新型全反式維甲酸衍生物 4-氨基-2-三氟甲基苯基維甲酸酯對肝癌細胞株 HepG2增殖的影響
2016-04-27 08:08:42劉慧,周青,汪淵
安徽醫科大學學報 2016年2期
關鍵詞:肝癌
劉 慧,周 青,汪 淵
新型全反式維甲酸衍生物 4-氨基-2-三氟甲基苯基維甲酸酯對肝癌細胞株 HepG2增殖的影響
劉 慧,周 青,汪 淵
目的研究 4-氨基-2-三氟甲基苯基維甲酸酯 (ATPR)對人肝癌細胞株 HepG2增殖的影響,并初步探討其可能的作用機制。方法選取 HepG2細胞為研究對象,全反式維甲酸(ATRA)和 ATPR分別作用細胞,應用 MTT比色法測定細胞增殖,流式細胞術檢測細胞周期分布情況,Western blot法檢測 HepG2細胞內周期蛋白 D(cyclinD)及細胞周期蛋白依賴性激酶 4(CDK4)和 CDK6蛋白表達量。結果ATPR能夠顯著抑制 HepG2細胞的增殖,將細胞阻滯在 G0/G1期,呈濃度與時間依賴性(P<0.05),并且同濃度、同一處理時間下,ATPR的抑 制率更高(P<0.05);此外,Western blot結果顯示,ATPR較 ATRA更能顯著下調 HepG2細胞中cyclinD和 CDK6蛋白表達水平(P<0.05),而 CDK4蛋白水平在各組變化均不明顯。結論同等濃度下,ATPR抑制HepG2細胞增殖的效果明顯強于 ATRA,其機制可能是 ATPR通過下調 cyclinD和 CDK6蛋白的表達水平,造成 G0/G1期阻滯,最終導致肝癌細胞株 HepG2的增殖抑制。
肝癌;HepG2細胞;全反式維甲酸;4-氨基-2-三氟甲基苯基維甲酸酯;細胞周期;細胞增殖
全 反 式 維 甲 酸 (all-trans retinoic acid,ATRA)(圖1)是維生素 A的主要活性代謝物,為維甲酸類藥物的代表,可誘導分化多種腫瘤細胞,已被廣泛用于治療急性早幼粒細胞性白血病[1]。Sait et al[2]發現 ATRA能上調腫瘤細胞中的血管內皮細胞生長因子(vascular endothelial growth factor,VEGF),促進腫瘤血管的生成而引起惡性增殖。此外,ATRA還可產生較強的維甲酸綜合征,長期使用會產生維甲酸抵抗,限制了其在腫瘤治療中的作用[3]。因此,開發低毒高效的新型維甲酸衍生物是必需的。4-氨基-2-三氟甲基苯基維甲酸酯 (4-amino-2-trifluoromethyl-phenyl retinate,ATPR)(圖2)是由本校合成的新型維甲酸衍生物,研究[4-6]表明在抑制細胞遷移與誘導分化和凋亡方面,ATPR效果明顯優于 ATRA。然而關于 ATPR對肝癌細胞株 HepG2增殖的研究較少,相關的機制尚不清楚。該研究以肝癌細胞株 HepG2為模型,通過比較 ATRA與 ATPR對其增殖能力的影響,為新型維甲酸衍生物用于肝癌臨床治療提供科學依據。
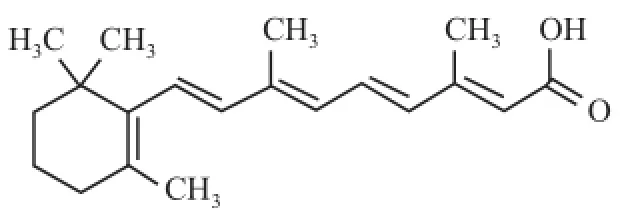
圖1 ATRA結構式
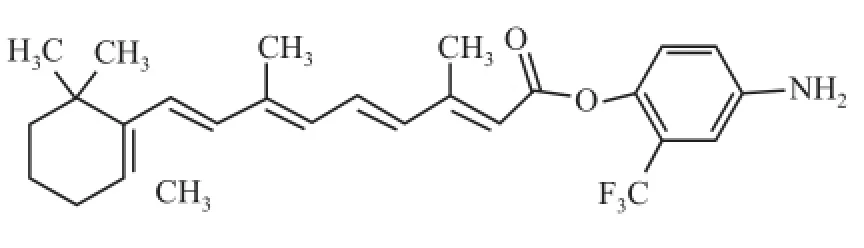
圖2 ATPR結構式
1 材料與方法
1.1 材料人肝癌細胞株 HepG2購自美國 ATCC公司;ATPR由安徽醫科大學藥學院提供;ATRA、二甲基亞砜(DMSO)、四甲基偶氮唑藍(MTT)購自美國 Sigma公司;低 糖 DMEM 培養基購自美國 Gibco公司,4℃儲存備用;新生牛血清購自杭州四季青生物工程材料有限公司,-20℃保存,用前 4℃融化,無需滅活;Coulter DNA檢測試劑盒購自美國 Beckman Coulter公 司;鼠 抗 人 β-actin購 自 美 國 Santa Cruz公司;鼠抗人 cyclinD、CDK6以及兔抗人 CDK4抗體購自上海聯世生物科技有限公司;辣根過氧化酶(horseradish peroxidase,HRP)標記的羊抗鼠 IgG、羊抗兔 IgG、ECL顯色試劑盒以及 BCA定量試劑盒均購自美國 Pierce公司。ATRA、ATPR用 DMSO溶解配制成 20 mmol/L,-20℃避光保存,使用時用完全培養基稀釋至使用濃度。
1.2 方法
1.2.1細胞株以及細胞培養 人肝癌細胞株HepG2用含 10%新生牛血清的低糖 DMEM培養基,并加入青霉素 100 U/ml和鏈霉素 100μg/m l,在 37℃、5%CO2飽和濕度的細胞培養箱中培養。每 2~3 d換一次液,當細胞密度達 80% ~90%,用 0.25%胰酶(含 0.53 mmol/L EDTA)傳代,取對數生長期的細胞進行實驗。
1.2.2細胞增殖實驗(MTT法) 收集對數生長期細胞,以 2.5×107/L的密度接種于 96孔板,每孔200μl,邊緣孔用 200μl PBS填充。待細胞貼壁后棄去培養液,用 PBS輕洗 3遍,然后分別加入含有不同濃度(5、12.5、20、25、50、75、100μmol/L)的ATRA和 ATPR的培養液孵育 48 h,以及同一濃度(25μmol/L)ATRA和 ATPR處理不同的時間(24、48、72、96 h),并設置細胞對照組、溶劑對照組(DMSO),其中溶劑對照組是與最大濃度 100μmol/L藥物中所含有的 DMSO等體積,其他藥物組均將 DMSO體積補齊至最大體積,以消除各組溶劑體積不同所造成的干擾。指定作用時間后,每孔加入 20μl MTT,于細胞培養箱中孵育 4~6 h。小心棄去上清液,每孔加入 100μl DMSO,37℃振動 15 min以溶解結晶,酶 標 儀 檢 測 570 nm 波 長 吸 光 度 (optical density,OD)。每個濃度設置 6個復孔,做 3次實驗,取平均值。抑制率(%)=(對照組 OD570-試驗組 OD570)/對照組 OD570×100%。用 SPSS 16.0計算藥 物 的 半 數抑制濃度 (half inhibitory concentration,IC50)。
1.2.3流式細胞術檢測細胞周期 收集處于對數生長期的 HepG2細胞,接種于 6孔板,待細胞貼壁后換成無血清培養基饑餓24 h,使細胞周期同步化,然后用含 25μmol/L的 ATRA和 ATPR的完全培養液處理細胞,48 h后常規消化收集各組細胞,預冷PBS洗3次后,用100μl PBS重懸并調整細胞密度為 1×106/ml。按 Coulter DNA檢測試劑盒操作,每組加入50μl固定破膜劑,反應30 s后立即加入500 μl PI染液,室溫避光反應 30 min后用流式細胞儀檢測,每個標本檢測 105個細胞,重復計數 3次,由ModFIT軟件分析 G0/G1、S、G2/M期細胞比例。
1.2.4Western blot檢測蛋白表達 5、25μmol/L的 ATRA和 ATPR分別處理 HepG2細胞 48 h后,取出細胞,用預冷的 PBS洗滌 3遍,在濾紙上控干PBS,并吸去殘余 PBS。然后每組加入 200μl蛋白提取緩沖液(Tris-HCl,pH 7.14,150 mmol/L NaCl,1 mmol/L EDTA,1%Triton X-100,0.1%SDS,5 mg/ml Leupeptin,1 mmol/L PMSF),冰 上 放 置 20 min后,置冰上用細胞刮刮取細胞,移入 1.5 ml EP管,并反復凍融 3次(-80℃、4℃)。之后,在超速冷凍離心機中 4℃、14 000 r/min離心 30 min,吸取上清液,使用 BCA試劑盒定量,用生理鹽水調整至相同濃度后,加入等體積的 4×蛋白上樣緩沖液,煮沸 8~10 min,-80℃保存待用。等量的各組樣品用于 SDS-PAGE電泳,然后將蛋白轉移到 PVDF膜上。轉膜成功后,室溫下用含 5%的脫脂奶粉封閉2 h或者4℃封閉過夜,接著將膜放入用一抗稀釋液配制的一抗中,4℃孵育過夜。經 TBST(含 0.05% Tween-20)洗滌 3遍后加入相應濃度 HRP標記的二抗室溫孵育 2 h,用 TBST洗滌 3遍后,TBS洗滌 1遍,每遍 10 min。在暗室中將等比例混合后的 ECL試劑 A液和 B液均勻涂于膜上,X線片曝光,顯影,定影。底片掃描后使用 Quantity One軟件分析條帶的灰度值。
1.3 統計學處理采用 SPSS 16.0軟件進行分析,實驗數據均以 ˉx±s表示;組間數據比較采用單因素方差分析,兩兩比較采用 LSD-t檢驗。
2 結果
2.1 ATRA和ATPR對HepG2細胞增殖的影響
MTT結果顯示,當細胞經不同濃度的 ATRA和ATPR處理 48 h后,與細胞對照組相比,低濃度的ATRA組(5、12.5、20、25μmol/L)對細胞無抑制作用,當作用濃度升至 50μmol/L,開始以劑量依賴方式抑制 HepG2細胞株的增殖(FATRA=53.919,P<0.05),而 ATPR組從低濃度到高濃度均能抑制細胞的生長,并呈濃度依賴關系(FATPR=8 269.029,P<0.05),見圖3。通過 SPSS 16.0計算出 ATRA和ATPR的 IC50分別為 102.190、31.723μmol/L。隨后選取 25μmol/L藥物濃度分別作用于 HepG2細胞24、48、72、96 h,隨著藥物處理時間的延長,抑制率逐漸升高。相比之下,同一處理時間下 ATPR較ATRA的抑制率更高(F24h=44.499,F48h=271.387,F72h=1 479.019,F96h=2 130.531,P<0.05),見圖4。以上結果均提示,與同一濃度的 ATRA相比,ATPR對 HepG2細胞增殖抑制作用更強。
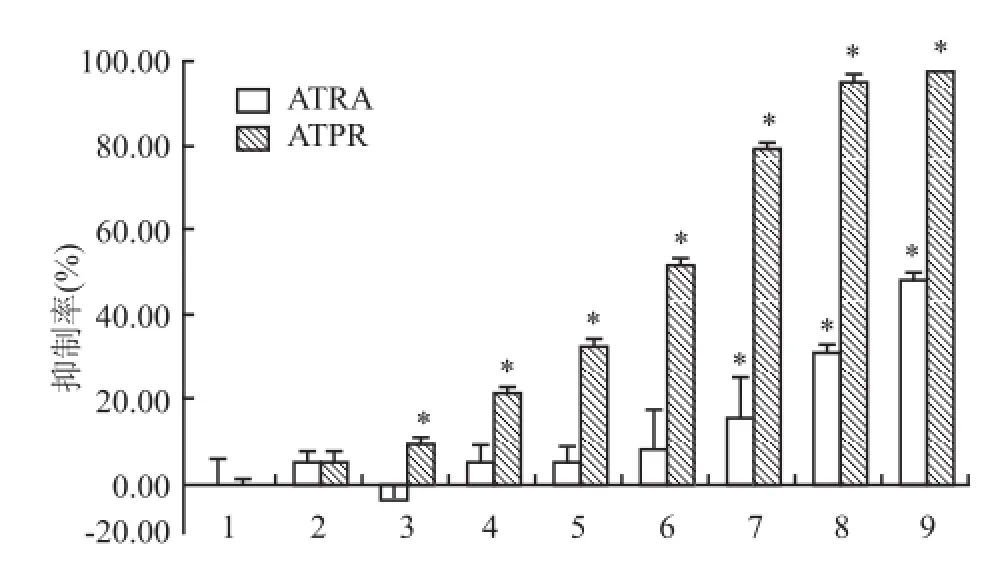
圖3 不同濃度的 ATRA及 ATPR對 HepG2細胞增殖的作用1:細胞對照組;2:溶劑對照組;3:5μmol/L;4:12.5μmol/L;5:20μmol/L;6:25μmol/L;7:50μmol/L;8:75μmol/L;9:100μmol/L;與細胞對照組比較:*P<0.05
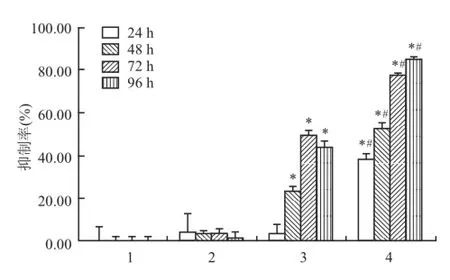
圖4 ATRA和 ATPR對 HepG2細胞增殖的作用1:細 胞對 照組;2:溶 劑 對 照 組;3:ATRA 25μmol/L;4:ATPR 25 μmol/L;與細胞對照組比較:*P<0.05;與同一時間的 ATRA組比較:#P<0.05
2.2 ATRA以及ATPR對HepG2細胞周期的影響25μmol/L ATRA和 ATPR處理 HepG2細胞 48 h后,流式細胞術檢測細胞周期顯示,與溶劑對照組相比,25μmol/L的 ATPR能夠顯著抑制 HepG2細胞增殖(P<0.05),使 G0/G1期細胞的比例明顯增加,而 同 濃 度 的 ATRA 組 無 明 顯 作 用 (FG0/G1= 123.523,FS=74.587,FG2=63.385),見圖5。以上結果提示 ATPR能將 HepG2細胞周期阻滯在 G0/G1期,從而抑制細胞增殖。
2.3 ATRA及ATPR對HepG2細胞中cyclinD、CDK 4和CDK 6蛋白水平的影響5、25μmol/L ATRA和 ATPR作用細胞 48 h后,與細胞對照組相比,ATPR能明顯下調 cyclinD與 CDK6的水平(P<0.05),而 ATRA 組 變 化 不 顯 著 (FcyclinD=65.190、FCDK6=65.651)。同時,CDK4蛋白的表達水平在各組中均變化不明顯(FCDK4=5.298),見圖6。
3 討論
肝癌是我國最常見的惡性腫瘤之一,經治療的早期肝癌,5年生存率僅為 50%,占惡性腫瘤死亡率的第二位[7-8]。除手術切除外,藥物化療是治療肝癌的重要手段,但由于療效缺乏特異性,并且長期用藥會產生 耐 性,從 而 影響了藥物 治 療 效 果[9]。因此,尋找高效、安全的新型抗癌藥是必要的。本實驗室采用的 ATPR是在 ATRA的結構基礎上進行羧基改造而成。研究[4-5,10]證實,ATPR可以有效抑制肺癌細胞株 A549、胃癌細胞株 BGC-823以及乳腺癌細胞株 MDA-MB-231的遷移。
細胞增殖是通過細胞分裂方式增加細胞數量的復雜過程,是細胞生命活動的重要特征之一。然而,當正常細胞在多種因素的影響下,細胞周期調節失控,獲得自主生長信號,導致細胞無限增殖,就會產生腫瘤[11],肝癌的發生發展也是肝癌細胞無限增殖的結果。因此,該文主要從細胞增殖方面,研究 ATRA與新型維甲酸衍生物 ATPR對肝癌 HepG2細胞株的作用。MTT實驗顯示,ATPR從低濃度到高濃度(5~100μmol/L)均能夠抑制 HepG2細胞的增殖,有濃度依賴性和時間依賴性,抑制率遠遠高于同等濃度下的 ATRA,即在相同濃度與作用時間下,ATPR抑制 HepG2細胞增殖的能力比 ATRA更強。
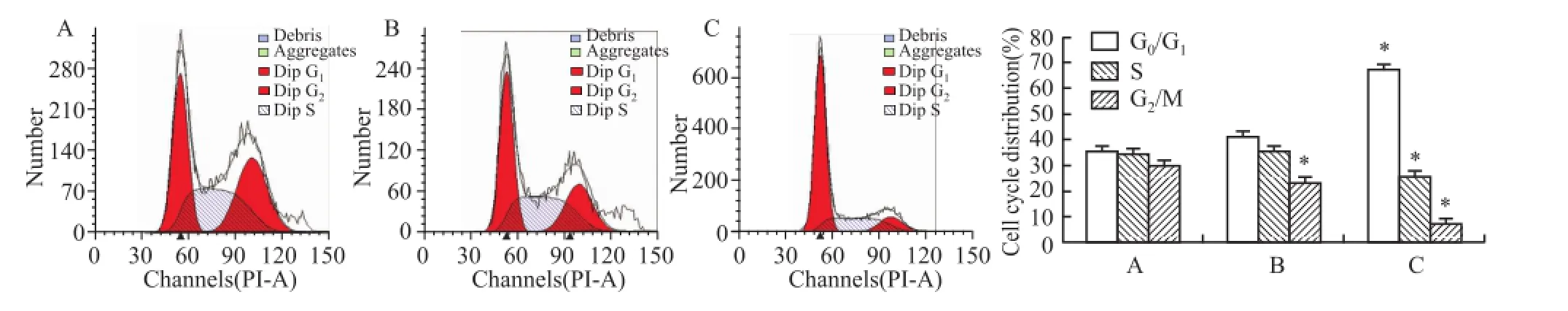
圖5 ATRA與 ATPR對肝癌 HepG2細胞周期的影響A:溶劑 對照組 ;B:ATRA 25μmol/L;C:ATPR 25μmol/L;與溶 劑對 照組 比較:*P<0.05
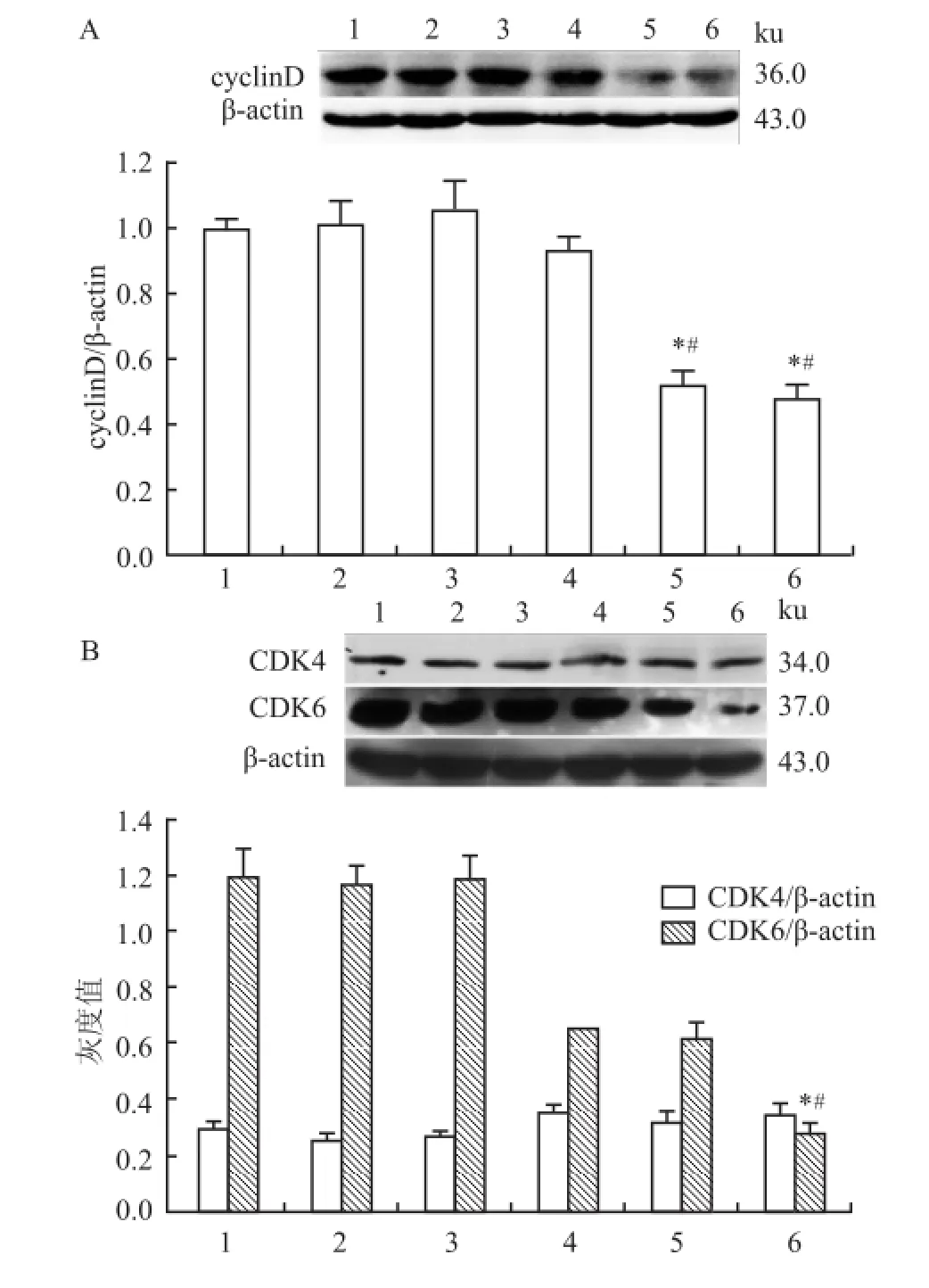
圖6 ATRA和 ATPR對 HepG2細胞 內 cyclinD、CDK 4和 CDK 6蛋白表達的影響1:細胞對照 組 ;2:溶 劑 對 照 組 ;3:ATRA 5μmol/L;4:ATRA 25 μmol/L;5:ATPR 5μmol/L;6:ATPR 25μmol/L;A:ATRA和 ATPR對HepG2細胞內 cyclinD蛋白的影響;B:ATRA和 ATPR對 HepG2細胞內 CDK4和 CDK6蛋白表達的影響;與細胞對照組比較:*P<0.05;與相同濃度的 ATRA組比較:#P<0.05
細胞周期檢測點是細胞增殖調控的關鍵位點,包括細胞周期蛋白 cyclin和細胞周期蛋白依賴性激酶 CDK的調控系統[11]。流式細胞術檢測細胞周期發現,ATPR可以顯著導致 HepG2細胞阻滯在 G0/G1期,抑制細胞增殖,這與MTT結果是一致的。CyclinD蛋白在細胞增殖過程中發揮重要作用,是調節 G1期細胞增殖信號的關鍵蛋白,其可與 CDK4或CDK6形成激酶復合物,使關鍵底物 PRb磷酸化,加快釋放轉錄因子 E2F,從而加速 G1/S期轉換[11-12]。細胞周期的失控會導致細胞惡性增殖,從而導致癌癥[11,13],因此,開發抗癌藥物來抑制細胞周期的進程顯得尤為重要。有研究[14]表明,在食管癌細胞株EC-9706和乳腺癌細胞株 MDA-MB-231中,通過抑制 cyclinD和 CDK4的水平,能夠誘導細胞 G0/G1期阻滯,從而抑制細胞增殖。因此,本研究檢測了 cyclinD、CDK4和 CDK6蛋白,結果顯示,經 ATPR處理48 h后的 HepG2細胞 cyclinD和 CDK6表達降低,而 CDK4蛋白水平恒定。因此,ATPR可能通過下調 cylinD和 CDK6蛋白水平來減少 cyclinD和 CDK6復合物的形成,抑制細胞周期 G1向 S期轉變,致使細胞阻滯在 G0/G1期,最終實現對 HepG2細胞的增殖抑制作用。
本實驗研究中,ATRA較其衍生物 ATPR對 cyclinD及 CDK6的抑制能力較弱,這可能與二者調節肝癌 HepG2細胞增殖方面存在不同的作用機制有關。研究[15]顯示 ATRA及其衍生物通過其核受體發揮抗癌作用,目前已發現兩種維甲酸受體家族:RARs和 RXRs家族,分別都有 α、β、γ3個亞型。ATRA與其衍生物通過與細胞內相應維甲酸受體結合后,特異性結合到靶基因調控區的維甲酸應答元件(RARE)上,選擇性地激活或抑制基因轉錄,調節細胞的增殖、分化和凋亡[15]。研究[5]證明 在胃癌BGC-823細胞中,ATPR較 ATRA能顯著下調 RARα和 RARβ受體,導致 ATPR抑制 BGC-823細胞遷移的能 力 更 強。因 此,ATRA 與 ATPR 調 節 肝 癌HepG2細胞增殖機制的不同可能與其進入 HepG2細胞內結合不同的維甲酸受體有關,二者對受體調節的研究也正在本實驗室進行。
綜上所述,在同等濃度下,新型維甲酸衍生物ATPR較 ATRA更能顯著抑制肝癌細胞株 HepG2的增殖。其機制可能是通過下調 cyclinD和 CDK6的表達水平阻滯 HepG2細胞周期進展,從而抑制細胞增殖,為肝癌的藥物治療提供了新的實驗依據。然而,細胞增殖是一個極其復雜且受嚴格調控的過程,ATPR如何下調 cyclinD和 CDK6的機制以及相關通路仍不清楚,有待后續研究進一步探討。
[1]Siddikuzzaman,Guruvayoorappan C,Berlin Grace V M.All trans retinoic acid and cancer[J].Immunopharmacol Immunotoxicol,2011,33(2):241-9.
[2]Saito A,Sugawara A,Uruno A,et al.All-trans retinoic acid induces in vitro angiogenesis via retinoic acid receptor:possible involvement of paracrine effects ofendogenous vascular endothelialgrowth factor signaling[J].Endocrinology,2007,148(3):1412-23.
[3]Su Y C,Dunn P,Shih L Y,et al.Retinoic acid syndrome in patients following the treatmentof acute promyelocytic leukemiawith all-trans retinoic acid[J].Chang Gung Med J,2009,32(5):535-42.
[4]Fan T T,Cheng Y,Wang Y F,et al.A novel all-trans retinoid acid derivative N-(3-trifluoromethyl-phenyl)-retinamide inhibits lung adenocarcinoma A549 cell migration through down-regulating expression ofmyosin light chain kinase[J].Asian Pac J Cancer Prev,2014,15(18):7687-92.
[5]Hu A,Yang Y,Zhang S,et al.4-Amino-2-trifluoromethyl-phenyl retinate inhibits themigration of BGC-823 human gastric cancer cells by downregulating the phosphorylation level of MLC II[J].Oncol Rep,2014,32(4):1473-80.
[6]周佳麗,顏蘊文,江巧玲,等.全反式維甲酸及其衍生物對乳腺癌細胞株 MDA-MB-231凋亡的影響[J].安徽醫科大學學報,2015,50(2):154-8.
[7]Koudah S,EIMouhadi S,ArrivéL.Fibrolamellar hepatocellular carcinoma[J].Clin Res Hepatol Gastroenterol,2012,36(1):5-6.
[8]Jemal A,Bray F,Center M M,et al.Global cancer statistics[J]. CA Cancer JClin,2011,61(2):69-90.
[9]Abrams P,Marsh JW.Current approach to hepatocellular carcinoma[J].Surg Clin North Am,2010,90(4):803-16.
[10]王 北,顏韻文,周 青,等.4-氨基-2-三氟甲基維甲酸酯對乳腺癌細胞株 MDA-MB-231遷移的影響及其可能機制[J].安徽醫科大學學報,2013,48(2):115-9.
[11]周春燕,馮作化.醫學分子生物學[M].北京:人民衛生出版社,2014:152-4.
[12]Okayama H.Cell cycle control by anchorage signaling[J].Cell Signal,2012,24(8):1599-609.
[13]Lange C A,Yee D.Killing the second messenger:targeting loss of cell cycle control in endocrine-resistant breast cancer[J].Endocr Relat Cancer,2011,18(4):C19-24.
[14]Wang H,Chen X,Chen Y,et al.Antitumor activity ofnovel chimeric peptides derived from cyclinD/CDK4 and the protein transduction domain 4[J].Amino Acids,2013,44(2):499-510.
[15]Soprano D R,Qin P,Soprano K J.Retinoic acid receptors and cancers[J].Annu Rev Nutr,2004,24:201-21.
Effects of a novel all-trans retinoid acid derivative 4-am ino-2-trifluoromethyl-phenyl retinate on the proliferation of HepG2 cells
Liu Hui,Zhou Qing,Wang Yuan
(Dept of Biochemistry and Laboratory of Molecular Biology,AnhuiMedical University;Dept of Key Laboratory,Gene Resource Utilization for Severe Disease of Anhui Province,Hefei 230032)
Objective To explore the effects of4-amino-2-trifluoromethyl-phenyl retinate(ATPR)on the proliferation of human hepatocellular carcinoma(HCC)HepG2 cells and preliminarily clarify the probable mechanisms. Methods HepG2 cells were used as the research model and treated with all-trans retinoic acid(ATRA)and ATPR respectively.The viability of HepG2 cells was investigated by MTT assay.Flow cytometry was used to detect cell cycle distribution.The expression of cyclinD,cyclin-dependent kinase 4(CDK4) and CDK6 proteins in HepG2 cellswere detected by Western blot.Results ATPR significantly inhibited the proliferation of HepG2 cells in a dose-and time-dependentmanner compared with cell group and arrested cells on G0/G1phase(P<0.05). The inhibition rate of ATPR wasmuch higher than that of ATRA at the same dose for the indicated time(P<0.05).In addition,Western blot results showed that ATPR down regulated the expression of cyclinD and CDK6 more dramatically than ATRA(P<0.05),while CDK4 expression was constant in all groups.Conclusion ATPR exhibits a better inhibitory effect on HepG2 cells proliferation than ATRA.The mechanism may be partly through down regulating the expression of cyclinD and CDK6 to cause G0/G1arrest,which finally leads to growth inhibition.
hepatocellular carcinoma;HepG2 cells;all-trans retinoic acid;4-amino-2-trifluoromethyl-phenyl retinate;cell cycle;cell proliferation
R 735.7;R 966;R 34
A
1000-1492(2016)02-0156-05
時間:2016-1-20 10:32:26
http://www.cnki.net/kcms/detail/34.1065.R.20160120.1032.004.htm l
2015-11-17接收
國家自然科學基金(編號:81272399);安徽省自然科學基金(編號:090413116)
安徽醫科大學分子生物學實驗室、生物化學與分子生物學
教研室、安徽省 /省部共建教育部重要遺傳病基因資源利用重點實驗室,合肥 230032
劉 慧,女,碩士研究生;
汪 淵,男,教授,博士生導師,責任作者,E-mail:aydesm-1@163.com
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
