農業科學院植物保護研究所實驗場內15 m2的水泥池中,水稻播種量為67.5 kg/hm2;1 m2留雜草稻0(對照CK),4(T1),8(T2),12(T3),16(T4) 株/m2,拔出小區內其他雜草。各處理重復4次,隨機區組排列,常規防治病蟲。
1.3測定項目與方法
1.3.1光合參數的測定分別于水稻拔節期、抽穗期和灌漿中期使用LI-6400便攜式紅外氣體分析儀(美國基因公司)于晴朗天氣的上午9:00-11:30測定水稻最新全展葉凈光合速率,各處理重復測定6張葉片。
1.3.2水稻葉片葉綠素的測定分別于水稻拔節期、抽穗期和灌漿中期,選擇長勢均勻的水稻植株,每小區隨機取10株,用SPAD-502便攜式葉綠素計(日本美能達公司)測定水稻最新全展葉,每處理重復4次。
1.3.3葉綠素熒光參數的測定分別于水稻拔節期、抽穗期和灌漿中期,選擇長勢均勻的水稻植株,于暗室處理30 min后使用FMS-2便攜式脈沖調制式熒光儀(英國漢莎科學儀器公司生產)測定水稻葉片葉綠素熒光參數。
1.3.4透光率的測定群體及冠層照度:于抽穗期,選擇AccuPAR植物冠層分析儀進行田間測定,測定高度為從地面向上20,50,80 cm 處及穗下10 cm處,4次重復。透光率=稻株不同位置的光照強度/稻株上方光照強度×100%。
1.3.5丙二醛、過氧化氫酶和過氧化物酶的測定[12]分別于水稻拔節期、抽穗期和灌漿中期測定水稻中丙二醛(Malondialdehyde,MDA)含量、用紫外吸收法測定出過氧化氫酶(Catalase,CAT)的活性,用愈創木酚法測定過氧化物酶(Peroxidase,POD)活性。
1.3.6考種、計產收獲前去除雜草稻。于成熟期每個小區取10穴水稻風干后進行考種,主要調查穗粒數、結實率和千粒質量等指標。
1.4數據分析與方法
采用Excel和SPSS軟件進行數據分析處理和作圖。
2結果與分析
2.1雜草稻脅迫對栽培稻光合參數的影響
由表1可見,水稻拔節期時,隨著雜草稻株數的增加對栽培稻各光合參數影響不大,對照與處理無顯著差異;抽穗期時,水稻凈光合速率、蒸騰速率和水分利用率隨著雜草稻株數的增加而呈降低趨勢,其中1 m2雜草稻株數為4~16時,凈光合速率顯著降低8.71%~23.67%,雜草稻株數為12~16株時,水稻蒸騰速率顯著降低9.08%~10.26%;灌漿中期時,水稻凈光合速率、氣孔導度和水分利用率隨著雜草稻株數的增加而呈降低趨勢,其中1 m2雜草稻株數為4~16株時,凈光合速率顯著降低4.83%~19.41%,1 m2雜草稻株數為8~16株時,水稻葉片氣孔導度顯著降低18.00%~22.00%。
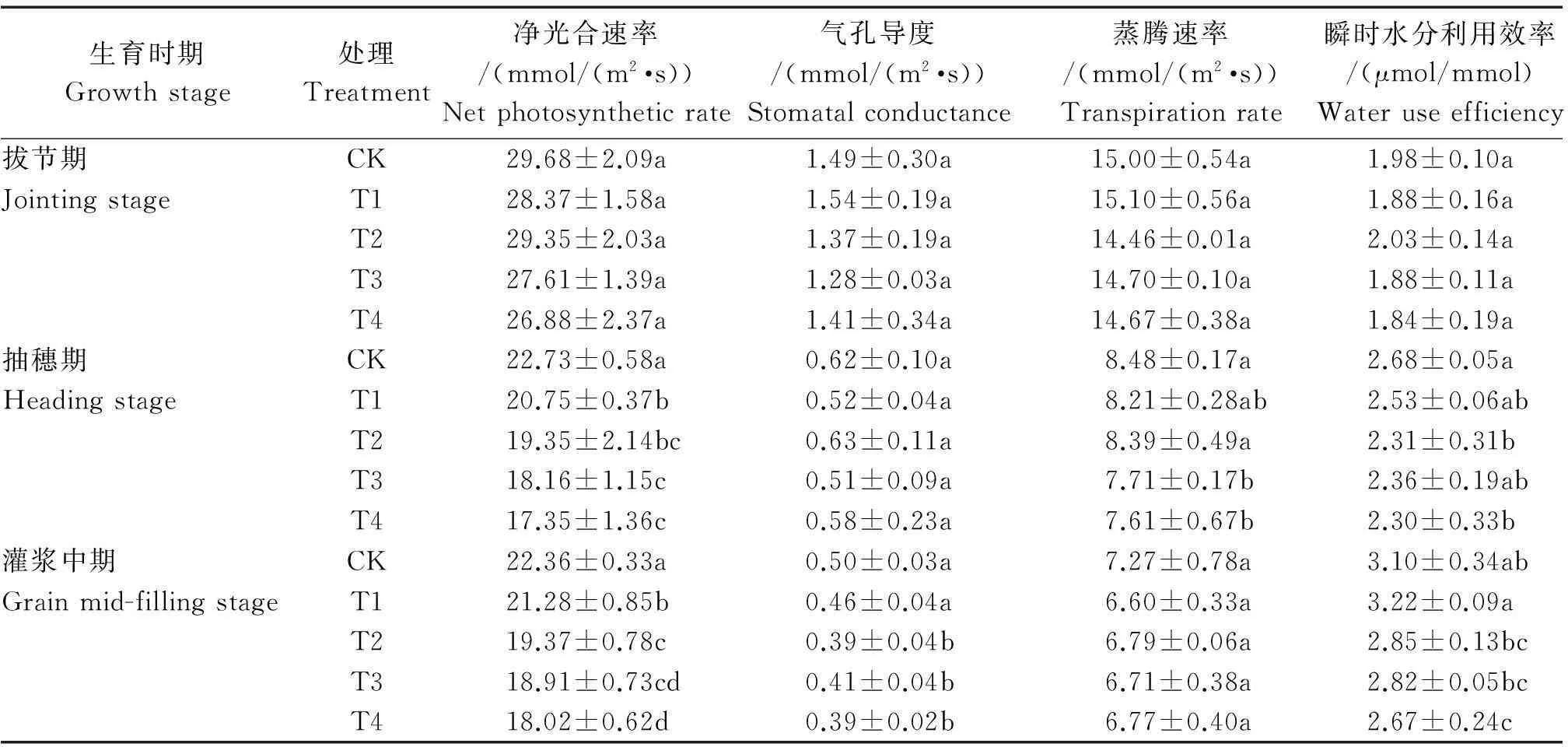
表1 雜草稻脅迫對各時期栽培稻劍葉光合參數的影響
注:不同小寫字母表示處理間差異顯著(P<0.05)。表2~3、圖1~4同。
Note:Different letters meant significant difference among treatments at 0.05 level.The same as Tab.2-3,Fig.1-4.
2.2雜草稻脅迫對栽培稻葉綠素及葉綠素熒光參數的影響
由表2可知,隨著雜草稻株數的增加對水稻拔節期、抽穗期和灌漿中期水稻葉片PSⅡ的最大光化學效率(Fv/Fm)和開放的PSⅡ反應中心的激發能捕獲效率(Fv′/Fm′)沒有明顯的影響;對水稻抽穗期和灌漿中期水稻葉片PSⅡ的實際光化學效率(ФPSⅡ)和光化學淬滅系數(qP)影響較大,當1 m2雜草稻株數為4~16株時,水稻葉片PSⅡ的實際光化學效率降低了13.27%~16.76%,光化學淬滅系數降低了9.78%~12.56%,達顯著水平;當1 m2雜草稻株數為16株時,水稻拔節期、抽穗期和灌漿中期水稻葉片葉綠素含量下降4.75%~14.07%。
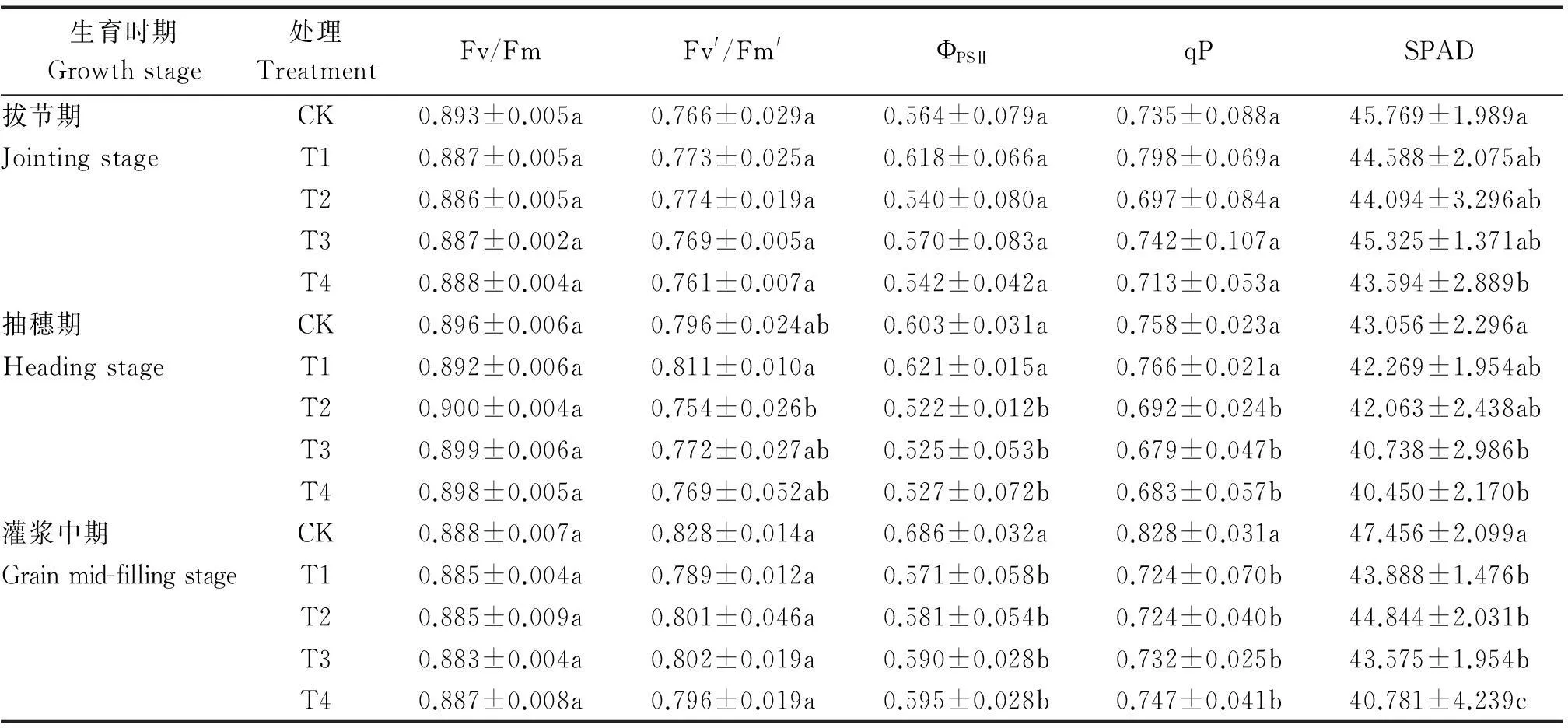
表2 雜草稻脅迫對各時期栽培稻葉綠素熒光參數的影響
2.3雜草稻脅迫對栽培稻透光率的影響
從圖1可以看出,隨著雜草稻株數的增加,在4個位置中栽培稻透光率均呈現逐漸降低的趨勢。在距地面20,50,80 cm,穗下10 cm,較對照分別降低54.03%~72.49%,39.68%~61.57%,48.98%~54.40%,38.07%~56.76%,達顯著水平。
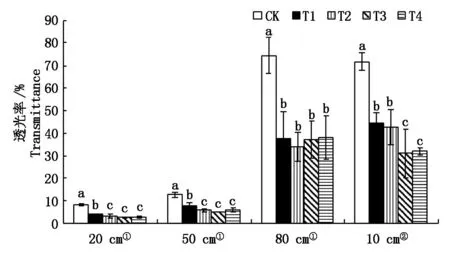
①.距地面的高度;②.距穗下的高度。
2.4雜草稻脅迫對栽培稻MDA含量、CAT活性、POD活性的影響
由圖2~4可見,隨著雜草稻株數的增加對水稻拔節期、抽穗期和灌漿中期MDA含量(以鮮質量計)影響較大,當1 m2雜草稻株數為8~16株時,各處理MDA含量較對照上升30.37%~82.63%,達顯著水平;在拔節期,栽培稻CAT活性和POD活性(以鮮質量計)隨著雜草稻株數的增加呈現先上升后下降的趨勢;在抽穗期水稻POD活性隨著雜草稻株數的增加呈現先上升后下降的趨勢,水稻CAT活性隨著雜草稻株數的增加呈現下降趨勢;在灌漿中期,栽培稻CAT活性和POD活性隨雜草稻株數的增加呈下降趨勢,POD活性較對照顯著下降22.22%~48.15%,CAT活性較對照顯著下降31.03%~56.32%。
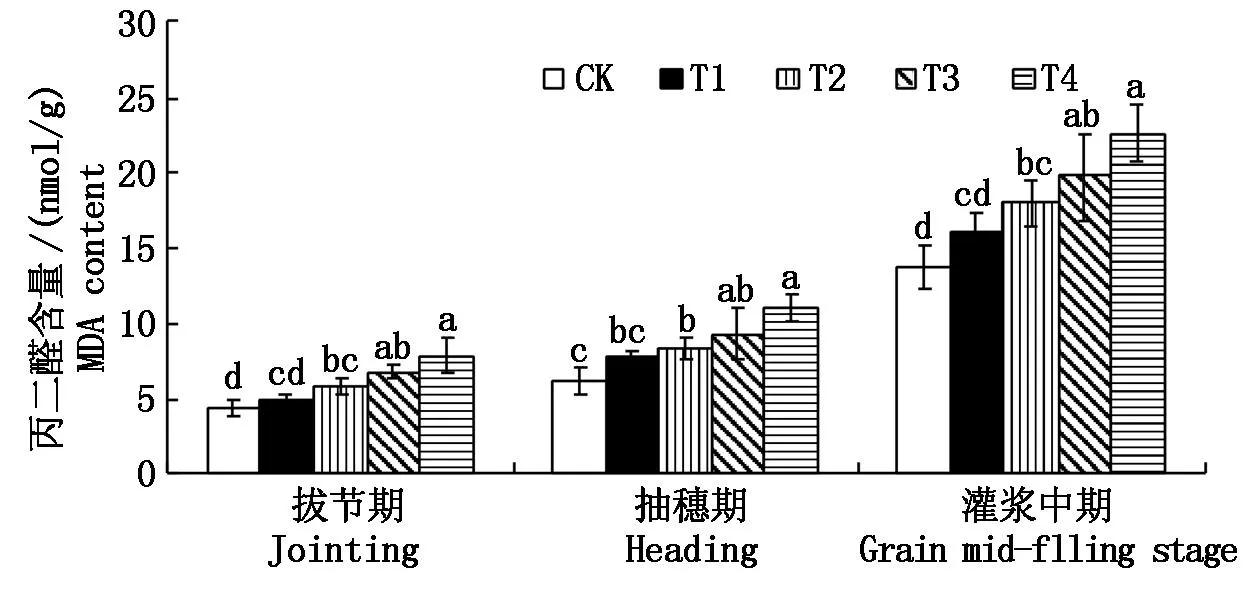
圖2 雜草稻脅迫對栽培稻MDA含量的影響

圖3 雜草稻脅迫對栽培稻POD活性的影響
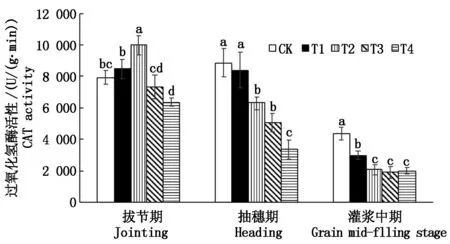
圖4 雜草稻脅迫對栽培稻CAT活性的影響
2.5雜草稻脅迫對栽培稻產量及其構成因素的影響
從表3可以看出,稻田中雜草稻密度為4株/m2,對栽培稻產量影響顯著,隨著雜草稻密度的增大,減產幅度加大。雜草稻密度為4,8,12,16株/m2處理分別比對照減產16.73%,43.01%,66.61%,83.24%。隨著雜草稻密度的上升,栽培稻的千粒質量、穗粒數、有效穗數和結實率均不同程度的降低,當雜草稻密度達到12株/m2株時,下降程度均達顯著水平。由此可得出,雜草稻對栽培稻產量影響顯著,栽培稻產量顯著降低,且隨著雜草稻密度的增加,降低程度也隨之大幅增加。

表3 雜草稻脅迫對栽培稻產量及其構成因素的影響
3討論
雜草稻與栽培稻之間存在對光照、土壤養分、水分和空間等資源的競爭,且這種對資源的競爭隨著雜草稻密度的增加而加劇。有研究表明,雜草稻與栽培稻競爭中所處的優勢地位及對栽培稻生長和產量的影響不同與雜草稻不同種群類型、混種密度和遺傳多樣性相關[13];吳云艷等[14]通過盆栽替代法研究雜草稻競爭對栽培稻根系發育的影響,結果表明,隨著栽培稻生育進程的推進和雜草稻密度的增加,導致栽培稻根系形態指標和生理機能受到抑制,進而引起栽培稻產量下降。本研究表明,隨著雜草稻密度的增大,減產幅度加大,雜草稻密度為4,8,12,16株/m2處理分別比對照減產16.73%,43.01%,66.61%,83.24%。
光照是水稻產量的限制因子,對作物生長及產量形成起著不可替代的作用[15]。當不同密度雜草稻存在時,必然影響水稻對光照的捕獲,影響共生群體的透光率。本試驗通過對抽穗期栽培稻群體透光率測定結果表明,隨著雜草稻株數的增加,在4個位置中栽培稻透光率均呈現逐漸降低的趨勢;透光率降低將導致光照強度減弱,引起光合能力下降,影響水稻生長發育。有研究表明,弱光使得水稻光合能力下降,干物質生產量下降,最終導致產量顯著下降[16-17]。本試驗通過對拔節期、抽穗期和灌漿中期光合參數的測定結果表明,隨著雜草稻株數的增加,抽穗期栽培稻凈光合速率、蒸騰速率和水分利用率降低;灌漿中期時,水稻凈光合速率、氣孔導度和水分利用效率隨著雜草稻株數的增加而降低;栽培稻拔節期時,隨著雜草稻株數的增加對栽培稻各光合參數影響不大;說明抽穗期后的栽培稻群體的透光率降低,導致栽培稻光合能力下降。葉綠素熒光動力學方法可以快速和靈敏的測定出光照對植物光合生理的影響,光照條件與植物熒光參數的受抑制程度密切相關。楊虎等[18]研究得出,遮陰條件下,水稻葉片吸收的光能達不到其固定CO2所需的能量,從而降低其所產生的激發能。這些激發能達不到自身體內細胞活動的基本需要,不能產生足夠的活性氧,從而降低了PSⅡ的反應中心和原初光化學反應。本研究表明,雜草稻脅迫導致栽培稻群體透光率下降,對水稻抽穗期和灌漿中期水稻葉片PSⅡ的實際光化學效率和光化學淬滅系數影響較大。
當植物遭受生物或非生物脅迫時,細胞內活性氧的動態平衡遭到破壞,活性氧含量上升和膜質過氧化加劇超過植物體內正常所能承受的閥值,會導致傷害的發生[19]。本研究結果表明,不同密度雜草稻脅迫下,在拔節期,栽培稻CAT活性和POD活性隨著雜草稻株數的增加呈現先上升后下降的趨勢,在灌漿中期時,栽培稻CAT活性和POD活性隨著雜草稻株數的增加而呈下降趨勢,表明栽培稻能夠通過提高保護酶活性、增強清除活性氧和自由基的能力而減輕逆境對自身的傷害,但隨著雜草稻脅迫時間的延長或者脅迫密度的增加,保護酶活性以及清除活性氧和自由基的能力減弱,從而導致質膜的嚴重損傷;栽培稻膜透性越來越大,使膜脂過氧化產物MDA含量增加。總之,不同密度雜草稻脅迫時,栽培稻群體的透光率下降,透光率的降低不僅使葉片細胞膜系統遭到破壞,還導致地上光合能力下降。有研究表明,光合能力的降低必將影響栽培稻的物質生產,最終造成減產[20-21],這可能也是雜草稻脅迫栽培稻減產的重要原因。
參考文獻:
[1]Tanzo I R,Martin E C,Chauhan B S.Weedy rice (OryzasativaL.) problem in rice (OryzasativaL.) based cropping systems in the Philippines [J].American Journal of Plant Sciences,2013,4:2359-2366.
[2]Azmi M,Azlan S,Yim K M,et al.Control of weedy rice in direct-seeded rice using the clearfield production system in Malaysia [J].Pakistan Journal of Weed Science Research,2012,18:49-53.
[3]Shen X E,Gao X H,Eneji A E,et al.Chemical control of weedy rice in precise hill-direct-seeded rice in South China[J].Weed Biology and Management,2013,13(1):39-43.
[4]Chauhan B S.Effect of weedy rice variants and weedy rice density on the growth and yield of two rice cultivars in the Philippines[J].Canadian Journal of Plant Protection,2013,1(1):28-34.
[5]Sales M A,Burgos N R,Shivrain V K,et al.Morphological and physiological responses of weedy red rice (OryzasativaL.) and cultivated rice (O.sativa) to N supply [J].American Journal of Plant Sciences,2011,2:569-577.
[6]Shivrain V K,Burgos N R,Agrama H A,et al.Genetic diversity of weedy red rice(Oryzasativa)in Arkansas[J].Weed Research,2010,50(4):289-302.
[7]Burgos N R,Shivrain V K,Scott R C,et al.Differential tolerance of weedy red rice (OryzasativaL.) from Arkansas,USA to glyphosate[J].Crop Protection,2011,30(8):986-994.
[8]Messeguer J,Fogher C,Guiderdoni E,et al.Field assessments of gene flow from transgenic to cultivated rice (OryzasativaL.) using a herbicide resistance gene as tracer marker[J].Theoretical and Applied Genetics,2001,103(8):1151-1159.
[9]Zhang Z,Dai W,Song X,et al.A model of the relationship between weedy rice seed-bank dynamics and rice-crop infestation and damage in Jiangsu Province,China[J].Pest Management Science,2014,70(5):716-724.
[10]宋冬明,馬殿榮,楊慶,等.雜草稻對栽培粳稻產量和品質及群體微生態環境的影響[J].作物學報,2009,35(5):914-920.
[11]馬殿榮,孔德秀,高齊,等.雜草稻對東北移栽稻群體生長環境及產量的影響[J].中國水稻科學,2014,28(2):211-216.
[12]王學奎.植物生理生化實驗原理和技術[M].2版.北京:高等教育出版社,2006:280-281.
[13]Dai L,Dai W,Song X,et al.A comparative study of competitiveness between different genotypes of weedy rice (Oryzasativa) and cultivated rice[J].Pest Management Science,2014,70(1):113-122.
[14]吳云艷,馬殿榮,李金英,等.雜草稻競爭對栽培稻根系形態生理特性及產量的影響[J].華北農學報,2010,25(5):150-154.
[15]許大全.光合作用效率[M].上海:上海科學技術出版社,2002:164-167.
[16]楊東,段留生,謝華安,等.花前光照虧缺對水稻物質積累及生理特性的影響[J].中國生態農業學報,2011,19(2):347-352.
[17]張自常,李永豐,楊霞,等.干濕交替灌溉條件下不同稗草對水稻光合特性和產量的影響[J].作物學報,2015,41(11):1748-1757.
[18]楊虎,戈長水,應武,等.遮蔭對水稻冠層葉片SPAD值及光合、形態特性參數的影響[J].植物營養與肥料學報,2014,20(3):580-587.
[19]Dong X,Bi H,Wu G,et al.Drought-induced chilling tolerance in cucumber involves membrane stabilisation improved by antioxidant system[J].International Journal of Plant Production,2013,7(1):67-79.
[20]陳乾,劉科,何愛斌,等.氮肥和密度互作對機插超級雜交稻葉型、光合特性及產量的影響[J].河南農業科學,2015,44(4):27-30.
[21]朱萍,楊世民,馬均,等.遮光對雜交水稻組合生育后期光合特性和產量的影響[J].作物學報,2008,34(11):2003-2009.
Effect of Weedy Rice with Different Densities Stress on the Photosynthesis and Physiological Characteristics of Cultivated Rice
WANG Xiaolin1,ZHANG Xiaofang2,LI Ke2,ZHANG Zhuoya2,LI Gui1
(1.Institute of Plant Protection,Jiangsu Academy of Agricultural Sciences,Nanjing210014,China;2.College of Life Sciences,Nanjing Agricultural University,Nanjing210095,China)
Abstract:In order to investigate the effect of weedy rice stress on grain yield of cultivated rice and its physiological mechanism,weedy rice JS-Y1 and rice cultivar Nanjing 44 were chosen as test materials in field experiment to study the effects of different densities(0,4,8,12,16 plant/m2) of weedy rice stress on photosynthetic characteristics,chlorophyll fluorescence parameters,transmittance,lipids peroxidation and defensive enzyme activities and yield components of cultivated rice.The results showed that with the increase of weedy rice density,weedy rice suppressed the physiological stress indicators of cultivated rice in varying degrees at different growth stages comparing with the control,the transmittance in the height above the soil surface 20,50,80 cm and height below panicles 10 cm were declined by 54.03%-72.49%,39.68%-61.57%,48.98%-54.40% and 38.07%-56.76%,had significant difference compared with their controls,SPAD,photosynthetic parameters,ФPSⅡ,qP,lipids peroxidation and defensive enzyme activities were declined with the different growth stages and the increase of weedy rice density,especially in the middle of filling stages.The production of cultivated rice with densities of 4,8,12,16 plant/m2weedy rice decreased by 16.73%,43.01%,66.61%,83.24%,had significant difference compared to control (no weedy rice planted).It was indicated that the main reasons causing the yield loss of cultivated rice was weedy rice with different densities stress on transmittance of cultivated rice population,caused the cell membrane system decomposed and the declined photosynthetic capacity.
Key words:Weedy rice;Cultivated rice;Photosynthesis;Physiological characteristics
doi:10.7668/hbnxb.2016.01.023
中圖分類號:S511.01
文獻標識碼:A
文章編號:1000-7091(2016)01-0141-06
通訊作者:李貴(1970-),男,安徽巢湖人,研究員,碩士,主要從事植物保護研究。
作者簡介:王曉琳(1986-),女,遼寧營口人,助理研究員,碩士,主要從事雜草生理治理研究。
基金項目:國家自然科學基金項目(31501656;131272080);公益性行業(農業)科研專項經費項目 (201303022);國家“十二五”科技支撐計劃項目(2012BAD19B02)
收稿日期:2015-09-25
