氮、磷、鉀對鹽脅迫谷子幼苗形態和生理指標的影響
2016-04-11 23:36:42呼紅梅王莉
江蘇農業科學 2016年2期
呼紅梅++王莉

摘要:以盆栽谷子晉谷21號為試驗材料,采用“3414”不完全正交試驗設計,從植物生長狀況、滲透調節物質、保護酶活性等研究氮(N)、磷(P)、鉀(K)對鹽脅迫谷子幼苗形態及生理指標的影響,探索營養元素N、P、K含量及配比與谷子抗鹽性的關系。結果表明,谷子幼苗在輕度鹽脅迫下施用N、P、K分別為100、87、249 mg/kg,中度鹽脅迫下施用N 100 mg/kg,P 43.5、87.0 mg/kg,K 124.5、249 mg/kg,重度鹽脅迫下施用N、P、K分別為100、43.5、124.5 mg/kg有利于植物生長;N、P、K配合施用,其交互作用效果更利于谷子生長而提高抗鹽性,但適宜的N、P、K肥料用量范圍變小;N、P、K用量過高會造成鹽害,不利于谷子抗鹽。
關鍵詞:谷子;幼苗;鹽脅迫;形態指標;生理指標
中圖分類號: S515.062文獻標志碼: A文章編號:1002-1302(2016)02-0117-05
收稿日期:2015-03-02
基金項目:國家科技支撐計劃重點項目(編號:2008BADA4B01);山西省引進優秀人才專項基金(2010);山西省留學基金(2011)。
作者簡介:呼紅梅(1987—),女,山西呂梁人,碩士,主要從事植物生理生態學研究。E-mail:hhm135527@126.com。
通信作者:王莉,副教授,碩士生導師,主要從事植物生理生態學研究。E-mail:wangli11882003@126.com。目前,我國耕地面積約為1.35億 hm2[1],其中鹽堿土占耕地面積的 20%,主要分布于土壤蒸發量大、降水量少的干旱、半干旱地區及濱海地區[2],這些地區也是我國谷子種植分布較多的地域。開發鹽堿地,提高作物的抗鹽性,對補充我國耕地資源、確保國家糧食安全具有不可替代的作用。
谷子[Setaria italic (L.) Beauv.]古稱粟,是禾本科狗尾草屬的1個栽培種,對土壤鹽分中度敏感,土壤鹽脅迫常常造成谷子萌發率低、出苗不全、幼苗長勢弱等現象,嚴重影響谷子的產量和經濟效益,研究鹽脅迫對谷子幼苗生長的影響對谷子生產具有重要意義[3-4]。氮(N)、磷(P)、鉀(K)作為作物生長的3大主要營養元素,對提高作物的抗鹽性有重要作用。有研究表明,鹽脅迫下適宜的氮肥可以改善馬鈴薯的光合能力,顯著提高馬鈴薯脫毒苗葉片的SOD和POD活性[5];適量的P、K肥能提高NaCl脅迫下玉米苗期葉片中的葉綠素含量和干物質量,增強滲透調節能力和POD、SOD、CAT活性,使MDA含量降低[6];增施磷肥能促進黑麥草幼苗生長,增加干物質積累,提高葉片葉綠素含量,增加可溶性糖、可溶性蛋白質及脯氨酸含量[7];適量的鉀肥,可提高菊芋幼苗在鹽境下的生物量,促進光合作用[8],馬鈴薯脫毒苗葉片的脯氨酸含量顯著提高[9],玉米幼苗葉片和根系中MDA 含量降低,POD、SOD、CAT 活性增加[10],NaCl脅迫下冬小麥幼苗地上部和地下部的生長、可溶性糖含量、抗氧化酶的活性提高,電解質外滲量和MDA含量降低[11]。目前,NaCl脅迫下,以N、P、K單一養分對作物抗鹽性的研究較多,且多集中于玉米、大麥、小麥、馬鈴薯、甜高粱等作物[5,9-13],對谷子耐鹽性的研究報道甚少,主要集中于谷子發芽期、幼苗前期耐鹽性鑒定指標研究、耐鹽品種和基因型的篩選、鹽脅迫對谷子農藝性狀及產量的影響[3-4,14-15],施肥對鹽脅迫下谷子幼苗的影響尚未見報道。本試驗通過研究N、P、K對鹽脅迫下谷子幼苗形態和生理指標的影響,分析N、P、K含量及配比與谷子幼苗抗鹽性的關系,為合理施肥提高谷子抵抗鹽脅迫能力提供理論依據。
1材料與方法
1.1試驗材料
試驗于2014年5月在山西師范大學生命科學學院實驗室內進行,采用直徑、高分別為9、12.5 cm的盆播種谷子,每盆裝沙500 g,供試沙子經清水反復洗滌,幾乎不含有養分。供試谷子品種為晉谷21號。供試肥料為:Ca(NO3)2·4H2O,含N 11.9%;KH2PO4,含P2O5 52.1%、K2O 34.6%;K2SO4,含K2O 54.1%。
1.2試驗設計
試驗采用農業部推薦的“3414”不完全正交設計(表1),最佳施肥量為N ∶P2O5 ∶K2O=1 ∶0.67 ∶1,2/3的氮肥、全部磷肥和鉀肥一次性施入沙中。另外,加入B、Mn、Zn、Cu、Mo、Fe、Mg分別為0.5、0.5、0.05、0.02、0.01、5.6、0.5 mg/kg。設無鹽脅迫、輕度鹽脅迫、中度鹽脅迫、重度鹽脅迫4個脅迫處理水平,沙中NaCl用量分別為0(CK)、0.4、0.8、1 g/kg。
試驗時,挑選籽粒成熟且飽滿的種子,用1% KMnO4溶液浸泡10 min進行消毒,用蒸餾水清洗干凈,備用;每盆播種30粒,置于晝夜溫度分別為30、20 ℃、光照15 h/d的人工氣候培養箱中進行培養;出苗后,留苗10株培養45 d,取樣調查測定相關指標。土壤水分控制采用稱質量法,每盆保持最大田間持水量的60%。
1.3指標測定與方法
幼苗株高、葉面積分別采用直接測量法、AM 300型葉面積儀進行測量;相對葉綠素含量用CM-1000葉綠素儀直接測定;莖、葉和根干物質的質量測定,先將莖、葉和根烘干,采用分析天平稱量;可溶性糖、可溶性蛋白質、脯氨酸含量分別
采用蒽酮比色法、考馬斯亮藍G-250染色法、磺基水楊酸法[16]測定;過氧化物酶(POD)、超氧化物歧化酶(SOD)、過氧化氫酶(CAT)活性分別采用愈創木酚法、氮藍四唑(NBT)光化還原法、紫外吸收法[17]測定;丙二醛(MDA)含量采用硫代巴比妥酸法[18]測定。
1.4數據處理
用Excel 2003處理數據和作圖;用SPSS 16.0在α=0.05水平上對試驗結果進行方差分析和Duncans多重比較。
2結果與分析
2.1N、P、K對鹽脅迫谷子幼苗生長的影響
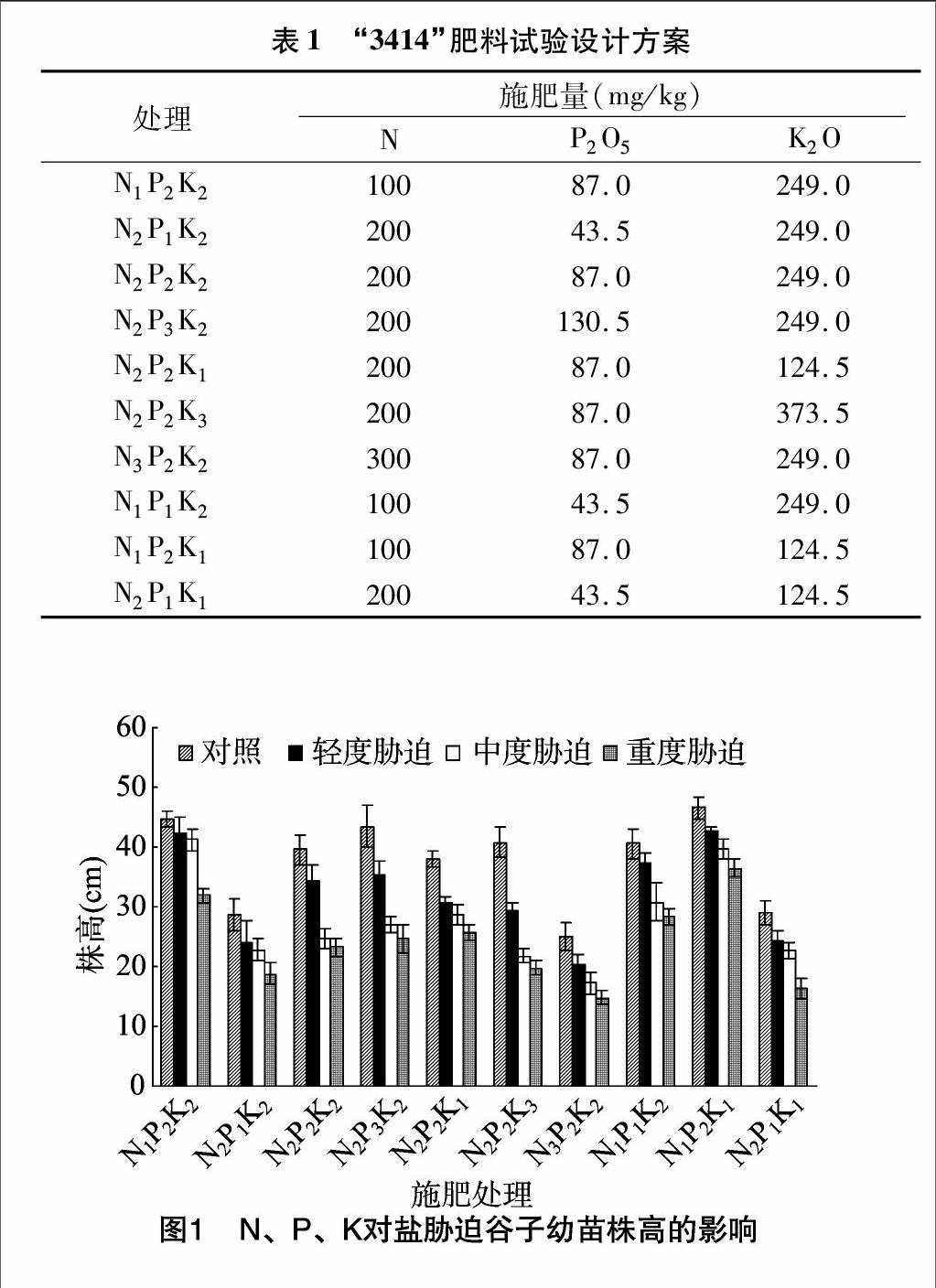
由圖1至圖4可知,相同P2K2水平下,3種鹽處理隨施N量的增加,谷子幼苗株高、葉面積、葉綠素含量、干物質質量呈減小趨勢,且差異顯著(P<0.05,下同),與無鹽脅迫(CK)相比,N1水平的谷子幼苗株高、葉面積、葉綠素含量、干物質質量降幅相對最小,輕度、中度與重度鹽脅迫的株高降幅分別為5.7%、7.9%、28.8%,葉面積降幅分別為9.5%、22.2%、40.7%,葉綠素含量降幅分別為4.7%、14.1%、20.3%,干物質質量降幅分別為25.8%、31.1%、46.0%。
相同N2K2水平下,3種鹽處理隨施P量的增加,谷子幼苗株高、葉面積呈增加趨勢,葉綠素含量呈減小趨勢;輕度鹽脅迫下,谷子幼苗P2、P3株高大于P1,P1、P2葉綠素含量大于P3,葉面積大小為P3>P2>P1,差異顯著;中度、重度鹽脅迫下,谷子幼苗的株高大小為P3>P2>P1,P3葉面積大于P1、P2,P1葉綠素含量大于P2、P3,差異顯著;輕度與中度鹽脅迫下,隨施P量的增加干物質質量呈先增加后減小趨勢,輕度鹽脅迫下P2干物質質量大于P1、P3,中度鹽脅迫下P1、P2干物
質質量大于P3,差異顯著;重度鹽脅迫下,隨施P量的增加干物質質量呈減小趨勢,P1、P2干物質質量與P3差異顯著。與無鹽脅迫相比,輕度鹽脅迫下,P2水平的株高降幅相對最小,為13.1%,葉面積在不同P水平間降幅差異不顯著,葉綠素含量在P1、P2水平降幅相對較小,分別為13.1%、16.2%,干物質質量在P1水平降幅相對最小,為30.6%;中度、重度鹽脅迫下,株高、葉面積、葉綠素含量、干物質質量在P1水平的降幅相對最小,株高降幅分別為20.7%、34.6%,葉面積降幅分別為30.9%、45.3%,葉綠素含量降幅分別為20.5%、289%,干物質質量降幅分別為37.4%、50.5%。
相同N2P2水平下,輕度鹽脅迫處理,隨施K量的增加,谷子幼苗株高、葉面積、干物質質量呈先增加后減小的趨勢,K2株高大于K1、K3,K1、K2的葉面積、干物質質量大于K3,差異顯著;中度、重度鹽脅迫下,隨施K量的增加,谷子幼苗株高、葉面積、干物質質量呈減小趨勢,中度鹽脅迫下均為K1>K2>K3,差異顯著,重度鹽脅迫下株高為K1>K2>K3,K1葉面積、干物質質量大于K2、K3,差異顯著;隨施K量的增加,輕度、中度鹽脅迫下葉綠素含量呈先增加后減小的趨勢,重度鹽脅迫下葉綠素含量呈減小趨勢,3種鹽脅迫下,K1、K2葉綠素含量大于K3,差異顯著。與無鹽脅迫相比,輕度鹽脅迫下株高在K2水平,葉面積、葉綠素含量、干物質質量在K1水平降幅相對最小,分別為13.1%、15.6%、10.0%、33.5%;中度、重度鹽脅迫下,株高、葉面積、葉綠素含量、干物質質量在K1水平降幅相對最小,其中,株高降幅分別為24.4%、32.6%,葉面積降幅分別為29.3%、39.8%,葉綠素含量降幅分別為225%、31.0%,干物質質量降幅分別為 41.5%、58.2%。
N、P、K交互作用情況下,輕度鹽脅迫處理,株高與干物質質量在N1P2K1,葉綠素含量在N1P2K2,葉面積在N1P2K2、N1P2K1相對較大;中度鹽脅迫下,株高、葉面積、葉綠素含量在N1P2K2、N1P2K1及干物質質量在N1P2K1相對較大;重度鹽脅迫下,株高、葉面積、干物質質量在N1P2K1,葉綠素含量在N1P2K2、N1P2K1相對較大。與無鹽脅迫相比,輕度鹽脅迫下,株高在N1P2K2、N1P2K1、N1P1K2的降幅相對較小,分別為5.7%、8.0%、8.4%,中度鹽脅迫下N1P2K2、重度鹽脅迫下N1P2K1的降幅相對最小,分別為7.9%、21.8%;葉面積、葉綠素含量在不同處理間降幅差異不顯著,干物質質量在N1P2K1、N1P2K2、N1P1K2的降幅相對較小。
2.2N、P、K對鹽脅迫谷子幼苗滲透調節物質的影響
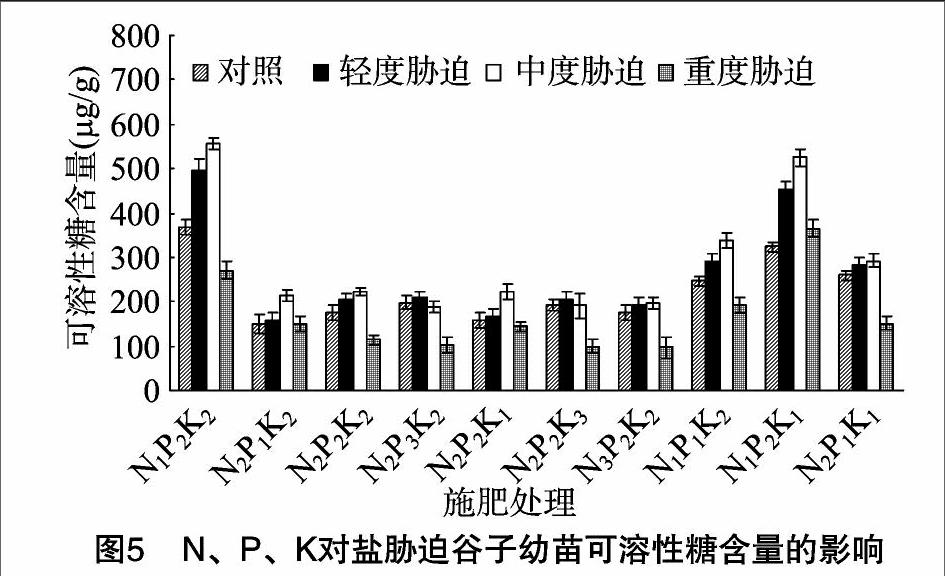
由圖5至圖7可知,相同P2K2水平下,3種鹽脅迫處理,隨施N量的增加,谷子幼苗可溶性糖、可溶性蛋白質、脯氨酸含量呈減小趨勢,輕度鹽脅迫下N1可溶性糖含量大于N2、N3,可溶性蛋白質、脯氨酸含量大小為N1>N2>N3,中度與重度鹽脅迫下,可溶性糖、可溶性蛋白質、脯氨酸含量大小為N1>N2>N3,差異顯著。與無鹽脅迫相比,輕度與中度鹽脅迫下可溶性糖、可溶性蛋白質、脯氨酸含量在N1水平增幅相對較大,其中,可溶性糖含量增幅分別為35.1%、51.3%,可溶性蛋白質含量增幅分別為22.5%、32.4%,脯氨酸含量增幅分別為45.5%、76.8%;重度鹽脅迫下,可溶性糖、可溶性蛋白質含量在N1水平降幅相對最小,分別為26.2%、13.4%,脯氨酸含量則增幅相對最大,為20.3%。
相同N2K2水平下,隨施P量增加,輕度鹽脅迫的谷子幼苗可溶性糖、脯氨酸含量呈增加趨勢,P2、P3>P1,中度鹽脅迫的谷子幼苗可溶性糖、脯氨酸含量呈先增加后減小趨勢,P1、P2>P3,重度鹽脅迫的谷子幼苗可溶性糖、脯氨酸含量呈減小趨勢,P1可溶性糖含量大于P2、P3,脯氨酸含量大小為P1>P2>P3,差異顯著;隨施P量的增加,輕度鹽脅迫的谷子幼苗可溶性蛋白質含量呈先增加后減小趨勢,P2>P3>P1,中度與重度鹽脅迫的谷子幼苗可溶性蛋白質含量呈減小趨勢,P1>
P2>P3,差異顯著。與無鹽脅迫相比,谷子幼苗可溶性糖、可溶性蛋白質、脯氨酸含量在輕度鹽脅迫下P2水平的增幅相對最大,分別為17.6%、9.3%、28.9%,中度鹽脅迫下P1水平的增幅相對最大,分別為42.4%、23.2%、46.4%;重度鹽脅迫下,可溶性糖、可溶性蛋白質含量在P1水平的降幅相對最小,分別為9.7%、10.7%,脯氨酸含量增幅相對最大,為2.8%。
相同N2P2水平下,隨施K量的增加,輕度與中度鹽脅迫處理谷子幼苗可溶性糖含量呈先增加后減小的趨勢,重度鹽脅迫下可溶性糖含量呈減小趨勢,輕度鹽脅迫下K2、K3可溶性糖含量大于K1,中度鹽脅迫下K1、K2可溶性糖含量大于K3,重度鹽脅迫下K1可溶性糖含量大于K2、K3,差異顯著。隨施K量的增加,輕度鹽脅迫下可溶性蛋白質、脯氨酸含量呈先增加后減小趨勢,K2可溶性蛋白質、脯氨酸含量大于K1、K3,中度與重度鹽脅迫下可溶性蛋白質、脯氨酸含量呈減小趨勢,K1、K2可溶性蛋白質含量大于K3,中度鹽脅迫下,K1脯氨酸含量大于K2、K3,重度鹽脅迫下脯氨酸含量大小為 K1>K2>K3,差異顯著。與無鹽脅迫相比,可溶性糖、可溶性蛋白質、脯氨酸含量在輕度鹽脅迫K2水平的增幅相對最大,分別為17.6%、9.3%、28.9%,中度鹽脅迫K1水平的增幅相對最大,分別為39.0%、47.1%、80.6%,重度鹽脅迫下,可溶性糖含量在K1水平降幅相對最小,為9.6%,可溶性蛋白質、脯氨酸含量增幅相對最大,分別為1.9%、22.3%。
N、P、K交互作用下,輕度鹽脅迫下可溶性糖與可溶性蛋白質含量在N1P2K2、脯氨酸含量在N1P2K1相對最高,中度鹽脅迫下可溶性糖含量在N1P2K2、可溶性蛋白質與脯氨酸含量在N1P2K1相對最高,重度鹽脅迫下可溶性糖、可溶性蛋白質、脯氨酸含量N1P2K1相對最高。與無鹽脅迫相比,輕度鹽脅迫下,可溶性糖含量在N1P2K1、N1P2K2的增幅相對最大,分別為39.6%、35.1%,可溶性蛋白質含量在N1P2K2的增幅相對最大,為22.5%,脯氨酸含量在N1P2K1、N1P2K2、N1P1K2的增幅相對較大,分別為53.9%、45.5%、39.6%;中度與重度鹽脅迫下,可溶性糖、可溶性蛋白質、脯氨酸含量在N1P2K1的增幅相對最大,中度鹽脅迫下的增幅分別為62.2%、59.3%、93.9%,重度鹽脅迫下的增幅分別為12.8%、11.3%、38.7%。
2.3N、P、K對鹽脅迫谷子幼苗保護酶活性的影響
由圖8至圖10可知,相同P2K2水平下,隨施N量的增加,3種鹽脅迫處理谷子幼苗的POD、SOD、CAT酶活性呈減小趨勢,N1>N2>N3,差異顯著。與無鹽脅迫相比,POD、SOD、CAT酶活性在輕度與中度鹽脅迫下N1水平增幅相對最大,重度鹽脅迫下降幅相對最小,其中,輕度鹽脅迫下的增幅分別為49.9%、31.6%、86.8%,中度鹽脅迫下的增幅分別為74.6%、48.2%、111.1%;重度鹽脅迫下的降幅分別為12.2%、11.2%、9.7%。
相同N2K2水平下,隨施P量的增加,輕度與中度鹽脅迫谷子幼苗的POD、SOD、CAT酶活性呈先增加后減小的趨勢,重度鹽脅迫下POD、SOD酶活性顯著減小;輕度鹽脅迫下,P2的POD、SOD酶活性大于P1、P3,P2、P3的CAT酶活性大于P1,中度鹽脅迫下,P1、P2的POD、SOD、CAT酶活性大于P3,重度鹽脅迫下POD、SOD酶活性大小為P1>P2>P3,差異顯著,CAT酶活性在不同P水平間差異不顯著。與無鹽脅迫相比,輕度鹽脅迫下POD酶活性在P1水平增幅相對最大,為19.2%,SOD、CAT酶活性在P2水平增幅相對最大,分別為11.7%、65.1%;中度鹽脅迫下,POD、SOD酶活性在P1水平增幅相對最大,分別為25.6%、26.2%,CAT酶活性在P1、P2水平增幅相對較大,分別為78.5%、74.0%;重度鹽脅迫下,POD、SOD、CAT酶活性在P1水平降幅相對最小,分別為 16.9%、15.5%、17.7%。
相同N2P2水平下,隨施K量增加,輕度鹽脅迫谷子幼苗的POD酶活性呈先增加后減小趨勢,中度與重度鹽脅迫下POD酶活性呈減小趨勢;輕度鹽脅迫下K2的POD酶活性大于K1、K3,中度鹽脅迫下K1的POD酶活性大于K2、K3,重度鹽脅迫下的POD酶活性為K1>K2>K3,差異顯著。隨施K量增加,輕度與中度鹽脅迫下SOD酶活性呈增加趨勢,重度鹽脅迫下SOD酶活性呈減小趨勢;輕度鹽脅迫下SOD酶活性為K3>K2>K1,中度鹽脅迫下K2、K3的SOD酶活性大于K1,重度鹽脅迫下K1、K2的SOD酶活性大于K3,差異顯著。隨施K量的增加,輕度與中度鹽脅迫下CAT酶活性呈先增加后減小趨勢,重度鹽脅迫下呈減小趨勢;輕度鹽脅迫下,K2、K3的CAT酶活性大于K1,中度鹽脅迫下K1、K2的CAT酶活性大于K3,重度鹽脅迫下K1的CAT酶活性大于K2、K3,差異顯著。與無鹽脅迫相比,輕度鹽脅迫下,POD酶活性在K1水平增幅相對最大,為18.6%,SOD、CAT酶活性在K2水平增幅相對最大,分別為11.7%、65.1%;中度鹽脅迫下,POD、SOD酶活性在K1水平增幅相對最大,分別為35.2%、270%,CAT酶活性在K1、K2水平增幅相對較大,分別為651%、74.0%;重度鹽脅迫下,POD、SOD、CAT酶活性在K1水平降幅相對最小,分別為17.9%、17.1%、9.0%。
N、P、K交互作用情況下,輕度鹽脅迫處理,POD酶活性在N1P2K2、N1P2K1及SOD酶活性在N1P2K2、CAT酶活性在N1P2K1相對較高;中度與重度鹽脅迫下,POD與CAT酶活性在N1P2K1、SOD酶活性在N1P2K2相對最高。與無鹽脅迫相比,輕度鹽脅迫下谷子幼苗的POD、SOD酶活性在N1P2K2、N1P2K1、N1P1K2的增幅相對較大,分別為49.9%、43.9%、35.2%和31.6%、27.2%、23.0%,CAT酶活性在N1P2K1、N1P2K2的增幅相對較大,分別為91.8%、86.8%;中度鹽脅迫下,POD、SOD酶活性在N1P2K1的增幅相對最大,分別為 90.9%、66.8%,CAT酶活性在N1P2K1、N1P2K2的增幅相對較大,分別為111.5%、111.1%;重度鹽脅迫下,POD、SOD酶活性在N1P2K1的降幅相對最小,分別為4.4%、1.8%,CAT酶活性增幅相對最大,為14.4%。
2.4N、P、K對鹽脅迫谷子幼苗MDA含量的影響
由圖11可知,相同P2K2水平下,隨施N量的增加,3種鹽脅迫處理MDA含量呈增加趨勢,N3>N2>N1,差異顯著。與無鹽脅迫相比,輕度、中度與重度鹽脅迫下MDA含量在N1水平增幅相對最小,分別為11.6%、55.%、85.8%。
相同N2K2水平下,隨施P量的增加,輕度鹽脅迫下MDA含量呈先減小后增加趨勢,中度與重度鹽脅迫下MDA含量
呈增加趨勢;輕度與中度鹽脅迫下P3的MDA含量大于P1、P2,重度鹽脅迫下MDA含量大小為P3>P2>P1,差異顯著。與無鹽脅迫相比,輕度、中度與重度鹽脅迫下MDA含量在P1水平的增幅相對最小,分別為47.7%、60.8%、86.3%。
相同N2P2水平下,隨施K量的增加,3種鹽脅迫處理MDA含量呈增加趨勢;輕度鹽脅迫下,K3的MDA含量大于K1、K2,中度與重度鹽脅迫下MDA含量為K3>K2>K1,差異顯著。與無鹽脅迫相比,輕度、中度與重度鹽脅迫下MDA含量在K1水平增幅相對最小,分別為49.4%、63.0%、88.8%。
N、P、K交互作用情況下,輕度鹽脅迫處理的谷子幼苗MDA含量在N1P2K2、N1P2K1水平相對較小,中度與重度鹽脅迫下N1P2K1水平相對最小。與無鹽脅迫相比,輕度鹽脅迫下MDA含量在N1P2K1、N1P2K2的增幅相對較小,分別為 10.1%、11.6%,中度與重度鹽脅迫下N1P2K1的增幅相對最小,分別為14.8%、32.7%。
3討論與結論
鹽害是農業生產上重要的逆境危害之一。生長指標和生物量反映逆境脅迫下植物的適應性和逆境脅迫效應[19],滲透調節是植物適應鹽脅迫的最基本特征之一[20],抗氧化酶活性的提高是活性氧自由基增加和脅迫條件下保護酶系統建立的標志[21],植物器官在逆境條件或衰老時,丙二醛(MDA)含量用于表示細胞膜質過氧化程度和植物對逆境條件反應的強弱。本試驗結果表明,輕度鹽脅迫下,谷子幼苗在N1、P2、K2水平,即N、P、K含量分別為100、87、249 mg/kg時各指標相對較好,此時株高的降幅較小,滲透調節物質含量、SOD、CAT酶活性增幅較大;中度鹽脅迫下,N1、P1、P2、K1、K2水平,即N含量為100 mg/kg,P含量為43.5、87 mg/kg,K含量為124.5、249 mg/kg時各指標相對較好,各形態指標及干物質質量的降幅相對較小,滲透調節物質含量和POD、SOD酶活性增幅相對大;重度鹽脅迫下,N1、P1、K1水平,即N、P、K含量分別為100、43.5、124.5 mg/kg 時各指標較好,各形態指標及干物質質量、保護酶活性的降幅相對較小,N1、P1水平可溶性糖和可溶性蛋白含量降幅相對較小,脯氨酸含量增幅相對較大,K1水平可溶性糖含量降幅相對最小,可溶性蛋白質、脯氨酸含量增幅相對最大;3種鹽脅迫下,N1、P1、K1水平MDA含量增幅相對最小。這表明谷子幼苗在輕度鹽脅迫下N1、P2、K2水平、中度鹽脅迫下N1、P1、P2、K1、K2水平及重度鹽脅迫下N1、P1、K1水平抗鹽效果較好,適量施肥對提高谷子體內滲透調節物質含量、保護酶活性有好處,有利于抵抗鹽脅迫,較高的N、P、K增加了總鹽分濃度,鹽害加重,對谷子生長不利。N、P、K交互作用下,輕度鹽脅迫谷子幼苗的形態指標及干物質質量在N1P2K2、N1P2K1、N1P1K2相對較好,N1P2K2有利于滲透調節物質、保護酶抗鹽性的發揮,N1P2K2、N1P2K1有利于減輕膜脂過氧輕度化作用;中度與重度鹽脅迫下,形態指標及干物質質量在N1P2K1相對最好,有利于滲透調節物質、保護酶抗鹽性的發揮及減輕膜脂過氧化作用。這說明,鹽脅迫下適當多施用P、K可以緩解鹽脅迫。
逄煥成等認為,NaCl脅迫下分別施N 150、300、450 mg/kg(土),能顯著增加小麥株高、葉面積、干物質積累量[12]。寧建鳳試驗發現,海鹽濃度8.3、16.7 g/L條件下施N 200 mg/kg,可使2種海鹽處理的蘆薈葉片可溶性糖、脯氨酸含量增幅最大,有利于增強蘆薈植株的滲透調節能力,降低蘆薈葉片中MDA含量[22]。沈振國等研究認為,Nacl 0.85、1.25 g/kg(土)脅迫下,大麥葉片的SOD酶活性下降,施N 100、200 mg/kg (土)能增加葉片的SOD酶活性,特別是在NaCl濃度較高時,N素的效應更為明顯[23]。逄煥成等研究得出的最佳N施肥量[12,22-23]高于本試驗結果,可能是由于本試驗采用沙培,沒有土壤吸附離子造成的緩解作用。因此,鹽地谷子苗期施肥時,可根據耕地土壤對肥料的吸附特性適當增加N、P、K用量。
王玉鳳研究表明,含150 mmol/L NaCl、1 mmol/L總磷的1/2 Hongland 營養液培養玉米幼苗的干質量、葉綠素、可溶性糖、可溶性蛋白質含量和POD、SOD、CAT酶活性等明顯增加,MDA含量降低[6]。楊曉英等認為,2、3 g/kg NaCl脅迫及施N 100 mg/kg(土)條件下,施P2O5 100 mg/kg (土)能明顯增加黑麥草幼苗干物質積累、葉綠素、可溶性糖、可溶性蛋白質、脯氨酸含量,促進鹽脅迫下黑麥草幼苗的生長[7]。鄭延海等研究發現,1.58 g/kg (沙) NaCl脅迫下,KNO3濃度為 10 mmol/L時可以使小麥幼苗株高、葉面積受到的鹽分傷害得到緩解,POD、SOD、CAT酶活性升高、MDA含量降低,過高的KNO3濃度(15 mmol/L)對小麥生長不利[11]。本試驗結果與以上結論均相似。
參考文獻:
[1]楊立,王博祺,韓鋒. 改革開放以來我國耕地保護績效定量研究——基于數量保護的視角[J]. 農機化研究,2015(3):1-6.
[2]趙可夫,李法曾. 中國鹽生植物[M]. 北京:科學出版社,1999:1-6.
[3]崔興國,時麗冉. 衡水地區14份夏谷品種種子萌發期耐鹽性研究[J]. 作物雜志,2011(4):117-119.
[4]田伯紅,王建廣,李雅靜,等. 谷子品種芽期和苗期對NaCl脅迫的反應及耐鹽性鑒定[C]. 2008中國作物學會學術年會論文摘要集,2008:123.
[5]張瑞玖,蒙美蓮,酈海龍,等. NaCl脅迫下氮對馬鈴薯的調控作用[J]. 中國農學通報,2010,26(6):146-149.
[6]王玉鳳. 玉米苗期對NaCl脅迫的響應與耐鹽性調控機理的研究[D]. 沈陽:沈陽農業大學,2008.
[7]楊曉英,楊勁松. 鹽脅迫對黑麥草幼苗生長的影響及磷肥的緩解作用[J]. 土壤通報,2005(6):85-88.
[8]黃增榮,隆小華,劉兆普,等. KNO3對NaCl脅迫下兩菊芋品種幼苗生長及光合能力的影響[J]. 草業學報,2011,20(1):82-88.[9]王朝霞,蒙美蓮,張靜,等. NaCl脅迫下鉀對馬鈴薯的調控作用[J]. 作物雜志,2012(4):94-97.
[10]王玉鳳,薛盈文,楊克軍,等. 鉀對NaCl脅迫下玉米幼苗保護酶活性等生理特性的影響[J]. 黑龍江八一農墾大學學報,2010,22(4):19-22.
[11]鄭延海,寧堂原,賈愛君,等. 鉀營養對不同基因型小麥幼苗NaCl脅迫的緩解作用[J]. 植物營養與肥料學報,2007,13(3):381-386.
[12]逄煥成,楊勁松,謝曉紅. 氯化鈉脅迫下施氮對冬小麥生長發育及體內氯、鈉離子積累的影響[J]. 植物營養與肥料學報,2005,11(5):654-658,664.
[13]穆靜,劉小京,徐進,等. 氮素對NaCl脅迫下甜高粱種子萌發及芽苗生長與生理的影響[J]. 中國生態農業學報,2012,20(10):1303-1309.
[14]智慧,刁現民,呂芃,等. 人工鹽脅迫法鑒定谷子及狗尾草物種耐鹽基因型[J]. 河北農業科學,2004,8(4):15-18.
[15]田伯紅. 鹽堿脅迫對谷子農藝性狀及產量的影響[J]. 河北農業科學,2009,13(10):4-5.
[16]李合生. 植物生理生化實驗原理和技術[M]. 北京:高等教育出版社,2000:258-260.
[17]王學奎. 植物生理生化實驗原理和技術[M]. 北京:高等教育出版社,2006:172-173.
[18]張志良,瞿偉菁. 植物生理學實驗指導[M]. 北京:高等教育出版社,2003:274-276.
[19]宋云鵬. 外源硅對NO-3脅迫下黃瓜幼苗生長及生理生化特性的影響[D]. 泰安:山東農業大學,2011:37.
[20]劉友良,汪良駒. 植物對鹽脅迫的反應和耐鹽性[M]//余叔文,湯章城.植物生理與分子生物學. 2版.北京:科學出版社,1998:752-798.
[21]Meloni D A,Oliva M A,Martinez C A,et al. Photosynthesis and activity of superoxide dismutase,peroxidase and glutathione reductase in cotton under salt stress[J]. Environmental and Experimental Botany,2003,49(1):69-76.
[22]寧建鳳. 氮對鹽脅迫下庫拉索蘆薈生長及生理特性的影響[D]. 南京:南京農業大學,2005:46.
[23]沈振國,沈其榮,管紅英,等. NaCl脅迫下氮素營養與大麥幼苗生長和離子平衡的關系[J]. 南京農業大學學報,1994,17(1):22-26.聶俊,邱俊榮,史亮亮,等. 有機肥和化肥配施對拋栽水稻產量、品質及鉀吸收轉運的影響[J]. 江蘇農業科學,2016,44(2):122-125.
