凋落物和林下植被對杉木林土壤碳氮水解酶活性的影響機制
2016-04-11 08:22:55王繼富張心昱李丹丹王輝民陳伏生孫曉敏溫學發(fā)
生態(tài)學報 2016年24期
楊 洋,王繼富,張心昱,李丹丹,王輝民,陳伏生,孫曉敏,溫學發(fā)
1 哈爾濱師范大學地理科學學院,哈爾濱 150025 2 中國科學院地理科學與資源研究所生態(tài)系統(tǒng)網(wǎng)絡觀測與模擬重點實驗室,北京 100101 3 沈陽農(nóng)業(yè)大學生物科學技術學院,沈陽 110866 4 江西農(nóng)業(yè)大學江西省竹子種質(zhì)資源與利用重點實驗室,南昌 330045
凋落物和林下植被對杉木林土壤碳氮水解酶活性的影響機制
楊 洋1,2,王繼富1,張心昱2, *,李丹丹2, 3,王輝民2,陳伏生4,孫曉敏2,溫學發(fā)2
1 哈爾濱師范大學地理科學學院,哈爾濱 150025 2 中國科學院地理科學與資源研究所生態(tài)系統(tǒng)網(wǎng)絡觀測與模擬重點實驗室,北京 100101 3 沈陽農(nóng)業(yè)大學生物科學技術學院,沈陽 110866 4 江西農(nóng)業(yè)大學江西省竹子種質(zhì)資源與利用重點實驗室,南昌 330045

凋落物;林下植被;杉木林;土壤環(huán)境因子;β- 1,4-葡萄糖苷酶;β- 1,4-N-乙酰葡糖氨糖苷酶
土壤微生物常被比作土壤有機質(zhì)(SOM)中碳、氮等養(yǎng)分元素循環(huán)的“轉(zhuǎn)化器”,是控制SOM周轉(zhuǎn)的重要因子[1]。微生物不同酶活性可以反映微生物對土壤中不同底物的分解強度[2],因而被作為表征SOM中碳、氮分解的敏感指標[3- 4]。
森林凋落物和林下植被是森林生態(tài)系統(tǒng)的重要組成部分。凋落物作為森林SOM的主要來源[5],為土壤微生物提供反應底物和棲息地[6]。凋落物通過微生物分解將植物吸收利用的養(yǎng)分重新釋放到環(huán)境中,參與土壤碳氮及其他養(yǎng)分的循環(huán)過程,在森林生態(tài)系統(tǒng)中起重要作用[7- 8]。林下植被的種類和數(shù)量會影響土壤根際分泌物的數(shù)量和多樣性[9],從而改變SOM含量,在維持生物多樣性和土壤小氣候、生態(tài)系統(tǒng)養(yǎng)分循環(huán)等方面扮演重要角色[10]。森林凋落物和林下植被通過改變土壤酶活性影響土壤內(nèi)部的物質(zhì)與能量循環(huán)[2]。改變地表凋落物和林下植被的數(shù)量將會引起土壤質(zhì)量以及微生物活性的變化。
目前,國內(nèi)關于凋落物對土壤酶活性影響的研究主要以改變凋落物種類或凋落物組成為主[5],而林下植被對森林土壤的影響主要集中于寒帶和溫帶森林[11]。已有研究發(fā)現(xiàn),添加凋落物會增強[12]或不影響[13- 15]土壤酶活性,大部分去除凋落物處理降低了土壤酶活性[12- 15]。關于林下植被對土壤酶活性的研究較少,在亞熱帶高山針葉林中去除林下植被導致微生物數(shù)量下降因而降低土壤酶活性[16],在溫帶樟子松人工林去除林下植被對酶活性沒有影響[9]。出現(xiàn)這種不一致性的原因可能是氣候條件、森林和土壤類型、樣地大小、處理時間長短等因素的差異[6, 8]。有研究指出,土壤水解酶活性與土壤溫度(ST)、土壤含水量(SWC)、土壤酸度(pH)以及土壤碳和養(yǎng)分含量有關,但是相關程度因不同生態(tài)系統(tǒng)而異[17]。凋落物和林下植被處理引起土壤物理、化學、生物性質(zhì)變化,從而導致酶活性的改變[12]。凋落物和林下植被管理措施對杉木林土壤碳氮水解酶活性影響機制還不清楚。
人類活動對森林凋落物和林下植被的管理有明顯影響。過去,在發(fā)展中國家,凋落物和林下植被常被作為燃料移出森林生態(tài)系統(tǒng)[18- 19]。目前,為減少林下植被與森林競爭養(yǎng)分、便于幼苗萌發(fā)、防止火災等,清理凋落物和林下植被成為很多國家森林生態(tài)系統(tǒng)管理的重要措施[6,20]。杉木(Cunninghamialanceolata)是我國亞熱帶種植最廣泛的速生豐產(chǎn)樹種,占地面積為9.11×106hm2,占我國人工林面積的18%[21]。然而營造杉木純林和重復連栽會造成林地土壤質(zhì)量下降[22]。清理凋落物和割除林下植被的經(jīng)營措施也可能加劇杉木林土壤質(zhì)量退化。本研究選取杉木二代人工林,通過添加和去除凋落物、去除林下植被處理,研究凋落物和林下植被對杉木林土壤環(huán)境因子和酶活性影響,揭示凋落物和林下植被對杉木林土壤碳氮水解酶活性影響機制。由于凋落物和林下植被添加增加了土壤水解酶反應底物含量,假設添加凋落物和保留林下植被有利于增強土壤碳氮水解酶活性,其影響機制主要是由于底物含量的增加。
1 材料與方法
1.1 試驗設計
試驗樣地位于江西省泰和縣中國科學院千煙洲石溪林場(115°03′29.9″E,26°44′29.1″N)。該地屬于紅壤丘陵區(qū),亞熱帶季風氣候,年平均氣溫約為17.8 ℃,年降水量為1471.2 mm。
2013年1月在杉木林樣地中設置6個30 m×30 m樣地,隨機排列。每個樣地中設置4個15 m×15 m樣方,總計24個樣方。為了研究凋落物、林下植被的作用,采用生態(tài)學上人為擴大處理量的方法,其中凋落物采用加倍處理,以期在較短時間內(nèi)開展生態(tài)學尺度上的機理研究。設置4個處理:去除凋落物+去除林下植被(LR+UR)、去除凋落物+保留林下植被(LR+U)、凋落物加倍+保留林下植被(LD+U)、凋落物加倍+割倒林下植被并歸還原樣地(LD+UC)(圖1)。每個處理6個重復。在試驗設立初期,對杉木林枯枝進行了清理,此后每個月對樣地進行維護:(1)LR+UR:清除樣方內(nèi)的凋落物,將樣方內(nèi)的林下植被割倒并移除樣方。(2)LR+U:僅清除樣方內(nèi)的凋落物,樣方內(nèi)的林下植被不做處理。(3)LD+U:將從去除凋落物樣地收集的凋落物均勻灑在樣方內(nèi),林下植被不做處理。(4)LD+UC:將從去除凋落物樣地收集的凋落物均勻灑在樣方內(nèi),將樣方內(nèi)的林下植被割倒后均勻灑在樣方內(nèi)。由于杉木林凋落物具有枝、葉同時凋落的特征,在處理期間每月凋落物量較少,每月1次維護可以保證凋落物處理效果。在每月維護中,重點是在相關樣地中進行草本、灌木的相關處理。杉木林凋落物量大約為1020 kg hm-2a-1,林下植被量大約為6236 kg/hm2。其中,通過比較LR+U和LD+U處理研究凋落物對土壤的影響;通過比較LR+UR和LR+U處理研究林下植被對土壤的影響;通過比較LR+UR和LD+U、LD+UC研究凋落物和林下植被的共同作用對土壤的影響,其中通過比較LD+U和LD+UC研究林下植被正常生長與林下植被割倒歸還對土壤影響。

圖1 不同凋落物和林下植被處理Fig.1 Different litter and understory treatments 去除凋落物+去除林下植被(LR+UR)、去除凋落物+保留林下植被(LR+U)、凋落物加倍+保留林下植被(LD+U)、凋落物加倍+割倒林下植被歸還原樣地(LD+UC)
1.2 樣品采集與分析

土壤酶活性分析采用微孔板熒光法[4]。β- 1,4-葡萄糖苷酶(βG)活性的測定以4-甲基傘形酮酰-β-D-吡喃葡萄糖苷(4-MUB-β-D-glucoside)為底物,β- 1,4-N-乙酰葡糖氨糖苷酶(NAG)以4-甲基傘形酮酰-乙酰基-β-D-氨基葡萄糖苷(4-MUB-N-acetyl-β-D-glucosaminide)為底物,使用多功能酶標儀(Synergy H4, BioTek)測定。
1.3 數(shù)據(jù)處理
數(shù)據(jù)為平均值±標準誤(n=6)。采用雙因素方差分析和Duncan多重比較,進行不同處理和采樣時間對土壤環(huán)境因子、酶活性影響的顯著性檢驗,顯著水平P<0.05;利用主成分分析(PCA)評價各處理對土壤環(huán)境因子和碳氮水解酶活性的影響,用SPSS 17.0(Chicago,IL, USA)分析。利用Canoco 4.5軟件冗余分析(RDA)評價酶活性與土壤環(huán)境因子的相關性。利用SigmaPlot 10.0軟件進行統(tǒng)計繪圖。
2 結(jié)果分析
2.1 凋落物和林下植被對土壤環(huán)境因子影響
各處理對ST沒有顯著性影響(表1)。除了LD+UC處理比LR+UR顯著增加了10%(P<0.05)外,其他處理SWC之間沒有顯著性差異(表2)。LD+U和LD+UC處理的土壤pH分別比LR+UR顯著增加了0.15和0.16(P<0.05)。ST、SWC和pH都存在明顯的季節(jié)變化(P<0.01)(表1),ST為:7月>4月>11月,SWC和pH:11月>7月>4月(表3)。
表1 凋落物和林下植被處理和采樣時間對土壤環(huán)境因子和碳氮水解酶活性影響的雙因素方差分析
Table 1 Two-way ANOVA of litter and understory treatments and sampling times on the soil environmental factors and the carbon and nitrogen hydrolase activities

因素FactorsSTSWCpHNO-3-NNH+4-NDOCβGNAG處理Treatment0.410.070.04?0.820.090.00??0.00??0.00??采樣時間Samplingtime0.00??0.00??0.00??0.00??0.00?0.00??0.00??0.00??

2.2 凋落物和林下植被對土壤碳氮水解酶活性的影響
各處理對土壤βG、NAG酶活性影響極顯著(表1)。βG、NAG活性都表現(xiàn)為LD+UC>LD+U>LR+U>LR+UR,LR+U、LD+U和LD+UC處理的βG活性分別比LR+UR增強了46%、63%和85%(P<0.05),NAG活性比LR+UR增強了26%、43%和106%(P<0.05)。βG和NAG活性都具有明顯的季節(jié)動態(tài)(表1),均表現(xiàn)為11月>7月>4月(P<0.05)(圖2)。

表2 凋落物和林下植被處理對土壤環(huán)境因子的影響
去除凋落物+去除林下植被litter and understory vegetation removal(LR+UR);去除凋落物+保留林下植被litter removal but understory vegetation intact(LR+U);凋落物加倍+保留林下植被litter doubled and understory vegetation intact(LD+U);凋落物加倍+割倒林下植被歸還原樣地litter doubled and understory vegetation clipped but leftinsitu(LD+UC);表中數(shù)值為平均值±標準誤,每列不同小寫字母表示不同處理之間差異顯著(P<0.05)
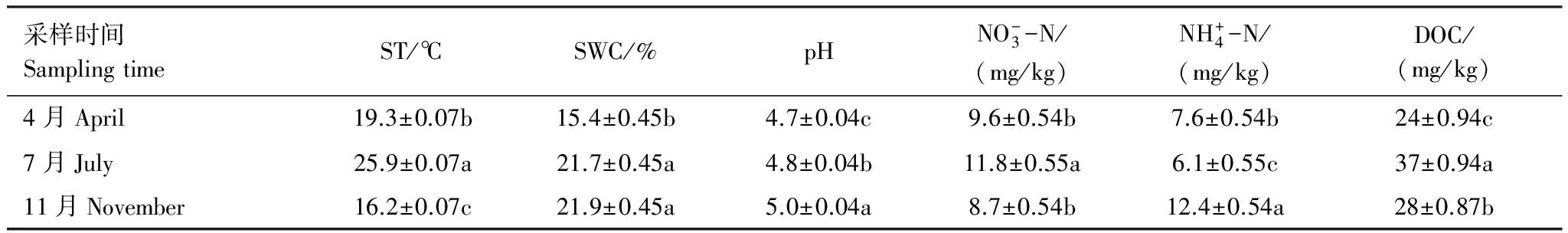
表3 采樣時間對土壤環(huán)境因子的影響

圖2 凋落物和林下植被處理及采樣時間對土壤β- 1,4-葡萄糖苷酶(βG)和β- 1,4-N-乙酰葡糖氨糖苷酶(NAG)活性的影響Fig.2 Effects of litter and understory treatments and sampling times on soil β- 1, 4-glucosidase (βG) activity and soil β- 1, 4-N-acetyl-glucosaminidase (NAG) activity
2.3 主成分分析(PCA)


圖3 土壤碳氮水解酶與土壤環(huán)境因子的主成分分析Fig.3 Principal component analysis of soil carbon and nitrogen hydrolase activities and soil environmental factorsa. 不同凋落物和林下植被處理的得分;誤差棒代表平均值±標準誤;b. 各土壤碳氮水解酶與土壤環(huán)境因子的載荷值
2.4 冗余分析(RDA)
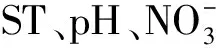

圖4 土壤碳氮水解酶與土壤環(huán)境因子冗余分析 Fig.4 Redundancy analysis of the soil carbon and nitrogen hydrolase activities and soil environmental factors
3 討論
土壤酶與土壤環(huán)境之間是相互作用的。土壤酶是土壤分解過程的主要驅(qū)動者,非生物因素通過改變微生物活動或者群落結(jié)構間接影響土壤酶活性[17],而土壤碳周轉(zhuǎn)和養(yǎng)分循環(huán)又依賴于土壤酶[12]。地表凋落物和林下植被的數(shù)量變化會通過改變養(yǎng)分有效性和微生物種群組成,調(diào)節(jié)土壤酶活性及其分配模式[12]。
與假設一致,本研究中凋落物添加有增加土壤βG、NAG活性的趨勢,但是未達到顯著水平。βG、NAG活性與土壤SWC、DOC含量相關。凋落物處理對SWC、DOC含量影響不明顯,因而沒有顯著改變碳氮水解酶活性。這可能與野外杉木林樣地土壤異質(zhì)性較高有關。細根周轉(zhuǎn)和根際分泌物為土壤提供不穩(wěn)定的有機化合物,與樹根相連的菌根也會產(chǎn)生胞外酶[14],林下植被去除后直接引起植物生物量損失和不穩(wěn)定性碳輸入減少[24]。βG在纖維素降解過程中發(fā)揮重要作用[13],參與土壤碳循環(huán)。本研究中,去除林下植被降低了土壤βG活性,可能和處理中DOC含量下降有關。已有研究通過切根試驗發(fā)現(xiàn)根際分泌物有增加SOM和提高酶活性的作用[5,24- 25]。本研究發(fā)現(xiàn)凋落物和林下植被的共同作用對土壤環(huán)境因子和水解酶活性影響顯著,并且林下植被割倒歸還比林下植被正常生長更有利于土壤NAG、βG活性的增加。導致這種現(xiàn)象的原因可能是林下植被割倒后,植物對土壤水分和養(yǎng)分的吸收減少;另一方面地上和地下部分林下植被殘體同時分解提供的碳源大于根系輸入。林下植被割倒歸還處理中較高的SWC、DOC含量與NAG、βG活性最強相一致,表明凋落物和林下植被對土壤碳氮水解酶活性的影響機制是通過提高SWC、DOC來增強土壤NAG、βG活性。有研究指出,林下植被的存在有利于凋落物的分解,從而增加了凋落物對土壤肥力和養(yǎng)分循環(huán)的影響[20]。但是本研究發(fā)現(xiàn)林下植被對土壤速效養(yǎng)分和水解酶的影響大于凋落物,表明土壤微生物活性對根系活性具有高度依賴性[14]。Schmidt等[1]認為土壤有機碳多來自植物根系,地上凋落物輸入對土壤碳儲量的影響有限。Wang等[26]也發(fā)現(xiàn)根系碳輸入對土壤微生物種群結(jié)構的影響大于凋落物輸入。這可能從側(cè)面反映了土壤碳和養(yǎng)分與微生物活性之間的內(nèi)在關系。
土壤環(huán)境因子會影響土壤胞外酶活性,這與我們的假設不完全一致。本研究中SWC與酶活性呈正相關,表明凋落物和林下植被通過影響土壤水分,進而影響βG、NAG活性。Rutigliano等[27]也發(fā)現(xiàn)微生物活性主要受到SWC變化的影響。這是因為SWC會影響土壤底物和酶的擴散速率[17]。與Kotroczo等[14]的結(jié)果一致,本研究未發(fā)現(xiàn)土壤酶活性與ST之間的相互關系。Kivlin等[17]指出在區(qū)域尺度范圍內(nèi),SWC對酶活性的影響要大于ST的影響。亞熱帶森林碳循環(huán)快,土壤可溶性碳和養(yǎng)分在凋落物分解初期從有機層向土壤快速傳輸,因而DOC被認為與酶活性具有顯著相關性[12],這與本研究結(jié)果一致,反映出凋落物和保留林下植被增強土壤碳氮水解酶活性的影響機制與底物DOC含量的增加有關。有研究表明,作為衡量酶活性的重要指標,土壤有機碳與酶活性具有相關性[28]。Hudson[29]發(fā)現(xiàn)土壤碳含量會改變酶活性,是因為碳含量增加提高了SWC。土壤DOC與水解酶活性呈正相關,可能是由于碳輸入增加通過激發(fā)效應促進微生物對SOM的分解,增加DOC含量[30],反過來DOC含量增加為土壤微生物提供更多可利用的底物,導致酶活性增強[17]。Sinsabaugh等[28]在全球尺度對來自40個生態(tài)系統(tǒng)的土壤酶活性數(shù)據(jù)進行meta分析,發(fā)現(xiàn)所測定的酶活性都與土壤pH值相關。大多數(shù)水解酶活性最適pH≈5[17]。雖然在本研究中各處理之間土壤pH出現(xiàn)了顯著性差異,但是酶活性和土壤pH值沒有相關性,這可能是因為土壤pH處于水解酶的最適pH范圍之內(nèi),且變化范圍較小,因而沒有顯著影響酶活性。
本研究中,凋落物、林下植被處理樣地βG和NAG活性均有明顯的季節(jié)動態(tài),這可能受試驗區(qū)降水和土壤pH的季節(jié)性變化影響。土壤酶活性雖然與土壤pH之間沒有相關性,但是兩者具有相同的季節(jié)變化趨勢。有研究發(fā)現(xiàn)胞外酶活性的季節(jié)性變化與碳和養(yǎng)分的有效性一致,植物也可能刺激生長季胞外酶的產(chǎn)生[17]。此外,凋落物輸入量的季節(jié)變化也會導致酶活性的季節(jié)變動[14]。
“保留凋落物、保留林下植被”還是“去除凋落物、去除林下植被”,是目前森林管理上兩種截然不同的選擇。從我們初步研究結(jié)果可以看出,凋落物、林下植被對杉木林土壤環(huán)境因子、土壤碳氮水解酶活性都有很好的改善作用。因此,保留凋落物、保留林下植被是接近林業(yè)實際的正確選擇。本研究為生態(tài)學機理研究,可以為林業(yè)生產(chǎn)實踐提供部分建議,但不完全等同于生產(chǎn)實踐處理。為了進一步證明在林業(yè)生產(chǎn)實踐管理的作用,本樣地在從2015年起補充了“保留凋落物,保留林下植被“處理,為未來凋落物、林下植被在杉木林管理研究方面提供野外平臺。
4 結(jié)論

致謝:感謝中國科學院千煙洲生態(tài)試驗站對本研究野外工作的支持和幫助。
[1] Schmidt M W I, Torn M S, Abiven S, Dittmar T, Guggenberger G, Janssens I A, Kleber M, K?gel-Knabner I, Lehmann J, Manning D A, Nannipieri P, Rasse D P, Weiner S, Trumbore S E. Persistence of soil organic matter as an ecosystem property. Nature, 2011, 478(7367): 49- 56.
[2] Cusack D F, Silver W L, Torn M S, Burton S D, Firestone M K. Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests. Ecology, 2011, 92(3): 621- 632.
[3] Burns R G, DeForest J L, Marxsen J, Sinsabaughd R L, Strombergere M E, Wallensteinf M D, Weintraubg M N, Zoppini A. Soil enzymes in a changing environment: current knowledge and future directions. Soil Biology and Biochemistry, 2013, 58: 216- 234.
[4] Saiya-Cork K R, Sinsabaugh R L, Zak D R. The effects of long term nitrogen deposition on extracellular enzyme activity in anAcersaccharumforest soil. Soil Biology and Biochemistry, 2002, 34(9), 1309- 1315.
[5] 劉星, 王娜, 趙博, 張青, 趙秀海. 改變碳輸入對太岳山油松林土壤酶活性的影響. 應用與環(huán)境生物學報, 2014, 20(4): 655- 661.
[6] Sayer E J. Using experimental manipulation to assess the roles of leaf litter in the functioning of forest ecosystems. Biological Reviews, 2006, 81(1): 1- 31.
[7] Yan W D, Chen X Y, Tian D L, Peng Y Y, Wang G J, Zheng W. Impacts of changed litter inputs on soil CO2efflux in three forest types in central south China. Chinese Science Bulletin, 2013, 58(7): 750- 757.
[8] Vitousek P M, Sanford J L Jr. Nutrient cycling in moist tropical forest. Annual Review of Ecology and Systematics, 1986, 17(1): 137- 167.
[9] 林貴剛, 趙瓊, 趙蕾, 李慧超, 曾德慧. 林下植被去除與氮添加對樟子松人工林土壤化學性質(zhì)和生物學性質(zhì)的影響. 應用生態(tài)學報, 2012, 23(5): 1188- 1194.
[10] Zhao J, Wan S Z, Fu S L, Wang X L, Wang M, Liang C F, Chen Y Q, Zhu X L. Effects of understory removal and nitrogen fertilization on soil microbial communities inEucalyptusplantations. Forest Ecology and Management, 2013, 310: 80- 86.
[11] Wang F M, Zou B, Li H F, Li Z A. The effect of understory removal on microclimate and soil properties in two subtropical lumber plantations. Journal of Forest Research, 2014, 19(1): 238- 243.
[12] Weintraub S R, Wieder W R, Cleveland C C, Townsend A R. Organic matter inputs shift soil enzyme activity and allocation patterns in a wet tropical forest. Biogeochemistry, 2013, 114(1/3): 313- 326.
[13] Veres Z, Kotroczó Z, Fekete I, Tóth J A, Lajtha K, Townsend K, Tóthmérész B. Soil extracellular enzyme activities are sensitive indicators of detrital inputs and carbon availability. Applied Soil Ecology, 2015, 92: 18- 23.
[14] Kotroczó Z, Veres Z, Fekete I, Krakomperger Z, Tóth J A, Lajtha K, Tóthmérész B. Soil enzyme activity in response to long-term organic matter manipulation. Soil Biology and Biochemistry, 2014, 70: 237- 243.
[15] Fekete I, Varga C, Kotroczó Z, Tóth J A, Várbíró G. The relation between various detritus inputs and soil enzyme activities in a Central European deciduous forest. Geoderma, 2011, 167- 68: 15- 21.
[16] 黃玉梅, 楊萬勤, 張健, 盧昌泰, 劉旭, 王偉, 郭偉, 張丹桔. 川西亞高山針葉林土壤微生物及酶對林下植物去除的響應. 生態(tài)學報, 2014, 34(15): 4183- 4192.
[17] Kivlin S N, Treseder K K. Soil extracellular enzyme activities correspond with abiotic factors more than fungal community composition. Biogeochemistry, 2014, 117(1): 23- 37.
[18] Mo H M, Brown S, Peng S L, Kong G H. Nitrogen availability in disturbed, rehabilitated and mature forests of tropical China. Forest Ecology and Management, 2003, 175(1/3): 573- 583.
[19] Zhang G L, Jiang H, Niu G D, Liu X W, Peng S L. Simulating the dynamics of carbon and nitrogen in litter-removed pine forest. Ecological Modelling, 2006, 195(3/4): 363- 376.
[20] Xiong Y M, Xia H P, Li Z A, Cai X A, Fu S L. Impacts of litter and understory removal on soil properties in a subtropicalAcaciamangiumplantation in China. Plant and Soil, 2008, 304(1/2): 179- 188.
[21] Huang Z Q, He Z M, Wan X H, Wan X H, Hu Z H, Fan S H, Yang Y S. Harvest residue management effects on tree growth and ecosystem carbon in a Chinese fir plantation in subtropical China. Plant and Soil, 2013, 364(1/2): 303- 314.
[22] 陳楚瑩, 張家武, 周崇蓮, 鄭鴻元. 改善杉木人工林的林地質(zhì)量和提高生產(chǎn)力的研究. 應用生態(tài)學報, 1990, 1(2): 97- 106.
[23] 中國生態(tài)系統(tǒng)研究網(wǎng)絡科學委員會. 陸地生態(tài)系統(tǒng)土壤觀測規(guī)范. 北京: 中國環(huán)境科學出版社, 2005.
[24] Matsushima M, Chang S X. Effects of understory removal, N fertilization, and litter layer removal on soil N cycling in a 13-year-old white spruce plantation infested with Canada bluejoint grass. Plant and Soil, 2007, 292(1/2): 243- 258.
[25] Fekete I, Kotroczó Z, Varga C, Nagy P T, Várbíró G, Bowden R D, Tóth J A, Lajtha K. Alterations in forest detritus inputs influence soil carbon concentration and soil respiration in a Central-European deciduous forest. Soil Biology and Biochemistry, 2014, 74: 106- 114.
[26] Wang Q K, He T X, Wang S L, Liu L. Carbon input manipulation affects soil respiration and microbial community composition in a subtropical coniferous forest. Agricultural and Forest Meteorology, 2013, 178- 179: 152- 160.
[27] Rutigliano F A, Castald S, D′Ascoli R, Papa S, Carfora A, Marzaioli R, Fioretto A. Soil activities related to nitrogen cycle under three plant cover types in Mediterranean environment. Applied Soil Ecology. 2009, 43(1): 40- 46.
[28] Sinsabaugh R L, Lauber C L, Weintraub M N, Ahmed B, Allison S D, Crenshaw C, Contosta A R, Cusack D, Frey S, Gallo M E, Gartner T B, Hobbie S E, Holland K, Keeler B L, Powers J S, Stursova M, Takacs-Vesbach C, Waldrop M P, Wallenstein M D, Zak D R, Zeglin L H. Stoichiometry of soil enzyme activity at global scale. Ecology Letters, 2008, 11(11): 1252- 1264.
[29] Hudson B D. Soil organic matter and available water capacity. Journal of Soil & Water Conservation, 1994, 49(2): 189- 194.
[30] Xu S, Liu L, Sayer E J. Variability of aboveground litter inputs alters soil physicochemical and biological processes: a meta-analysis of litterfall-manipulation experiments. Biogeosciences Discussions, 2013, 10(3): 5245- 5272.
Mechanism of litter and understory vegetation effects on soil carbon and nitrogen hydrolase activities in Chinese fir forests
YANG Yang1,2, WANG Jifu1, ZHANG Xinyu2,*, LI Dandan2,3, WANG Huimin2, CHEN Fusheng4, SUN Xiaomin2, WEN Xuefa2
1CollegeofGeographicScience,HarbinNormalUniversity,Harbin150025,China2KeyLaboratoryofEcosystemNetworkObservationandModeling,InstituteofGeographicSciencesandNaturalResourcesResearch,ChineseAcademyofSciences,Beijing100101,China3BiologicalScienceandTechnology,ShenyangAgriculturalUniversity,Shenyang110866,China4JiangxiProvincialKeyLaboratoryforBambooGermplasmResourcesandUtilization,JiangxiAgriculturalUniversity,Nanchang330045,China
litter; understory; Chinese fir; soil environmental factors;β- 1, 4-glucosidase activity;β- 1, 4-N-acetyl-glucosaminidase activity
國家重點基礎研究發(fā)展計劃資助項目(2012CB416903);國家自然科學基金資助項目(31130009, 31290222, 41571251, 41071332)
2015- 05- 04;
日期:2016- 04- 12
10.5846/stxb201505040908
*通訊作者Corresponding author.E-mail:zhangxy@igsnrr.ac.cn
楊洋,王繼富,張心昱,李丹丹,王輝民,陳伏生,孫曉敏,溫學發(fā).凋落物和林下植被對杉木林土壤碳氮水解酶活性的影響機制.生態(tài)學報,2016,36(24):8102- 8110.
Yang Y, Wang J F, Zhang X Y, Li D D, Wang H M, Chen F S, Sun X M, Wen X F.Mechanism of litter and understory vegetation effects on soil carbon and nitrogen hydrolase activities in Chinese fir forests.Acta Ecologica Sinica,2016,36(24):8102- 8110.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數(shù)理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
當代陜西(2021年2期)2021-03-29 07:41:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
