產5α羥化紫杉二烯醇人工酵母的組合設計構建
2016-03-19 07:31:02翟芳宋田青肖文海丁明珠喬建軍元英進
化工學報 2016年1期
翟芳,宋田青,肖文海,丁明珠,喬建軍,元英進
(系統生物工程教育部重點實驗室,天津大學化工學院制藥工程系,天津化學化工協同創新中心,天津 300072)
?
產5α羥化紫杉二烯醇人工酵母的組合設計構建
翟芳,宋田青,肖文海,丁明珠,喬建軍,元英進
(系統生物工程教育部重點實驗室,天津大學化工學院制藥工程系,天津化學化工協同創新中心,天津 300072)
摘要:利用工程化微生物生產天然藥物如紫杉醇在近年來受到研究者廣泛關注。本工作研究如何設計和構建人工酵母以生產紫杉醇生物合成途徑中第一個由細胞色素P450酶催化的羥化產物——5α羥化紫杉二烯醇(taxadien-5α-ol)。利用組合設計原理,選用3種不同紅豆杉來源的紫杉二烯5α羥化酶和2種不同植物源的細胞色素P450還原酶,分別對其進行N端穿膜區域的預測和截短,并對還原酶使用2種不同強度啟動子進行調控,所得羥化酶模塊和還原酶模塊之間分別表達共產生72種組合方式。然后將其分別整合入紫杉二烯生產菌株基因組中,最終在72個組合中篩選到48個可以生產5α羥化紫杉二烯醇的菌株,最高產量67.3 μg·L-1,在釀酒酵母中實現紫杉二烯C5位羥化產物的從頭合成。研究結果表明,利用組合設計策略研究模塊間相互作用及其與底盤間適配性將為在微生物細胞中實現細胞色素P450酶系介導的催化反應過程提供重要參考。
關鍵詞:合成生物學;5α羥化紫杉二烯醇;組合設計;釀酒酵母;生物催化;發酵
2015-06-26收到初稿,2015-08-10收到修改稿。
聯系人:肖文海。第一作者:翟芳(1990—),女,碩士研究生。
Received date: 2015-06-26.
引 言
近年來利用合成生物學構建工程化人工細胞成為生產稀缺植物源的具有復雜結構及獨特性質的化合物的重要手段。研究者通過對已有的生物系統進行改造、設計并構建全新的功能模塊,進行天然化合物的異源生物合成和生產[1],目前已在很多研究方向取得突破性進展[2-15]。Keasling等[2-5]通過系統的模塊化構建及合成路徑調控成為抗瘧疾藥物青蒿素前體生產研究的典范;Ajikumar等[11]開發利用多元模塊代謝工程方法(multivariate-modular approach),使紫杉醇前體紫杉二烯在大腸桿菌中的產量提高了15000倍。
在紫杉醇的生物合成途徑中,5α羥化紫杉二烯醇是紫杉醇母核紫杉二烯下游第一個由細胞色素P450酶催化的羥化產物[16]。該過程是在紫杉二烯5α羥化酶(taxadiene 5α-hydroxylase,T5αOH)和細胞色素P450還原酶(cytochrome P450 reductase,CPR)的共同作用下,催化底物紫杉二烯C5位雙鍵發生遷移并同時引入羥基而完成。隨后5α羥化紫杉二烯醇在酰基轉移酶的作用下乙酰化,再在10β、13α、7β、2α羥化酶的作用下依次進行羥化[9,17]。DeJong 等[14]在釀酒酵母細胞中成功表達了5α羥化酶(T5αOH),5α酰基轉移酶(TAT)和10β羥化酶(T10βOH),并且完成了這些酶類的表征,但沒有實現從IPP到taxdiene-5α-acetate-10β-ol生物合成路徑的表達;Ajikumar等[11]在高產紫杉二烯的研究基礎上,利用穿膜工程在大腸桿菌中融合表達T5αOH和細胞色素P450還原酶,實現5α羥化紫杉二烯醇產量達到58 mg·L-1;最近,Zhou等[15]利用微生物混合培養策略,在大腸桿菌與釀酒酵母中對紫杉醇前體生物合成路徑進行分工,在共培養條件下成功實現氧化紫杉烷化合物的生物合成,其產量達到33 mg·L-1。
目前的研究對細胞色素P450酶催化系統中氧化酶(CYP)和還原酶主要采用分別表達和融合表達兩種方式。分別表達的方式可以實現對氧化酶和還原酶表達強度的分別調控,從而使兩者在轉錄和表達水平上達到協調和相互適配[5];而融合表達策略能夠縮短氧化酶和還原酶的空間距離,從而在一定程度上促進氧化還原電子傳遞反應的高效進行[11]。然而,細胞色素P450酶系介導的生物催化反應過程由于機理較為復雜[17-19],因而如何更有效地選擇氧化酶和還原酶的表達方式和后續優化仍是現階段研究者所面臨的一個難題。
為了解決在釀酒酵母中P450催化5α羥化紫杉二烯醇生物合成這一難題,同時為了對羥化酶和還原酶的表達進行精確調控,本研究將利用組合設計原理,選取3種不同來源的紫杉二烯5α羥化酶和2種不同來源的細胞色素P450還原酶,對其進行跨膜區域不同長度截短,并采用分別表達方式,使用不同強度啟動子進行調控,并將所得羥化酶模塊和還原酶模塊進行組合,以釀酒酵母紫杉二烯生產菌為出發菌株,對上述功能模塊進行基因組特定位點整合,以篩選得到最佳適配的5α羥化紫杉二烯醇生產菌株。本研究以真核細胞釀酒酵母為底盤,相比于原核系統,釀酒酵母具備完整的內膜系統以及與植物細胞類似的電子傳遞系統,是適合作為包含細胞色素P450酶反應過程的復雜天然產物異源合成的宿主平臺;將5α羥化紫杉二烯醇與紫杉二烯生物合成路徑進行整合,能夠避免采用混菌培養系統可能存在的競爭等諸多復雜相互作用關系,同時這也符合后續潛在工業發酵應用對單菌培養偏好的要求;組合設計原理的應用與基因組整合相結合,在基因表達水平上進行微調,有助于實現最佳羥化酶和還原酶的高效篩選和代謝通路優化,可為后續紫杉醇前體生物合成的研究奠定基礎。該研究對構建其他天然化合物的生物合成途徑或由細胞色素P450酶系介導的催化反應過程提供重要的參考。
1 實驗材料和方法
1.1 基因元件載體和基因表達盒的構建
設計特異性引物,以釀酒酵母單敲菌株YNL280C (erg24Δ::kanMX4)(購自Open Biosystems ,Huntsville,AL)的基因組為模板擴增得到KanMX基因的上游部分片段kmx1、組氨酸營養標記基因his3和終止子eno2t,通過重疊延伸PCR(OE-PCR)將3個片段連接,用限制性內切酶Pst I/Bam HI酶切,酶切產物經純化后連接到載體pRS425K[20]的相應位點,得到元件載體pRS425K-kmx1-his3-eno2t。用相同的方法構建載體pRS425K-eno2t-tef1p-gpm1t、pRS425K-gpm1t-pgi1p-gpdt-kmx2和pRS425K-gpm1t-tdh3pgpdt-kmx2,且其中啟動子和第二個終止子之間留有Pme I酶切位點,用于紫杉二烯5α羥化酶或還原酶基因的連接,圖1(a)所示為以zf11為例,說明元件載體的構建過程。

圖1 基因表達盒zf11-T5αOH的構建過程Fig.1 Construction of gene cassette zf11-T5αOH
紫杉二烯5α羥化酶基因Tw5αOH(Taxus wallichiana var. chinensis)、Tm5αOH(Taxus x media)、Tc5αOH(Taxus cuspidata)和P450還原酶基因TcCPR (Taxus cuspidata)、AtCPR(Arabidopsis thaliana)均由金唯智公司依照釀酒酵母密碼子偏好性對序列進行優化后合成。在http://www.cbs.dtu.dk/services/ TMHMM/網站中預測上述基因的穿膜區域。設計特異性引物,擴增不同長度基因片段,用CPEC方法[21]將基因片段分別連入相應的元件載體中得到完整的基因表達盒[圖1(b)]。所有模塊均經過測序驗證堿基序列的正確性。表1所示為本實驗所構建的質粒。

表1 實驗中涉及的質粒Table 1 Plasmids involved in this study
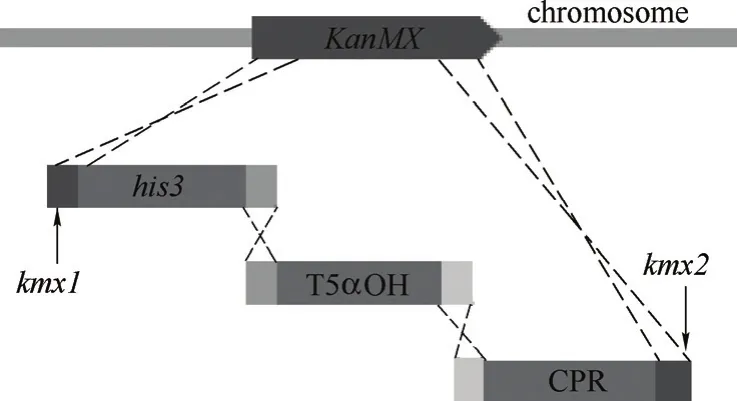
圖2 5α羥化紫杉二烯醇合成模塊的基因組整合Fig.2 Genomic integration of taxadien-5α-ol modules
1.2 酵母轉化和篩選
用限制性內切酶Pst I/Bam HI對元件載體zf01以及所有基因表達盒進行酶切,凝膠回收得到元件模塊和基因表達盒模塊。將相應的羥化酶基因模塊與還原酶基因模塊組合,同元件模塊zf01按等物質的量混合,采用醋酸鋰法轉化釀酒酵母紫杉二烯生產菌株SyBE_001051[13]。模塊之間以終止子片段作為同源臂連接,兩端通過KanMX位點同源序列與基因組該位點發生雙交換同源重組(圖2)。使用SD-ura-leu固體培養基進行篩選,于30℃培養2~3d,挑取單菌落在SD-ura-leu固體培養基上劃線分純,挑取分純后的單菌落進行酵母菌落PCR,通過驗證目的片段存在且大小正確,從而確定完整組裝的陽性克隆。
1.3 人工酵母菌株的發酵
將100 μl液體甘油菌接入5 ml SD-ura-leu液體培養基中,30℃、220 r·min-1培養24 h后轉接入5 ml SD-ura-leu液體培養基中,使其初始OD600為0.2,在30℃、220 r·min?1條件下培養至菌體對數生長中期(OD600約為5),接入50 mlYPD培養基中,使其初始OD600為0.1,在30℃、220 r·min-1條件下發酵,10 h時加入5%(體積)正十二烷進行兩相培養,70 h結束發酵,將發酵液于11000 r·min-1離心5 min,收集上層有機相,除水過濾后備用GC-MS測定產物。
1.4 5α羥化紫杉二烯醇的檢測與定量
使用三重四極氣質聯用系統(Agilent公司)測定產物。色譜條件:HP-5ms 30 m×0.25 μm×0.25 mm硅膠毛細管色譜柱(Agilent公司);恒流模式,載氣氦氣流速為1 ml·min-1;進樣量10 μl。柱溫起始溫度100℃,保持1 min,以15℃·min-1升至175℃;再以4℃·min-1升至220℃,保持2 min;最后以50℃·min-1升至290℃,保持1 min;進樣口溫度與色譜-質譜接口溫度均為250℃。質譜條件:電子轟擊電離EI+;電子束能量70 eV;離子源溫度250℃;溶劑延遲時間7 min;質量掃描方式:全離子掃描與SIM選擇離子掃描。
采用與5α羥化紫杉二烯醇結構最為相近的化合物紫杉二烯的純品繪制標準曲線,通過外標法對5α羥化紫杉二烯醇進行半定量。
2 實驗結果與討論
2.1 5α羥化紫杉二烯醇生物合成中組合設計原理的應用
在紫杉醇生物合成途徑中,5α羥化紫杉二烯醇是紫杉二烯下游第一個由細胞色素P450單加氧酶催化的羥化產物,該催化反應由紫杉二烯5α羥化酶與細胞色素P450還原酶通過一系列電子傳遞過程共同完成。由于P450酶在異源宿主中都是通過膜結合而表達并發揮功能,因此相比于原核生物大腸桿菌,具有與植物細胞類似的細胞色素P450電子傳遞系統和完整內膜系統的真核生物釀酒酵母更加適合作為5α羥化紫杉二烯醇及其下游一系列細胞色素P450介導的氧化催化產物合成的宿主細胞,圖3所示為在釀酒酵母紫杉二烯生物合成模塊基礎上,引入5α羥化紫杉二烯醇相關基因模塊的生物合成路徑。
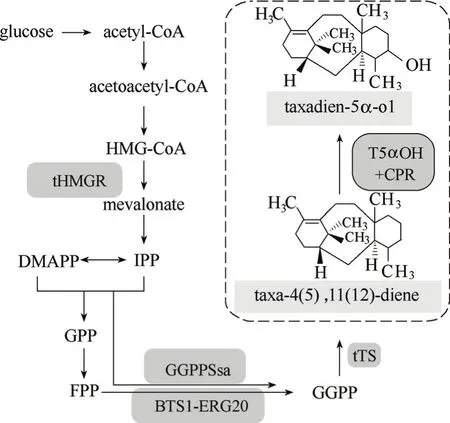
圖3 工程釀酒酵母中5α羥化紫杉二烯醇生物合成路徑Fig.3 Biosynthesis pathway of taxadien-5α-ol in engineered S. cerevisiaeHMG-CoA—3-hydroxy-3-methylglutaryl coenzyme A; DMAPP—dimethylallyl diphosphate; IPP—isopentenyl pyrophosphate; GPP—geranyl diphosphate; FPP—farnesyl diphosphate; GGPP—geranylgeranyl diphosphate; tHMGR—HMG-CoA reductase (truncated version); GGPPSsa—GGPP synthase of Sulfolobus acidocaldarius; BTS1-ERG20—fusion gene of BTS1 (GGPP synthase of yeast) and ERG20 (FPP synthase of yeast); tTS—taxadiene synthase of Taxus brevifolia (truncated version); T5αOH—taxadiene 5α-hydroxylase of Taxus; CPR—cytochrome P450 reductase
為了研究紫杉二烯5α羥化酶和還原酶的協同表達,選擇3種不同紅豆杉品種來源的羥化酶,分別是中國紅豆杉源、曼地亞紅豆杉源和東北紅豆杉源,以及兩種不同物種來源的還原酶,分別是東北紅豆杉源和擬南芥源。Croteau等[22]的研究表明,紅豆杉來源的不同P450單加氧酶間序列相似性很高,而與其他植物來源羥化酶序列差異較大,推斷其可能具有較高底物專一性和特殊催化特性,因此本研究僅選取紅豆杉源的羥化酶進行研究;此外,釀酒酵母中雖然具備細胞色素P450氧化還原系統,但其還原酶(CPR)的表達量低且與植物P450的電子傳遞效率不匹配[23-24],因此外源CPR的引入是必要的,它作為電子傳遞體與羥化酶之間發生電子傳遞作用,而不直接作用于底物,因此選取了2種不同物種來源的還原酶進行研究。
通過http://www.cbs.dtu.dk/services/TMHMM/網站預測得到不同來源羥化酶的穿膜區均為N端42個氨基酸,對其進行穿膜區24個氨基酸的部分截短以及全部截短,共得到9個羥化酶基因片段:Tw5αOH、t24Tw5αOH、t42Tw5αOH、Tm5αOH 、t24Tm5αOH、t42Tm5αOH、Tc5αOH、t24Tc5αOH、t42Tc5αOH;而紅豆杉源和擬南芥源還原酶的穿膜區分別為N端74個和46個氨基酸,對其進行全部截短,共得到4個還原酶基因片段:TcCPR、t74TcCPR、AtCPR、t46AtCPR。此外,有研究還指出,紫杉二烯5α羥化酶的活性和催化效率較低是造成紫杉二烯向下游代謝物轉化的代謝瓶頸[14],因此本研究對羥化酶基因使用釀酒酵母組成型強啟動子tef1p進行調控以獲得其高表達量,對還原酶基因使用強啟動子tdh3p和弱啟動子pgi1p分別進行調控,以研究并獲得與羥化酶相匹配的還原酶表達水平。利用組合設計原理,如圖4所示,將所述羥化酶與還原酶模塊進行組合,共得到72種不同組合的5α羥化紫杉二烯醇生物合成模塊。
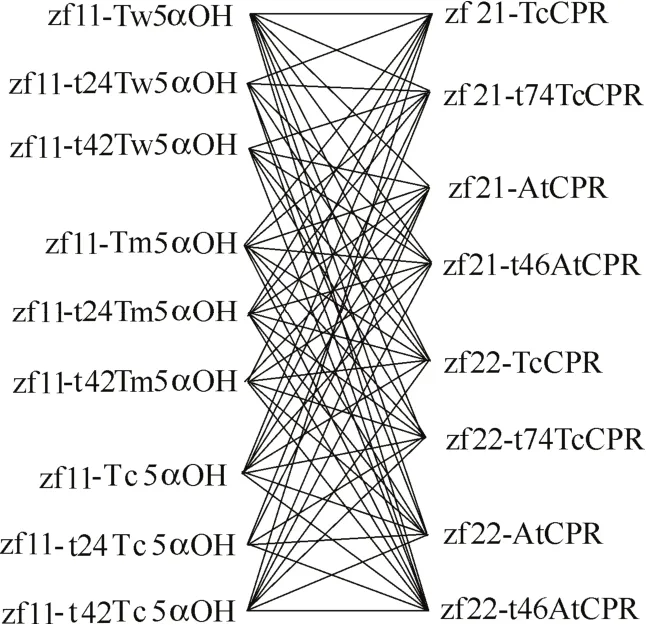
圖4 紫杉二烯5α羥化酶與細胞色素P450還原酶的組合設計Fig.4 Combinatorial design of taxadiene 5α-hydroxylase and CYP450 reductase
2.2 產5α羥化紫杉二烯醇人工酵母的構建與發酵
本研究所采用的出發菌株為SyBE_001051,它是以釀酒酵母單敲菌株YNL280C(敲除麥角固醇合成途徑中關鍵基因erg24,從而有效降低競爭支路的代謝通量)為底盤,整合入含有紫杉二烯生物合成途徑關鍵基因tHMGR、GGPPSsa、BTS1-ERG20、tTS的著絲粒質粒,所構建的紫杉二烯生產菌株,其搖瓶發酵紫杉二烯產量達75 mg·L-1[13]。本研究將組合設計中相應的羥化酶模塊和還原酶模塊進行基于該出發菌株基因組Kan位點的整合共表達,分別從每個組合中選取3個陽性克隆進行發酵與GC-MS測定。
發酵樣品的總離子流檢測結果如圖5(a)所示,可以看出,發酵產物種類較復雜,對其中具有離子特征峰m/z 288[272(紫杉二烯)+16(氧原子)]的色譜峰進行質譜分析,并依據其特征峰進行SIM模式下的選擇離子掃描,結果如圖5(b)所示,保留時間為17.03 min的色譜峰,其離子特征峰[圖6(a)]與文獻報道的5α羥化紫杉二烯醇[16]完全吻合,判斷為目標產物;保留時間為16.50 min的色譜峰,其離子特征峰[圖6(b)]與文獻報道的5(12)-oxa-3(11)-cyclotaxane(OCT)[16]完全吻合,判斷為副產物OCT;其他3個同樣具有離子m/z 288的色譜峰,可能為其他紫杉烷化合物或氧化產物[圖6(c)~(e)],產量很低,未能進行分離純化,因此目前不能對其定性。
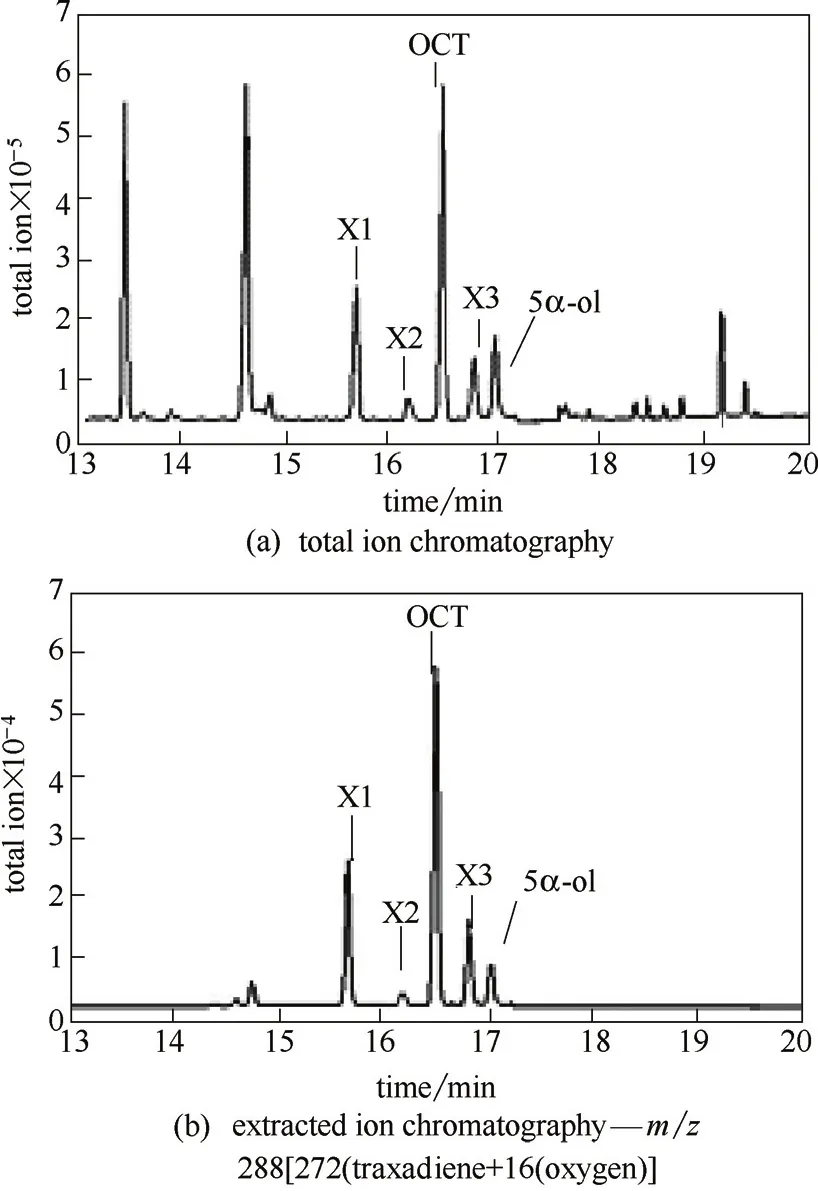
圖5 菌株t24Tc5αOH-zf22t74TcCPR的發酵GC-MS檢測Fig.5 GC-MS profile for strain t24Tc5αOH-zf22t74TcCPR
利用紫杉二烯標準曲線對5α羥化紫杉二烯醇進行半定量,全部菌株的產量如表2所示。
5α羥化紫杉二烯醇產量最高的菌株為62號,它是通過N端截短24個氨基酸的東北紅豆杉源羥化酶與N端截短74個氨基酸的東北紅豆杉源還原酶組合并構建而得,且還原酶與羥化酶基因均為強啟動子調控,由此推斷,雖然不同來源還原酶均可在該體系中發揮電子傳遞作用,但紅豆杉源羥化酶與同一來源的還原酶適配性優于異源還原酶,且與羥化酶相當的還原酶轉錄和表達水平在此體系中更優,因而更能發揮其最佳催化效力。
2.3 羥化酶的不同來源和截短形式對5α羥化紫杉二烯醇生物合成的影響
對5α羥化紫杉二烯醇進行產量比較發現,使用不同來源的羥化酶,目標產物含量差異較為明顯,東北紅豆杉源羥化酶的效果優于另外兩者(圖7)。經序列比對發現,3種羥化酶的氨基酸序列相似性在98%以上,而曼地亞紅豆杉源和東北紅豆杉源羥化酶只有一個氨基酸差異,為N端31位,位于所預測的蛋白穿膜區域,曼地亞紅豆杉源蛋白序列中該位點為絲氨酸,而東北紅豆杉源蛋白序列中該位點為丙氨酸,推斷不同極性氨基酸的差異可能會導致蛋白空間結構的不同,進而影響了酶的催化活性,有待對蛋白結構及其與底物間相互作用進行模擬以進一步揭示其催化作用機制。
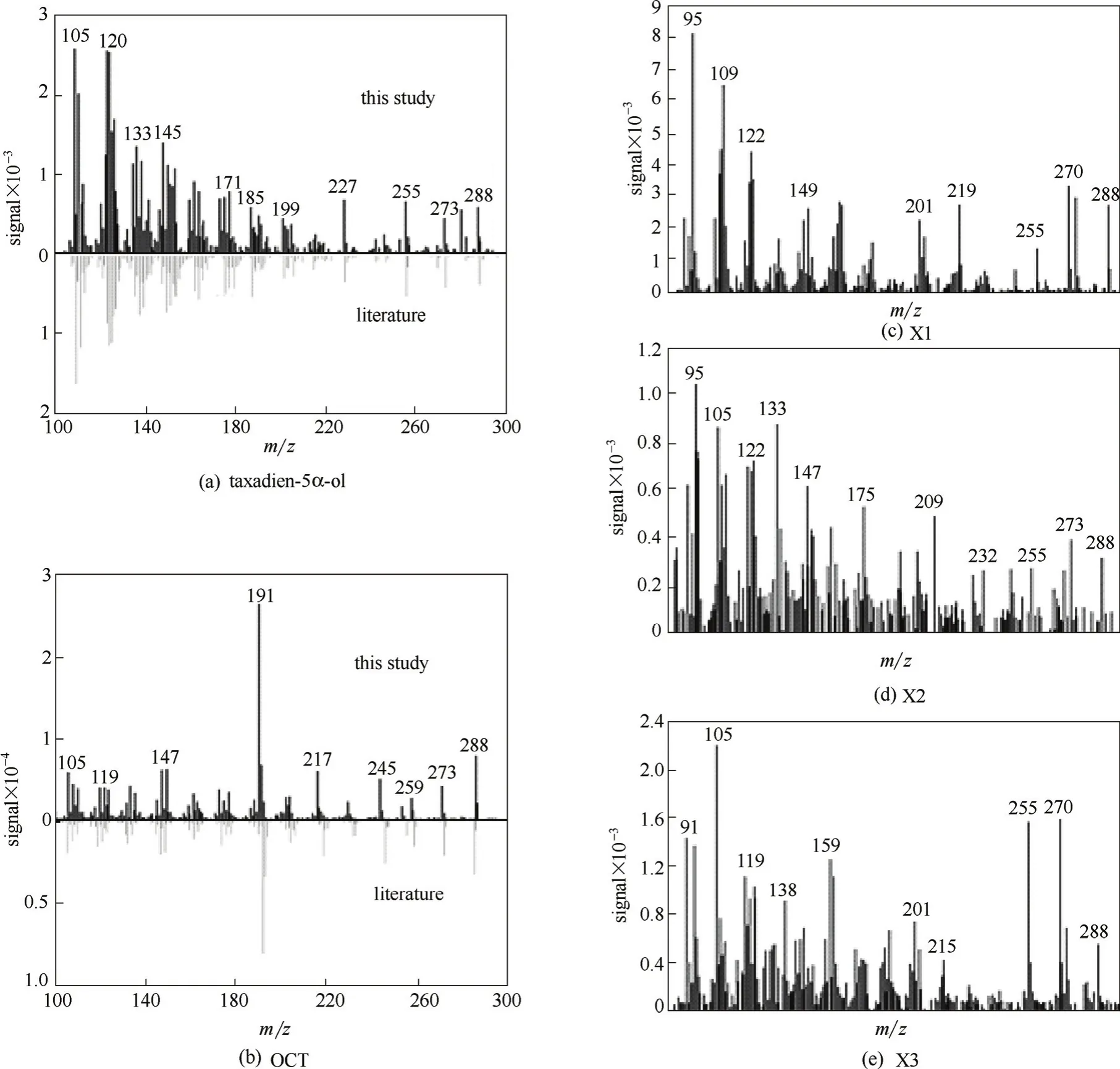
圖6 紫杉烷化合物的質譜檢測圖Fig.6 Mass spectra of taxanes
此外,使用未截短或截短N端24個氨基酸的羥化酶,均有不同程度C5位羥化產物的產出,而羥化酶N端42個氨基酸被截短后,均不能合成羥化產物,推斷植物源P450酶可由其本身的N端跨膜區定位于酵母內質網上而表達并發揮功能,這是酵母作為真核細胞用于表達異源蛋白相對于原核宿主的優勢之一[25];而截短42個氨基酸可能使其不能進行膜定位,影響了蛋白的正確折疊和催化活性區域空間結構的形成,從而導致其功能遭到破壞。
3 結 論
(1)利用組合設計策略將不同來源、不同截短形式以及不同啟動子調控的紫杉二烯5α羥化酶和P450還原酶進行組合和分別表達,在72種組合中篩選得到48個產5α羥化紫杉二烯醇的釀酒酵母菌株,產量最高為67.3 μg·L-1,其對應的最佳組合為東北紅豆杉源羥化酶(t24)與東北紅豆杉源還原酶(t74)組合,且羥化酶基因和還原酶基因均為強啟動子調控。因而在后續的研究中,可以嘗試在釀酒酵母中對羥化酶和還原酶采取融合表達策略,對兩者進行統一調控,通過酶的空間距離的調節來研究其對催化目標產物合成的影響。
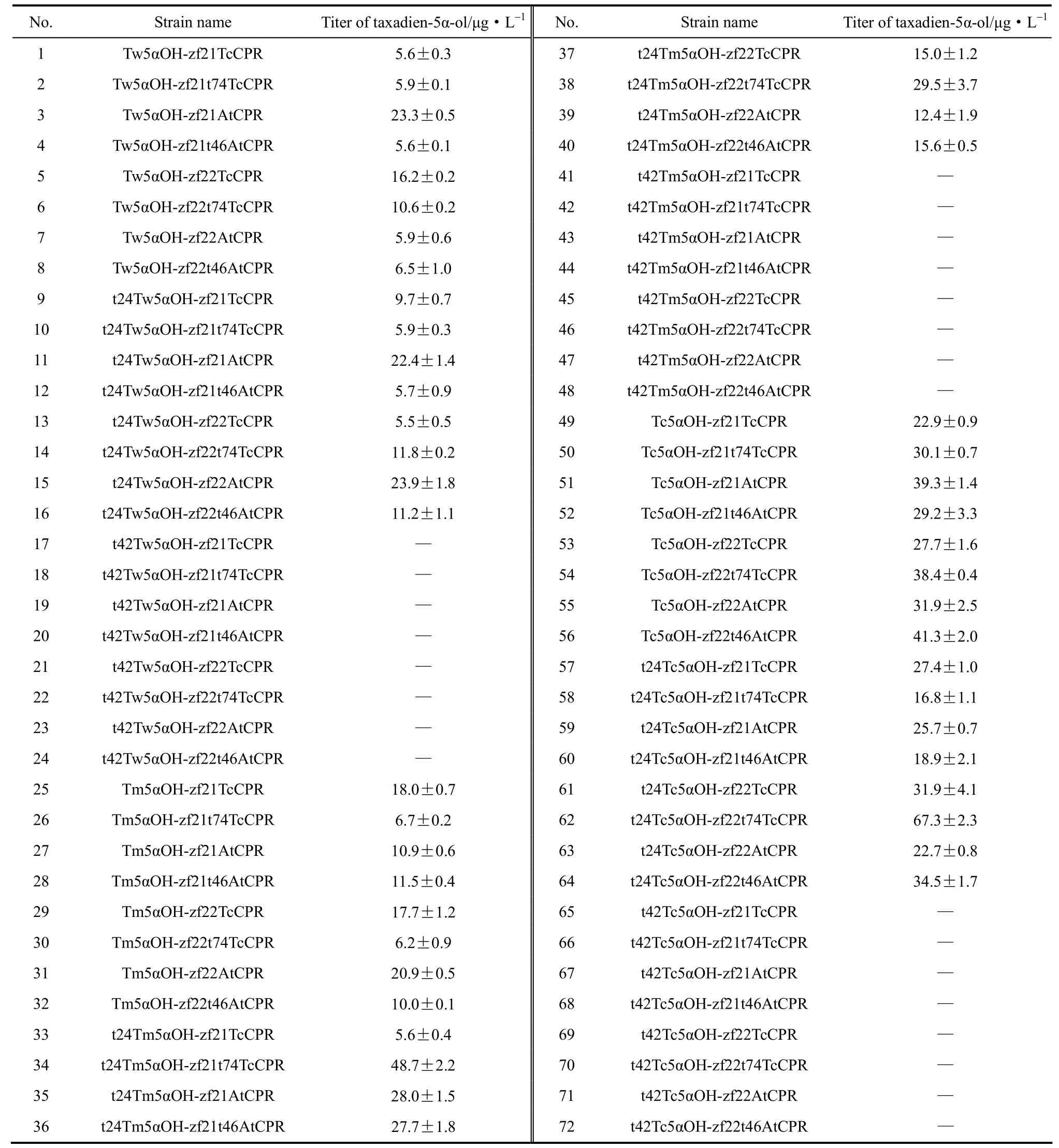
表2 本研究所構建菌株5α羥化紫杉二烯醇產量Table 2 Titer of taxadien-5α-ol of all strains constructed in this study
(2)在產紫杉二烯的釀酒酵母底盤中共表達紫杉二烯5α羥化酶和P450還原酶,得到的氧化產物并不單一,除目標產物5α羥化紫杉二烯醇外,還包含副產物OCT以及多個(本研究中為3個)未知的紫杉二烯羥化物,統稱為紫杉烷類化合物。
(3)在所選用的3種不同來源的紫杉二烯5α羥化酶中,東北紅豆杉源羥化酶催化活性最高,其次為曼地亞紅豆杉源羥化酶,而中國紅豆杉源羥化酶活性較差。
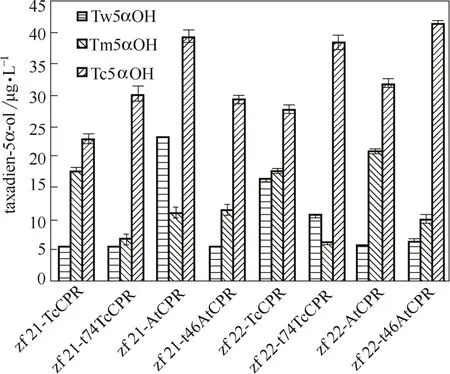
圖7 不同來源羥化酶對5α羥化紫杉二烯醇產量的影響Fig.7 Effect of different taxadien 5α-hydroxylase on taxadien-5α-ol production
(4)在釀酒酵母中表達全長或N端截短24個氨基酸殘基的紫杉二烯5α羥化酶,均可以得到5α羥化紫杉二烯醇及其他羥化產物,而表達N端截短42個氨基酸殘基的羥化酶,則不能得到任何羥化產物。后續可借助分子模擬等手段分析羥化酶的膜結合及其與底物紫杉二烯的相互作用方式,用于指導對羥化酶的定向進化等改造,提高其催化效率和產物專一性,進而提高該催化過程的轉化率,提高5α羥化紫杉二烯醇產量。
References
[1] 劉奪, 杜瑾, 趙廣榮, 等. 合成生物學在醫藥及能源領域的應用[J].化工學報, 2011, 62 (9): 2391-2397. LIU D, DU J, ZHAO G R, et al. Applications of synthetic biology in medicine and energy [J]. CIESC Journal, 2011, 62 (9): 2391-2397.
[2] RO D K, PARADISE E M, OUELLET M, et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast [J]. Nature, 2006, 440 (7086): 940-943.
[3] RO D K, OUELLET M, PARADISE E M, et al. Induction of multiple pleiotropic drug resistance genes in yeast engineered to produce an increased level of anti-malarial drug precursor, artemisinic acid [J]. BMC Biotechnol., 2008, 8: 83.
[4] WESTFALL P J, PITERA D J, LEIHAN J R, et al. Production of amorphadiene in yeast, and its conversion to dihydroartemisinic acid, precursor to the antimalarial agent artemisinin [J]. PNAS, 2012, 109 (3): 111-118.
[5] PADDON C J, KEASLING J D. Semi-synthetic artemisinin: a model for the use of synthetic biology in pharmaceutical development [J]. Nature Reviews, 2014, 12 (5): 335-367.
[6] ZHOU Y J, GAO W, RONG Q X, et al. Modular pathway engineering of diterpenoid synthases and the mevalonic acid pathway for miltiradiene production [J]. J. Am. Chem. Soc., 2012, 134 (6): 3234-3241.
[7] DAI Z B, LIU Y, HUANG L Q, et al. Production of miltiradiene by metabolically engineered Saccharomyces cerevisiae [J]. Biotechnology and Bioengineering, 2012, 109 (11): 2845-2853.
[8] CARRAU F M, MEDINA K, BOIDO E, et al. De novo synthesis of monoterpenes by Saccharomyces cerevisiae wine yeasts [J]. FEMS Microbiol. Lett., 2005, 243 (1): 107-115.
[9] JENNEWEIN S, RITHNER C D, WILLIAMS R M, et al. Taxol biosynthesis: taxane 13 alpha-hydroxylase is a cytochrome P450-dependent monooxygenase [J]. PNAS, 2001, 98 (24): 13595-135600.
[10] WANG H H, ISAACS F J, CARR P A, et al. Programming cells by multiplex genome engineering and accelerated evolution [J]. Nature, 2009, 460 (7257): 894-898.
[11] AJIKUMAR P K, XIAO W H, TYO K E, et al. Isoprenoid pathway optimization for Taxol precursor overproduction in Escherichia coli [J]. Science, 2010, 330 (6000): 70-74.
[12] DING M Z, YAN H F, LI L F, et al. Biosynthesis of taxadiene in Saccharomyces cerevisiae: selection of geranylgeranyl diphosphate synthase directed by a computer-aided docking strategy [J]. PLoS ONE, 2014, 9 (10): e109348.
[13] 張正偉, 丁明珠, 元英進. 紫杉二烯生物合成模塊與不同底盤的適配 [J].高等學校化學學報, 2014, 35 (1): 75-79. ZHANG Z W, DING M Z, YUAN Y J. A research about the fitness of the taxadiene biosynthetic modules with different S. cerevisiae chassis [J]. Chemical Journal of Chinese Universities, 2014, 35 (1): 75-79.
[14] DEJONG J M, LIU Y, BOLLON A P, et al. Genetic engineering of taxol biosynthetic genes in Saccharomyces cerevisiae [J]. Biotechnology and Bioengineering, 2006, 93 (2): 212-224.
[15] ZHOU K, QIAO K J, EDGAR S, et al. Distributing a metabolic pathway among a microbial consortium enhances production of natural products [J]. Nature Biotechnology, 2015, 33 (4): 377-383.
[16] JENNEWEIN S, LONG R, WILLIAMS R M, et al. Cytochrome P450 taxadien 5α-hydroxylase, a mechanistically unusual monooxygenase catalyzing the first oxygenation step of Taxol biosynthesis [J]. Chem. Biol., 2004, 11 (3): 379-387.
[17] CHAU M, CROTEAU R. Molecular cloning and characterization of a cytochrome P450 taxoid 2alpha-hydroxylase involved in Taxol biosynthesis [J]. Arch. Biochem. Biophys., 2004, 427 (1): 48-57.
[18] RONTEIN D, ONILLON S, HERBETTE G, et al. CYP725A4 from yew catalyzes complex structural rearrangement of taxa-4(5),11(12)-diene into the cyclic ether 5(12)-oxa-3(11)-cyclotaxane [J]. J. Biol. Chem., 2008, 283 (10): 6067-6075.
[19] GUERRA-BUBB J, CROTEAU R, WILLIAMS R M. The early stages of Taxol biosynthesis: an interim report on the synthesis and identification of early pathway metabolites [J]. Nat. Prod. Rep., 2012, 29 (6): 683-696.
[20] 郭睿, 丁明珠, 元英進. 產青蒿二烯的人工酵母細胞的構建及發酵優化 [J].化工學報, 2015, 66 (1): 378-385. GUO R, DING M Z, YUAN Y J. Construction of artificial yeast cell for producing amorphadiene, and optimization of fermentation [J]. CIESC Journal, 2015, 66 (1): 378-385.
[21] QUAN J Y, TIAN J D. Circular polymerase extension cloning of complex gene libraries and pathways [J]. PLoS ONE, 2009, 4 (7): e6441.
[22] KASPERA R, CROTEAU R. Cytochrome P450 oxygenases of Taxol biosynthesis [J]. Phytochemistry Reviews, 2006, 5 (2/3): 433-444.
[23] JENNEWEIN S, PARK H, DEJONG J M, et al. Coexpression in yeast of Taxus cytochrome P450 reductase with cytochrome P450 oxygenases involved in Taxol biosynthesis [J]. Biotechnology and Bioengineering, 2005, 89 (5): 588-598.
[24] 李杰, 王春梅. 紫杉醇組合生物合成的研究進展 [J].生物工程學報, 2014, 30 (3): 355-367. LI J, WANG C M. Advances of taxol combinatorial biosynthesis [J]. Chinese Journal of Biotechnology, 2014, 30 (3): 355-367.
[25] HAMANN T, M?LLER B L. Improved cloning and expression of cytochrome P450s and cytochrome P450 reductase in yeast [J]. Protein Expression and Purification, 2007, 56 (1): 121-127.
Foundation item: supported by the National High Technology Research and Development Program of China (2012AA02A701) and the Foundation for the Author of National Excellent Doctoral Dissertation of China (201456).
Combinatorial design and construction of artificial yeast for production of taxadien-5α-ol
ZHAI Fang, SONG Tianqing, XIAO Wenhai, DING Mingzhu, QIAO Jianjun, YUAN Yingjin
(Key Laboratory of Systems Bioengineering, Ministry of Education; Department of Pharmaceutical Engineering, School of Chemical Engineering and Technology, Tianjin University; Collaborative Innovation Center of Chemical Science and Engineering (Tianjin), Tianjin 300072, China)
Abstract:Producing natural drugs such as Taxol by engineered microbes have attracted extensive attention in recent years. This work focused on how to design and construct artificial yeast for the production of taxadien-5α-ol, which was the first hydroxylated product catalyzed by cytochrome P450 enzymes in Taxol biosynthesis pathway. Based on combinatorial design, three hydroxylase genes derived from different yew subspecies and two reductase genes from different organisms with N-terminal truncatd with various length were tested simultaneously to investigate the fitness between the genes and the chassis. Two kinds of promoters were used for regulating the cytochrome P450 reductase genes. In total, 72 kinds of combinations were constructed and then integrated into the chromosome of the taxadiene producing strain. 48 taxadine-5α-ol producing strains were obtained out of 72 combinations by co-expression of the hydroxylase and the reductase. The highest titer was 67.3 μg·L-1. This was the first time to realize the biosynthesis of taxadine-5α-ol form glucose in Saccharomyces cerevisiae. The result provided a good reference for the research of the catalytic reaction mediated by cytochromebook=316,ebook=324P450 enzymes in microbes through the interactions between modules and chassis by combinatorial design.
Key words:synthetic biology; taxadien-5α-ol; combinatorial design; Saccharomyces cerevisiae; biocatalysis; fermentation
Corresponding author:XIAO Wenhai, wenhai.xiao@tju.edu.cn
基金項目:國家高技術研究發展計劃項目(2012AA02A701);高等學校全國優秀博士學位論文作者專項基金(201456)。
中圖分類號:Q 819
文獻標志碼:A
文章編號:0438—1157(2016)01—0315—09
DOI:10.11949/j.issn.0438-1157.20150992
