大熊貓糞便野外暴露時間對微衛星實驗的影響
2016-04-13 06:15:57王承東楊波黃杰黃炎成彥曦李仁貴黃山李德生張和民
四川動物 2016年4期
王承東, 楊波, 黃杰, 黃炎, 成彥曦, 李仁貴, 黃山, 李德生, 張和民*
(1.中國保護大熊貓研究中心,四川臥龍623006; 2. 四川省瀕危野生動物保護生物學重點實驗室,四川大學生命科學學院,成都610064; 3. 河南商丘師范學院生命科學學院,河南商丘476000)
大熊貓糞便野外暴露時間對微衛星實驗的影響
王承東1, 楊波1, 黃杰2, 3, 黃炎1, 成彥曦1, 李仁貴1, 黃山1, 李德生1, 張和民1*
(1.中國保護大熊貓研究中心,四川臥龍623006; 2. 四川省瀕危野生動物保護生物學重點實驗室,四川大學生命科學學院,成都610064; 3. 河南商丘師范學院生命科學學院,河南商丘476000)
非損傷性取樣,如從糞便中獲取DNA用于分子生物學研究已經在珍稀野生動物中得到了廣泛應用。DNA質量與后續研究很大程度上依賴于糞便暴露時間的長短。本研究通過分析不同新鮮程度糞便樣品的DNA質量和微衛星基因分型效果探討大熊貓糞便野外暴露時間對分子實驗的影響。研究表明,暴露時間越短,糞便DNA降解程度越低,所提取的DNA質量也較高。同時糞便DNA的降解程度與季節和暴露環境也有一定的關系:春、秋季14 d以內、夏季7 d以內、冬季21 d以內的糞便樣品提取的DNA質量較好,微衛星能成功分型;另外,野外臥穴口的糞便樣品DNA保存時間相對較長。本研究對野外大熊貓糞便采集工作具有指導意義。
大熊貓;糞便;暴露時間;微衛星
20世紀90年代中期出現的野生動物非損傷性取樣已得到廣泛應用(Taberletetal.,1999;陳璐,岳曦,2007;Roquesetal.,2014;Sugimotoetal.,2014)。在所有非損傷性取樣中,糞便最易收集,且對動物的干擾及負面影響最小,因此是非損傷性取樣中最具有潛在價值的研究材料(魏輔文等,2001;Zhanetal.,2006;Zhuetal.,2013)。在野生動物的物種鑒定(Roquesetal.,2014)、個體識別(Taberlet & Luikart,1999;Chakrabortyetal.,2014)、性別鑒定(Baumgardtetal.,2013;Roquesetal.,2014)、交配方式分析(Canigliaetal.,2014)、種群遺傳結構分析(Canigliaetal.,2014)、遺傳多樣性評估(Sugimotoetal.,2014)、基因流分析(Saarinenetal.,2014)、種群數量及保護管理單元的制定(方盛國等,1996)等方面已得到廣泛應用。
糞便暴露在野外環境中,DNA會因被核酸內切酶作用而降解,DNA降解隨著暴露時間的延長而增加,影響PCR的擴增甚至不能擴增(Frantzenetal.,1998)。雖然有研究表明,在野外暴露12 h甚至1 d的糞便樣品能夠獲得很好的實驗結果(Murphyetal.,2002;Chambersetal.,2004;Nsubugaetal.,2004),但DNA的降解受到多種因素的影響,如溫度、濕度、光照強度等(Femandoetal.,2003)。大熊貓Ailuropodamelanoleuca野生個體數量稀少,在野外生活十分警覺,遇見率極低,野外樣品的采集對糞便的依賴度更高,而且在野生環境中很難獲得新鮮糞便。因此,較準確地辨別大熊貓糞便暴露時間和確定暴露多久的糞便能夠用于進一步的分子生物學研究對于大熊貓野外種群數量調查(方盛國等,1996)、種群遺傳學(Luetal.,2001;Zhangetal.,2007;Shenetal.,2009;Yangetal.,2011;Zhuetal.,2011;Huangetal.,2015)以及野化放歸(楊波等,2013)等生態學問題的研究具有重要意義。本研究旨在探討不同新鮮程度糞便樣品的DNA提取質量與后續分子實驗的效果,為野外工作提供參考。
1 材料和方法
1.1 樣品采集與處理
2013年1月采集中國保護大熊貓研究中心臥龍核桃坪野化基地3只圈養大熊貓的糞便樣品,每只大熊貓采集2團糞便,做2個環境處理。仿野外環境:將糞便置于四川大學生命科學學院樓外草叢中(溫度9.97 ℃±3.16 ℃,濕度48.27%±16.38%);恒定環境:室內恒溫箱溫度22.5 ℃±0.5 ℃,濕度62.5%±2.5%。每周1次采樣并提取DNA,利用10個微衛星標記進行分析。
2013年6月獲取中國保護大熊貓研究中心雅安碧峰峽基地的大熊貓糞便樣品共24份,編號為YA01~YA24。樣品均置于人工竹林中,竹密度為每平方米31株,平均竹高1.5 m,竹蓋度92%,溫度21.4 ℃±2.0 ℃,濕度80.8%±10.7%,暴露不同時間用于取樣檢測。
2014年8月采集中國保護大熊貓研究中心臥龍核桃坪野化基地4只半野化大熊貓的糞便樣品,做4個環境處理。
A. 竹林地:竹密度為每平方米27株,竹蓋度95.5%,基質為腐竹葉和枯竹稈混合的松軟泥土,郁閉度0.92,25°左右的半陰陽坡,溫度16.6 ℃±2.9 ℃,濕度77.2%±28.3%。
B. 苔蘚地:竹密度為每平方米12株,竹蓋度90%,基質為潮濕的苔蘚地,郁閉度0.85,2°左右的半陰陽坡,為竹林邊緣地帶,類似大熊貓過道的生境類型,溫度16.8 ℃±3.0 ℃,濕度77.7%±28.0%。
C. 臥息地:高大喬木底部,基質為覆蓋有枯枝葉的硬質泥地,郁閉度0.90,18°左右的半陰陽坡,溫度17.2 ℃±2.8 ℃,濕度80.0%±30.0%。
D. 臥穴口:位于巖石底部內凹處,基質為碎石,郁閉度0.78,10°左右的陰坡,溫度17.3 ℃±2.8 ℃,濕度76.0%±27.0%。
2015年9月下旬采集中國保護大熊貓研究中心都江堰基地的圈養大熊貓糞便樣品,做2個環境處理。
E. 草地:竹蓋度約95%,松軟泥土,上蓋腐竹葉,并夾生草本植物,陰涼潮濕;溫度19.5 ℃±3.5 ℃,濕度64.5%±6.8%。
F. 裸露地:完全暴露在陽光下的碎石地,相對干燥,東南面有一建筑,距離約1 m;溫度20.0 ℃±2.9 ℃,濕度61.9%±3.9%。
糞便新鮮程度共設計6個時間梯度,即1 d、3 d、7 d、14 d、21 d和28 d。
1.2 糞便DNA提取
大熊貓糞便樣品DNA提取使用德國Qiagen生產的試劑盒QIAamp DNA Stool Kit,步驟及方法按廠商使用說明操作,瓊脂糖凝膠電泳初步檢測DNA提取是否成功、DNA濃度及質量,-20 ℃保存。
1.3 微衛星位點選擇
Huang等(2015)基于大熊貓基因組微衛星篩選了能夠有效使用的大熊貓糞便DNA的微衛星位點。本文使用了其中10個微衛星位點,并對每1個微衛星位點的上游引物F’端進行熒光標記。
1.4 PCR反應條件及基因分型
25 μL PCR反應體系:25 ng DNA,2.5 μL 10×PCR buffer,1.0~2.0 μm MgCl2,200 μM dNTP,上、下游引物各1.0 μm,1 U Taq酶,ddH2O補足至25 μL。
PCR反應程序:95 ℃ 4 min;95 ℃ 30 s,60 ℃ 50 s,72 ℃ 50 s,35個循環;72 ℃延伸10 min。PCR反應在iCycler PCR反應儀(Bio-Rad,USA)中進行,PCR產物4 ℃避光保存,每個樣本PCR產物取5 μL,用1%瓊脂糖電泳檢測每個樣本是否成功擴增,確定獲得產物為單一、大小正確的DNA片段。各個樣本PCR產物使用377 DNA sequencer(ABI PRISM,USA)進行基因分型,使用GeneMapper v3.2確定樣本等位基因數,等位基因大小相對于分子內標ROX-500[基點認知技術(北京)有限公司]決定。基因分型由北京閱微基因公司完成。
1.5 數據分析

2 結果
2.1 DNA提取質量
糞便樣品暴露時間的不同,所提取的DNA質量存在差異。2013年1月放置于四川大學生命科學學院樓外草叢中的糞便樣品暴露不同時間后,提取DNA電泳檢測發現,14 d以內均能獲得較高質量的DNA(圖1)。2013年6月放置于雅安碧峰峽基地人工竹林中的糞便樣品,暴露7 d后已有一些降解,但仍有部分樣品能夠提取較高質量的DNA(編號為13~15),而暴露了18 d(編號為16~19)和30 d的樣品(編號為20~24)提取的DNA質量較差,降解嚴重(圖2)。2014年8月放置于臥龍核桃坪野化基地野生環境下的樣品(圖3),3 d內均能提取質量較好的DNA,從7 d后開始出現不同程度的降解;而臥穴口的糞便即使暴露14 d后,其DNA質量仍然較好。2015年9月下旬放置于都江堰基地模擬野生環境下的樣品,暴露21 d后還可以提取出較好質量的DNA(圖4)。表明不同環境下,糞便樣品DNA的保存時間差異較大。
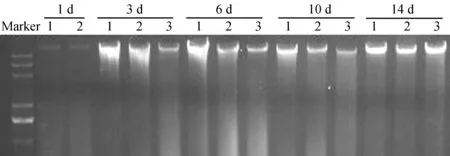
圖1 2013年1月放置于四川大學生命科學學院樓外的大熊貓糞便樣品DNA提取質量Fig. 1 DNA extracted from Ailuropoda melanoleuca feces exposed outside the building of College of Life Sciences, Sichuan University, January 2013

圖2 2013年6月放置于中國保護大熊貓研究中心雅安碧峰峽基地大熊貓糞便樣品DNA提取質量Fig. 2 DNA extracted from Ailuropoda melanoleuca feces exposed in the field of Ya’an Bifeng Gorge Base, China Conservation and Research Center for Giant Panda, Sichuan, June 2013

圖3 2014年8月放置于中國保護大熊貓研究中心臥龍核桃坪野化基地大熊貓糞便樣品DNA提取質量Fig. 3 DNA extracted from Ailuropoda melanoleuca feces exposed in the field of Wolong Base, China Conservation and Research Center for Giant Panda, Sichuan, August 2014A.竹林地 bamboo forest, B.苔蘚地 moss, C.臥息地 bed site, D.臥穴口 entrance of cave; 下同, the same below.
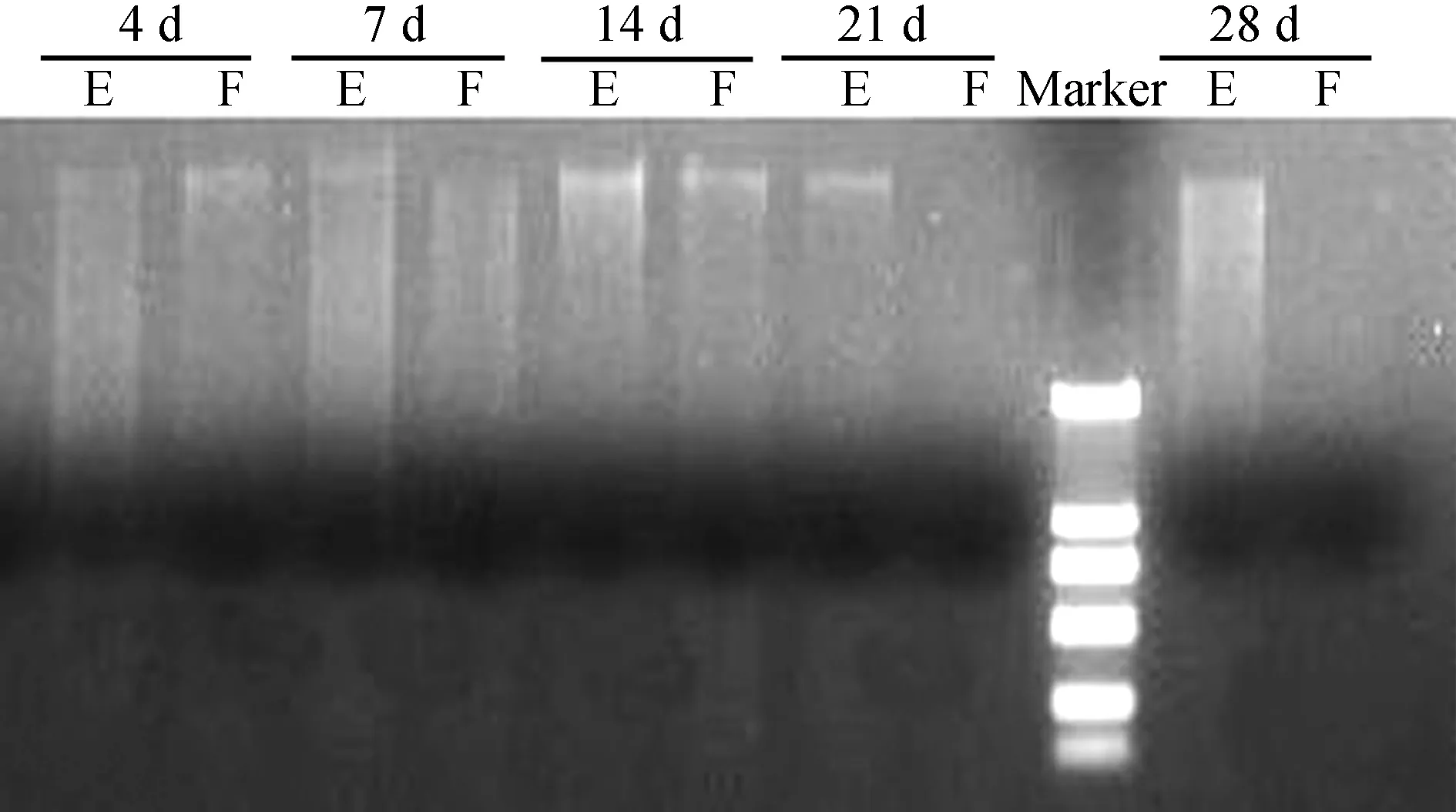
圖4 2015年9月放置于中國保護大熊貓研究中心 都江堰基地大熊貓糞便樣品DNA提取質量Fig. 4 DNA extracted from Ailuropoda melanoleuca feces that exposed in the field of Dujiangyan Disease Control Center, China Conservation and Research Center for Giant Panda, Sichuan, September 2015
E.草地 grassland, F.裸露地 open ground; 下同, the same below.
2.2 微衛星基因分型結果
利用10個微衛星位點對不同暴露時間糞便樣品的DNA進行了PCR擴增及基因分型。每個樣品3次重復,結果無差異,同環境、同暴露時間樣品間的分析結果差異無統計學意義(P>0.05)。不同環境中的樣品分析結果存在一定差異,即樣品DNA的PCR擴增成功率均隨著暴露時間的增加而降低。樣品暴露35 d后,環境導致的差異有統計學意義(P<0.05)。置于恒溫(22.5 ℃±0.5 ℃)、恒濕(62.5%±2.5%)的環境條件下,暴露35 d后,選取的10個位點中有6個以上的位點能成功分型;而置于室外環境條件下(溫度9.97 ℃±3.16 ℃),由于溫度較低,暴露42 d后樣品中的10個位點中仍有6個以上的位點能成功分型(表1)。
根據Huang等(2015)建立的個體識別系統,15個微衛星位點中,使用6個位點能夠成功進行大熊貓的個體識別和親子鑒定。放置于中國保護大熊貓研究中心雅安碧峰峽基地模擬野化環境中的糞便樣品,基因分型結果顯示,暴露18 d內的樣品平均有6個以上的位點能成功分型,能夠用于個體識別研究。而超過18 d的樣品,DNA降解較為嚴重,給基因分型帶來一定的難度,而超過30 d的樣品,分型成功率僅為45%,不能滿足個體識別及親子鑒定的要求(表2)。
2014年8月放置于中國保護大熊貓研究中心臥龍核桃坪野化基地野外環境中的糞便樣品暴露7 d后,微衛星分型成功率出現明顯下降,苔蘚地樣品的分型成功率只有50%左右,不能用于后續分析。不管怎樣,臥穴口樣品的DNA保存較好,暴露21 d后仍有大約60%的位點可以成功分型。2015年9月放置于中國保護大熊貓研究中心都江堰基地的樣品暴露7 d后DNA保存較好,約有60%以上的位點可以成功分型,暴露14 d時,草地樣品的DNA降解嚴重,成功分型的位點降至一半以下,而裸露地樣品的位點仍有60%以上能夠成功分型,可以進行后續分析(表3)。

表1 2013年1月四川大學生命科學學院樓外大熊貓糞便樣品在不同暴露時間下微衛星成功分型的平均位點數)Table 1 The mean numbers of successfully genotyped microsatellite loci using Ailuropoda melanoleuca feces DNA (S1, S2, and S3) with different exposure times outside the building of College of Life Sciences, Sichuan University )
注Notes:*P<0.05,**P<0.01; In. 恒溫恒濕環境constant temperature and humidity environment, Out. 室外環境outdoor environment; 下表同, the same below.
表2 2013年6月在中國保護大熊貓研究中心雅安碧峰峽基地大熊貓糞便樣品在不同暴露時間下微衛星成功分型的平均位點數)Table 2 The mean numbers of successfully genotyped microsatellite loci using Ailuropoda melanoleuca feces DNA with different exposure times in Ya’an Bifeng Gorge Base, China Conservation and Research Center for Giant Panda, Sichuan )

表3 2014年8月在中國保護大熊貓研究中心臥龍核桃坪野化基地和2015年9月在都江堰基地 大熊貓糞便樣品在不同暴露時間下微衛星成功分型的平均位點數)Table 3 The mean numbers of successfully genotyped microsatellite loci using Ailuropoda melanoleuca feces DNA with different exposure times in the field of Wolong Base (A, B, C, D) and in the field of Dujiangyan Disease Control Center (E, F), China Conservation and Research Center for Giant Panda, Sichuan )
3 討論
本研究旨在探討大熊貓糞便暴露時間與DNA提取質量以及后續分子生物學研究的關系,進而對野外糞便采集提供參考。本研究于2012年12月—2013年1月,在四川大學校園內模擬野外環境暴露糞便樣品,發現冬季寒冷干燥的環境最適于糞便的保存,即使暴露時間長達42 d,依然可以獲得較高質量的DNA。但是,2013年6月在更接近野外環境的中國保護大熊貓研究中心雅安碧峰峽基地的人工竹林中,暴露18 d的樣品降解嚴重,這可能由于研究期處于初夏,正是多雨時節,溫、濕度高,糞便DNA降解速度快。2014年8月在有野生大熊貓分布的中國保護大熊貓研究中心臥龍核桃坪野化基地的野外環境中,暴露7 d的糞便樣品DNA降解就較為嚴重,8月氣溫更高,相關酶活性也更高,DNA降解更快。因此不同季節,糞便樣品中DNA的保存時間差異顯著,建議野外糞便樣品的采集盡可能選擇在氣溫較低、少雨的秋、冬季進行。
糞便DNA降解嚴重與否,除與氣溫和天氣有關外,與糞便暴露的環境也有一定的關系。野外環境中的植被類型、郁閉度、灌木蓋度、坡度和坡向等因子直接影響環境中的溫度和濕度,因而與糞便中DNA的保存時間必然相關。先前研究表明,四川野生大熊貓常出現在郁閉度大于0.50的落葉闊葉林和針闊混交林中,微生境為竹林,其多選擇平均高度為2~5 m、蓋度大于50%的竹林中覓食,喜歡在平緩的(坡度在6~30°范圍內)東南坡向或陽坡與半陽坡的生境中活動,在海拔2 000~3 000 m的活動較多(魏輔文,馮祚建,1999;張澤鈞,胡錦矗,2000;康東偉等,2011;趙秀娟等,2012)。本研究中臥龍的試驗樣地符合大熊貓日常活動的生境。研究表明暴露7 d的糞便樣品中的DNA約有一半以上的微衛星位點能夠成功分型,且微生境較干燥的環境,比如臥穴口糞便樣品中的DNA保存時間更長,暴露14 d以內的糞便可以滿足微衛星分析要求,甚至暴露21 d的糞便也可以分析。這些結果可以為野外糞便的收集提供參考。
4 糞便樣品采集建議
在野外糞便樣品采集時,需要根據樣品的外形特點,如糞便表皮黏膜組織的干燥程度、黏膜的光澤度、黏膜厚度,判斷糞便樣品暴露的大致時間。據野外觀察,在3 d以內排出的大熊貓糞便很容易辨認,糞便表面的黏液完整,有光澤,觸摸有光滑細膩的感覺,打開即能嗅到一股清香的竹味,表面顏色與里面相同,成色新鮮,糞團表面無霉菌滋生(郭建,胡錦矗,2001)。暴露7 d的糞便在濕熱的微環境下,其表面無黏膜,有少量白色菌絲,糞便稍松散;而在較干燥的環境下,其表面有一層猶如薄紙狀的略帶光澤的黏膜,無菌絲滋生,外形保持完整。暴露14 d的糞便松散,表面霉菌明顯增多,菌絲長度在1 cm左右,有霉味;較干燥環境下的糞便與暴露7 d時的幾乎無變化。暴露21 d的糞便外形松散嚴重,霉味嚴重,霉菌布滿糞便表面,菌絲最長長度在2 cm左右;較干燥的環境下,糞便干裂,表面無薄紙狀黏膜或極少。
基于本研究結果,我們對野外大熊貓糞便的采集有以下建議:(1)野外糞便采集盡量選擇在秋、冬季開展,糞便暴露時間最好不超過14 d,如果需要在春、夏季采樣,則糞便暴露時間最好不超過7 d。(2)野外糞便采集時,盡量選擇干燥、透氣微生境中的糞便,因為干燥、透氣環境中糞便DNA保存時間較長。(3)采集時記錄清晰、詳盡,并注意重復采樣,以便后續研究。
陳璐, 岳曦. 2007. 非損傷性取樣研究進展[J]. 四川動物, 26(1): 224-226.
方盛國, 陳冠群, 馮文和, 等. 1996. 大熊貓DNA指紋在野生種群數量調查中的應用[J]. 獸類學報, 16(4): 246-249.
郭建, 胡錦矗. 2001. 大熊貓糞團分布型的研究及其應用[J]. 獸類學報, 21(8): 180-186.
康東偉, 趙志江, 郭文霞, 等. 2011. 大熊貓的生境選擇特征[J]. 應用生態學報, 22(2): 519-525.
魏輔文, 馮祚建. 1999. 相嶺山系大熊貓和小熊貓對生境的選擇[J]. 動物學報, 45(1): 57-63.
魏輔文, 饒剛, 李明, 等. 2001. 分子糞便學及其應用——可靠性、局限性和展望[J]. 獸類學報, 21(2): 143-152.
楊波, 楊承忠, 涂飛云, 等. 2013. 大熊貓野化放歸中的遺傳學分析[J]. 四川動物, 32(1): 149-155.
張澤鈞, 胡錦矗. 2000. 大熊貓生境選擇研究[J]. 西華師范大學學報(自然科學版), 21(1): 234-239.
趙秀娟, 張澤鈞, 胡錦矗. 2012. 唐家河與蜂桶寨自然保護區大熊貓生境選擇初步比較[J]. 西華師范大學學報(自然科學版), 33(3): 234-239.
Baumgardt JA, Goldberg CS, Reese KP,etal. 2013. A method for estimating population sex ratio for sage-grouse using noninvasive genetic samples[J]. Molecular Ecology Resources, 13(3): 393-402.
Caniglia R, Fabbri E, Galaverni M,etal. 2014. Noninvasive sampling and genetic variability, pack structure, and dynamics in an expanding wolf population[J]. Journal of Mammalogy, 95(1): 41-59.
Chakraborty S, Boominathan D, Desai AA,etal. 2014. Using genetic analysis to estimate population size, sex ratio, and social organization in an Asian elephant population in conflict with humans in Alur, southern India[J]. Conservation Genetics, 15(4): 897-907.
Chambers KE, Reichard UH, M?ller A,etal. 2004. Cross-species amplification of human microsatellite markers using noninvasive samples from white-handed gibbons (Hylobateslar)[J]. American Journal of Primatology, 64(1): 19-27.Femando P, Vidya TN, Rajapakse C,etal. 2003. Reliable noninvasive genotyping: fantasy of reality?[J]. Journal of Heredity, 94(2): 115-123.
Frantzen MA, Silk JB, Ferguson JW,etal. 1998. Empirical evaluation of preservation methods for faecal DNA[J]. Molecular Ecology, 7(10): 1423-1428.
Huang J, Li YZ, Du LM,etal. 2015. Genome-wide survey and analysis of microsatellites in giant panda (Ailuropodamelanoleuca), with a focus on the applications of a novel microsatellite marker system[J]. BMC Genomics, 16(1): 1-12.
Lu Z, Johnson WE, Menotti M,etal. 2001. Patterns of genetic diversity in remaining giant panda populations[J]. Conservation Biology, 15(6): 1596-1607.
Murphy MA, Waits LP, Kendall KC,etal. 2002. An evaluation of long-term preservation methods for brown bear (Ursusarctos) faecal DNA samples[J]. Conservation Genetics, 3(4): 435-440.
Nsubuga AM, Robbins MM, Roeder AD,etal. 2004. Factors affecting the amount of genomic DNA extracted from ape faeces and the identification of an improved sample storage method[J]. Molecular Ecology, 13(7): 2089-2094.
Oosterhout CV, Hutchinson WF, Wills DPM,etal. 2004. Micro-checker: software for identifying and correcting genotyping errors in microsatellite data[J]. Molecular Ecology Notes, 4(3): 535-538.
Park S. 2001. Trypanotolerance in west African cattle and the population genetic effects of selection[D]. Dublin, Ireland: University of Dublin.
Roques S, Furtado M, Jácomo ATA,etal. 2014. Monitoring jaguar populationsPantheraoncawith non-invasive genetics: a pilot study in Brazilian ecosystems[J]. Oryx, 48(3): 361-369.
Saarinen EV, Daniels JC, Maruniak JE. 2014. Local extinction event despite high levels of gene flow and genetic diversity in the federally-endangered Miami blue butterfly[J]. Conservation Genetics, 15(15): 811-821.
Shen FJ, Zhang ZH, He W,etal. 2009. Microsatellite variability reveals the necessity for genetic input from wild giant pandas (Ailuropodamelanoleuca) into the captive population[J]. Molecular Ecology, 18(6): 1061-1070.
Sugimoto T, Aramilev VV, Kerley LL,etal. 2014. Noninvasive genetic analyses for estimating population size and genetic diversity of the remaining far eastern leopard (Pantherapardusorientalis) population[J]. Conservation Genetics, 15(3): 521-532.
Taberlet P, Luikart G. 1999. Noninvasive genetic sampling and individual identification[J]. Biological Journal of the Linnean Society, 68(1-2): 41-55.
Taberlet P, Waits LP, Luikart G. 1999. Noninvasive genetic sampling: look before you leap[J]. Trends in Ecology & Evolution, 14(8): 323-327.
Yang JD, Hou R, Shen FJ,etal. 2011. Microsatellite variability reveals significant genetic differentiation of giant pandas (Ailuropodamelanoleuca) in the Minshan A habitat[J]. African Journal of Biotechnology, 10(60): 12804-12811.
Zhan X, Li M, Zhang Z,etal. 2006. Molecular censusing doubles giant panda population estimate in a key nature reserve[J]. Current biology, 16(12): R451-R452.
Zhang BW, Li M, Zhang ZJ,etal. 2007. Genetic viability and population history of the giant panda, putting an end to the “evolutionary dead end”?[J]. Molecular Biology and Evolution, 24(8): 1801-1810.
Zhu L, Hu Y, Qi D,etal. 2013. Genetic consequences of historical anthropogenic and ecological events on giant pandas[J]. Ecology, 94(10): 2346-2357.
Zhu LF, Zhang SN, Gu XD,etal. 2011. Significant genetic boundaries and spatial dynamics of giant pandas occupying fragmented habitat across southwest China[J]. Molecular Ecology, 20(6): 1122-1132.
Effect of Exposure Time ofAiluropodamelanoleucaFeces on Microsatellites Analysis
WANG Chengdong1, YANG Bo1, HUANG Jie2, 3, HUANG Yan1, CHENG Yanxi1,LI Rengui1, HUANG Shan1, LI Desheng1, ZHANG Hemin1*
(1. China Conservation and Research Center for the Giant Panda, Wolong, Sichuan Province 623006, China; 2. Sichuan Key Laboratory of Conservation Biology on Endangered Wildlife, College of Life Sciences, Sichuan University, Chengdu 610064,China; 3. College of Life Sciences, Shangqiu Normal University, Shangqiu, Henan Province 476000, China)
Noninvasive sampling methods, such as acquiring host genomic DNA from fecal samples for molecular biology research, are increasingly used in endangered wild animals. However, it is difficult to obtain fresh feces in the field. By focusing on the effects of feces after different exposure times in the field on molecular studies for giant pandas (Ailuropodamelanoleuca), we found that shorter exposure time of feces would lower the level of DNA degradation, and increase the quality of the feces DNA. The degradation level of feces DNA was also related to seasonal variation and field environment. The valid time was no longer than 14 days in spring and autumn, less than 7 days in summer and no longer than 21 days in winter. Moreover, dry environment such as the entrance of the cave may be the best place for preserving feces samples in the field. The results of this study provided guidance for the feces collection of wild giant pandas in the field.
Ailuropodamelanoleuca; feces; exposure time; microsatellite
2016-02-16 接受日期:2016-03-24
大熊貓國合資金項目(AD1413):大熊貓飼養、繁育及管理; 重點區域野生大熊貓疫病本底調查與監測
王承東, 博士, 高級工程師, 主要從事大熊貓飼養管理、獸醫及放歸研究
*通信作者Corresponding author, E-mail:wolong-zhm@126.com
10.11984/j.issn.1000-7083.20160033
Q958.1; Q953
A
1000-7083(2016)04-0481-07
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中老年保健(2021年12期)2021-08-24 03:30:40
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
科技傳播(2019年22期)2020-01-14 03:06:54
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年11期)2019-01-07 01:28:02
