高榕雌花期傳粉榕小蜂和欺騙性小蜂的繁殖特點
2016-05-10 02:33:20彭艷瓊楊大榮中國科學院西雙版納熱帶植物園昆明650222中國科學院大學北京00049西南林業大學云南生物多樣性研究院昆明650224
生態學報 2016年4期
徐 睿,張 媛,彭艷瓊,楊大榮中國科學院西雙版納熱帶植物園,昆明 650222中國科學院大學,北京 00049西南林業大學云南生物多樣性研究院,昆明 650224
?
高榕雌花期傳粉榕小蜂和欺騙性小蜂的繁殖特點
徐睿1,2,張媛3,彭艷瓊1,*,楊大榮1
1中國科學院西雙版納熱帶植物園,昆明650223
2中國科學院大學,北京100049
3西南林業大學云南生物多樣性研究院,昆明650224
摘要:榕樹及其專一性傳粉榕小蜂組成了動植物界最為經典的協同進化關系,傳粉榕小蜂演化出欺騙性是非常罕見的。在雌雄同株的高榕隱頭果內,共存著一種傳粉榕小蜂Eupristina altissima和一種欺騙性的小蜂Eupristina sp.,兩種小蜂在雌花期進入隱頭果內繁殖,但有不同的繁殖特點。對比研究了兩種小蜂從成蟲羽化到產卵和傳粉這個階段的雌蜂個體大小、孕卵量及繁殖差異,結果表明:羽化期兩種雌蜂的平均個體小,經飛行小個體的雌蜂易死亡,大個體雌蜂到達接受樹,但通過苞片通道,一些個體較大的傳粉榕小蜂被夾死導致進入果腔的雌蜂相對小,而欺騙性小蜂易通過苞片以至進入果腔的雌蜂個體較大。兩種未產卵雌蜂均表現為個體大者孕卵量較多,但兩種雌蜂的平均孕卵量沒有差異。即使有充足雌花資源產卵,兩種雌蜂均未產完所有卵,產卵后兩種雌蜂卵巢中的卵量均顯著減少,遺留下的卵量兩種小蜂間沒有差異。傳粉榕小蜂只有部分個體傳完所攜帶花粉,并表現為傳粉越成功的雌蜂,產卵越多。存在種內競爭時,兩種小蜂的產卵量均減少,傳粉榕小蜂的傳粉效率也降低。在種間競爭背景下,欺騙性小蜂產卵更成功,傳粉榕小蜂的產卵和傳粉量均受到極大抑制。研究結果說明雌花期隱頭果內傳粉榕小蜂只適量利用雌花資源產卵繁殖后代,更有效地傳粉繁殖榕樹種子,這可能是維持榕-蜂互惠系統穩定共存的重要機制之一;欺騙者穩定存在需降低與傳粉者的直接競爭,而欺騙者和傳粉者分散在不同果內,甚至是不同的樹上繁殖是理想的繁殖策略。
關鍵詞:榕樹;傳粉榕小蜂;互惠;欺騙者;繁殖差異
徐睿,張媛,彭艷瓊,楊大榮.高榕雌花期傳粉榕小蜂和欺騙性小蜂的繁殖特點.生態學報,2016,36(4):1134-1140.
Xu R,Zhang Y,Peng Y Q,Yang D R.Reproductive characteristics of pollinator and cheater wasps that utilize the female flowers of Ficus altissima.Acta Ecologica Sinica,2016,36(4):1134-1140.
榕樹依賴專一的榕小蜂傳粉實現有性繁殖,同時榕小蜂借助榕樹的雌花子房繁衍后代,兩者繁殖上相互依賴,形成了互不可缺的互惠共生體,最新分子證據顯示二者有約7500萬年協同進化的歷程[1-2]。榕樹開放的雌花能釋放特殊的化學揮發物,吸引專一的傳粉榕小蜂訪問[3],找到雌花期接受果后,榕小蜂便著陸于隱頭果表面,搜尋隱頭果頂部的苞片通道,鉆進層層苞片,到達果腔的小蜂往往是缺翅、斷觸角的,但它們仍將完成傳粉和產卵的使命。傳粉榕小蜂成蟲期壽命非常短,約1—2 d[4],離開隱頭果的傳粉榕小蜂必須盡快找到雌花期隱頭果,才能維持榕-蜂互惠系統的正常繁殖[5]。在雌雄同株的隱頭果內,一部分雌花傳粉后發育為種子,另一部分雌花子房被榕小蜂產卵后繁殖小蜂后代,種子及傳粉榕小蜂在同一果內繁殖[6]。關于雌花資源如何在傳粉和產卵之間進行平衡分配,主要有3種假說:短產卵器假說、不可摧毀種子假說及榕小蜂產卵不飽和假說[7-8],但隨著研究的深入,這3種假說均遭到質疑[9]。此外,果腔內傳粉榕小蜂完成傳粉和產卵是否有先后順序也一直是懸而未決的問題。通過雌花期不同時間段引入傳粉榕小蜂,間接顯示傳粉和產卵權衡與花開放的時間及傳粉榕小蜂雌蜂壽命均有關系[10]。
高榕隱頭果內雄花較少,成熟花粉囊不會自動破裂,為典型主動授粉模式的榕樹。其傳粉榕小蜂Eupristina altissima成蟲羽化、交配,雌蜂離開癭花進入果腔后,主動尋找雄花,并用觸角第3節上的一個勾刺破花粉囊,然后用前足基節上的花粉刷收集花粉,并裝入位于中胸腹面的囊狀花粉筐中;當雌蜂飛離羽化的榕樹找尋到新的雌花期高榕后,很快從頂生苞片進入雌花期果的果腔,又用花粉刷把花粉從花粉筐中刷出,為榕樹主動傳粉。此外,高榕隱頭果內還共存著另一種Eupristina sp.小蜂,該蜂花粉刷消失,花粉筐退化,沒有主動尋找雄花、收集花粉和傳粉的行為;由于蟲體很難粘附到花粉,該種小蜂已演化為只產卵不傳粉的欺騙性小蜂[11]。高榕隱頭果雌花期,傳粉者和欺騙者同時被吸引,在進入頂生苞片通道的過程中,存在強烈的種內打斗行為,但種間和平相處;兩種蜂繁殖季節差異明顯,并偏愛進入不同的隱頭果繁殖[12];單蜂引入雌花期隱頭果,欺騙性小蜂繁殖的后代更少[13]。在前期研究的基礎上,本文將深入對比研究高榕隱頭果內傳粉榕小蜂和欺騙性小蜂的產卵、繁殖差異。回答如下問題:(1)是否個體大的雌蜂更有機會獲得繁殖成功?(2)個體大的雌蜂是否孕卵量較多?(3)雌花資源充足時,雌蜂是否產下所有卵?(4)雌花期兩種小蜂單獨存在和共存時,產卵和傳粉的差異?
1 材料與方法
1.1材料與樣地
研究材料:高榕(Ficus altissima Blume),雌雄同株,隸屬于榕亞屬(Urostigma)、榕組(Section Urostigma)、環紋榕亞組(Subsection Conosycea)。高榕分布于亞洲熱帶大陸,在西雙版納熱帶地區,該種自然分布于熱帶雨林中,作為綠化樹或“神樹”也被種植于城市、鄉村及廟宇旁,具獨木成林現象。高榕葉腋結果,通常發新葉、結新果同時出現,單株樹一年可結果多次,種群內常年有樹結果;結果時樹內花期同步,樹間花期異步。高榕依賴專一的傳粉榕小蜂Eupristina altissima Balakrishnan&Abdurahiman傳粉獲得有性繁殖;在高榕隱頭果內,還共存著另一種Eupristina sp.小蜂,該種蜂原本是一種傳粉榕小蜂,但是其前足基節上的花粉刷消失,胸部的花粉筐退化,并且丟失主動傳粉行為,在高榕隱頭果內只產卵不傳粉,演化為一種欺騙性小蜂。高榕隱頭果內的欺騙者主要發生于高溫多雨的夏季,與傳粉者共存同一果的比率較低,約9%[11-12]。除了Eupristina屬兩種榕小蜂外,高榕隱頭果內還寄生著23種非傳粉小蜂,是榕小蜂群落最為豐富的一種榕樹[14]。
研究樣地位于云南省西雙版納傣族自治州景洪市(21°58'36.10″—22°00'25.59″N,100°45'51.95″—100°47'40.36″E,海拔552 m)。西雙版納地處東南亞熱帶北緣,屬于熱帶季風型氣候,年平均氣溫21.8℃,相對濕度86%,全年的降水量1556.9 mm,干濕季分明,降雨主要集中在雨季(6—10月)。旱季又可分為霧涼季和干熱季,霧涼季從11月到次年的2月,干熱季是每年的3—5月。
1.2研究方法
分別選擇一株寄生著傳粉者或欺騙者的接近雄花期(即:榕小蜂快羽化的榕果)的高榕,每樹采摘20個隱頭果,單果分裝在120目的紗網隔離袋內,讓果內榕小蜂自然羽化進入袋中,從中挑選有多個傳粉者或欺騙者羽化的隱頭果,每果隨機抽取6只傳粉者或6只欺騙者雌蜂,借助解剖鏡(Leica S8AP0),首先確定種類是傳粉者還是欺騙者,再通過目鏡內安裝的測微尺,測量雌蜂的頭寬,因為榕小蜂頭寬是代表其體大小的一個指標[15]。然后在解剖鏡下解剖榕小蜂腹部,整體拉出榕小蜂的卵巢,并分散卵巢中左右兩束卵[9],計數兩種小蜂卵巢中總孕卵量,傳粉者和欺騙者各測量、共計數30只雌蜂。
選擇一株榕果發育到雌花接受期,同時有傳粉者和欺騙者到達的高榕,在雌花期果周圍的樹枝上懸掛粘蟲板,收集訪問雌花期果的傳粉者和欺騙者。從粘蟲板上各取30只傳粉者和欺騙者雌蜂,用上述方法在解剖鏡下,鑒定榕小蜂種類、測量頭寬,并解剖統計卵巢內孕卵量。
在榕果發育到雌花期的樹上,觀察傳粉者和欺騙者進入接受期隱頭果的情況,榕小蜂進入的果將作上標記,待到第2天,絕大多數榕小蜂已完成傳粉或產卵,采集這些有榕小蜂進入的隱頭果,帶回實驗室,解剖隱頭果,專門挑選果腔內進入1只傳粉者、2只傳粉者、1只欺騙者、2只欺騙者,以及1只傳粉者和1只欺騙者的隱頭果,而且需要果腔內的小蜂處于奄奄一息,接近死亡的狀態,這樣能保證榕小蜂已傳粉、產卵完成,其次,卵巢中的卵能散開計數。每只雌蜂從果腔內取出,置于載玻片上,直接在解剖鏡下測量了頭寬、再檢查胸部花粉筐里是否有花粉[16],之后,用滴管加一滴75%的酒精解剖腹部,統計卵巢中左右兩束卵剩下的卵量。最終,獲得單果進1只傳粉者、單果進2只傳粉者、單果進1只欺騙者、單果進2只欺騙者以及單果進1只傳粉者和1只欺騙者的隱頭果,樣本量分別為30、8、30、26、9果。
1.3數據分析
在比較兩組數據的平均數時,若數據呈正態分布,選用獨立樣本T檢驗,若數據不呈正態分布則選用非參檢驗中的Mann-Whitney U測驗來分析;采用獨立樣本T檢驗進行如下數據分析:雌蜂羽化及到達雌花期接受樹兩個階段,傳粉榕小蜂和欺騙性小蜂個體大小差異;未產卵時,傳粉者和欺騙者雌蜂平均孕卵量的差異;傳粉榕小蜂產卵前后卵巢中遺留卵量的差異;競爭產卵后,傳粉者和欺騙者遺留卵量的差異;以及傳粉完全與否傳峰榕小蜂遺留卵量的差異。采用Mann-Whitney U測驗比較進入果腔內傳粉者和欺騙者的個體差異;以及比較欺騙者產卵前后卵巢中遺留卵量的差異。采用單因素方差分析(ANOVA)比較蜂羽化、到達接受樹及進入果腔3個階段,傳粉者和欺騙者個體的變異,并用Tamhane's T2(方差不齊)對欺騙性小蜂進行多重比較。此外,采用Pearson相關分析比較兩種榕小蜂頭寬與孕卵量的相關關系。所有的數據分析采用SPSS16.0統計軟件完成。
2 研究結果
2.1兩種榕小蜂的個體大小比較
用榕小蜂的頭寬代表其個體大小,分別比較了共存于高榕隱頭果內的傳粉榕小蜂和欺騙性小蜂在羽化、到達接受樹及進入果腔3個階段雌蜂個體的大小。結果顯示:剛羽化出來的傳粉者和欺騙者平均個體小;經過一段距離的飛行,小個體的雌蜂死亡,個體大的榕小蜂容易到達接受樹找到接受期隱頭果,然后雌蜂經過一個由苞片組成的通道進入雌花期果。經過苞片通道,一些個體較大的傳粉榕小蜂被夾死導致進入果腔的雌蜂個體相對較小,但3個階段傳粉榕小蜂平均個體大小之間的變異不顯著(F2,107=1.09,P=0.34)。欺騙性小蜂個體變異則表現為:剛羽化的雌蜂平均個體小,到達接受樹的雌蜂個體稍大,變異未達顯著水平(P=0.99),進入果腔內的雌蜂個體也較大,顯著大于剛羽化雌蜂個體的平均值(P<0.01),但與到達接受樹的雌蜂個體差異不顯著(P=0.15)。當比較種間個體大小時,羽化階段兩種雌蜂個體沒有差異(t=1.81,df=58,P= 0.08),到達階段,傳粉者顯著大于欺騙者(t=3.84,df=58,P<0.001),而進入果腔后欺騙者又比傳粉者顯著大(Z=-2.67,P<0.01)(圖1)。這說明:這兩種小蜂穿過苞片通道的能力有差異。

圖1 羽化、到達接受樹及進入果腔的兩種雌蜂個體大小(平均數±標準誤差)Fig.1Body sizes of female wasps that newly emerged,arrived at receptive fig trees and entered the cavity of figs for two wasp species(Mean±SE)
2.2兩種榕小蜂個體大小與孕卵量的關系
高榕傳粉榕小蜂平均頭寬為(0.451±0.003)(SE,n=60)mm,未產卵雌蜂的卵巢中有左右對稱的兩束卵,兩束卵量幾乎相等(左:(108.40±1.39),右:(108.55±1.23)),單蜂平均孕卵總量(215.12±2.94)(SE,n= 60)粒。欺騙性小蜂平均頭寬為(0.435±0.002)(SE,n=60)mm,同樣卵巢中左右兩束卵量也近似相等(左: (106.43±1.42),右:(107.45±1.34)),單蜂平均孕卵總量(213.72±2.71)(SE,n=60)粒,總孕卵量與傳粉者無差異(t=-0.35,df=118,P=0.73)。無論傳粉榕小蜂還是欺騙性小蜂,其雌蜂的個體大小均與孕卵量呈顯著正相關關系(傳粉者:r=0.33,P<0.05;欺騙者:r=0.50,P<0.001),即個體大的雌蜂孕卵量較多(圖2)。

圖2 兩種榕小蜂個體大小與孕卵量的關系Fig.2 The relationship between wasp body sizes and egg loads for two wasp species
2.3產卵前后兩種榕小蜂卵巢中卵量
通過解剖雌蜂卵巢統計卵數量,結果表明:到達接受樹的雌蜂,傳粉榕小蜂的平均孕卵量為(217.10± 3.88)(SE,n=30)粒,欺騙性小蜂的平均孕卵量為(211.87±4.20)(SE,n=30)粒,未產卵時兩種榕小蜂的孕卵量無差異(t=0.92,df=58,P=0.36)。進入接受果的傳粉榕小蜂將傳粉雌花,并產卵于雌花子房,而進入接受果的欺騙性小蜂只在雌花上產卵,并不進行傳粉。兩種榕小蜂產卵完成后,卵巢中仍然遺留一些卵,傳粉榕小蜂平均遺留卵數量(73.30±9.96)(SE,n= 30)粒,欺騙性小蜂平均遺留卵數量(69.47±8.78) (SE,n=30)粒,兩種榕小蜂遺留在卵巢中的卵數量沒有差異(t=0.29,df=58,P=0.77)。然而,與到達接受樹未產卵的雌蜂相比,兩種小蜂卵巢中的卵均顯著減少(傳粉者:Z=-6.57,P<0.001;欺騙者:Z=-6.65,P<0.001)(圖3)。
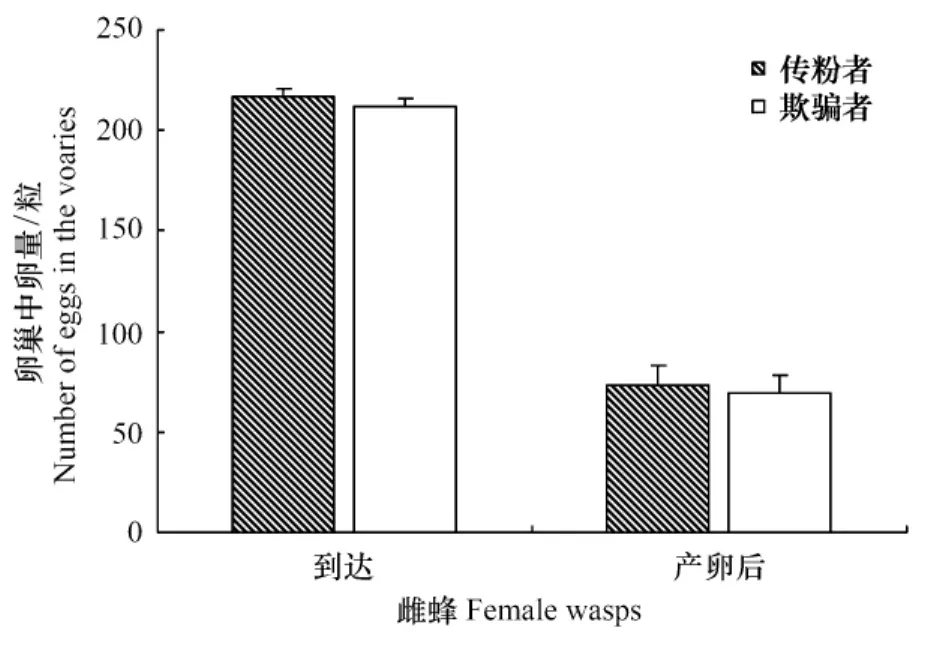
圖3 兩種榕小蜂產卵前后卵巢中卵數量(平均數±標準誤差)Fig.3The numbers of eggs in the ovary of female wasps before and after oviposition for two wasp species(Mean±SE)
2.4兩種榕小蜂單獨進入果時產卵和傳粉的情況
當隱頭果內只進入一只傳粉榕小蜂,完成產卵時卵巢中遺留卵量為(73.30±9.96)(SE,n=30)粒,同時傳粉完成后,只有67%的雌蜂用完了所攜帶的花粉,其余雌蜂的花粉筐中或多或少還留有一些花粉。這些傳粉不完全的小蜂,其對應卵巢里遺留的卵量也多,平均有卵(103.80±17.38)(SE,n=10)粒,而傳粉徹底的小蜂,其對應卵巢里遺留的卵量平均(58.05±10.53)(SE,n=20)粒,比前者顯著減少(t=2.32,df=28,P<0.05)。這說明:傳粉成功的榕小蜂,產卵也成功。當隱頭果內進入兩只傳粉榕小蜂時,產卵和傳粉效率均降低,產卵后卵巢遺留的卵量增加了0.5倍,傳粉后帶粉小蜂比率增加了近1倍,說明一果內進2只以上傳粉榕小蜂時,在產卵和傳粉的過程中,均相互受到干擾與影響。雖然欺騙性小蜂并不攜帶花粉,但一只蜂和兩只蜂產卵后,卵巢中遺留的卵量與傳粉榕小蜂的相當,同樣隱頭果進入兩只欺騙性小蜂時,產卵效率降低,卵巢中遺留卵量比進一只蜂時增加了0.6倍。無論隱頭果內進入一只還是兩只傳粉者或欺騙者,產卵后遺留在卵巢中左右兩卵束的卵量近似相等,這說明榕小蜂產卵時,左右對稱的兩束卵是等量釋放、產卵的(表1)。
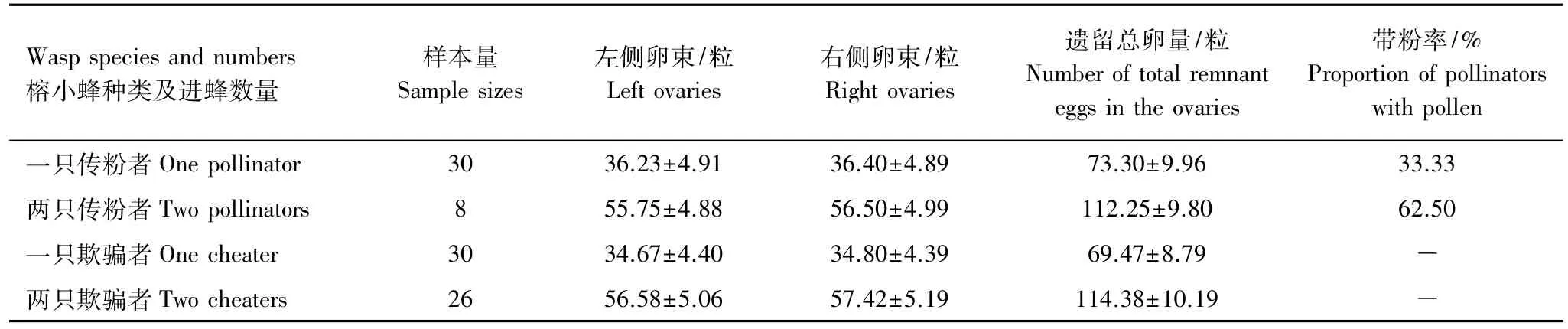
表1 單種榕小蜂在果腔內產卵和傳粉情況Table1 The oviposition and pollination status of wasps in fig entry by only one wasp species
2.5兩種榕小蜂進入同一果時產卵和傳粉的情況
當隱頭果內同時進入一只傳粉榕小蜂和一只欺騙性小蜂時,傳粉榕小蜂完成傳粉時,有44%的雌蜂用完所攜帶花粉,其余56%的雌蜂花粉筐中仍殘留部分花粉;產卵完成時,卵巢中還殘留有(102.00±12.54)(SE,n=9)粒卵。相比之下,欺騙性小蜂在果內只產卵不傳粉,產卵后卵巢遺留的卵量僅(40.22±10.23)粒,比傳粉榕小蜂遺留卵量顯著少(t=-3.82,df=16,P<0.05),即兩種小蜂在同一果內競爭產卵時,欺騙性小蜂產卵占據優勢(圖4)。
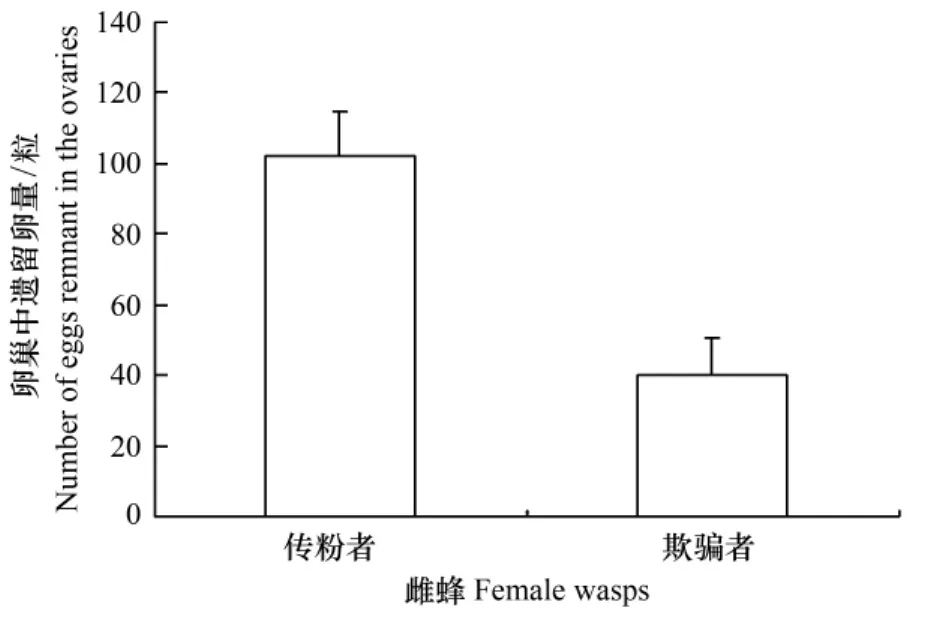
圖4 兩種小蜂競爭產卵時卵巢中遺留卵量(平均數±標準誤差)Fig.4 Number of remnant eggs in the ovaries for two wasp species in figs entered by both pollinators and cheaters(Mean± SE)
3 討論
本研究比較了高榕隱頭果內傳粉榕小蜂和欺騙性小蜂在羽化、到達接受樹及進入果腔內雌蜂的個體大小,兩種小蜂表現為個體大的雌蜂更有機會到達接受樹,但通過頂生苞片通道的篩選,個體較大的傳粉榕小蜂不易進入果腔,這與前人的研究結果相似[15,17];而欺騙性小蜂則表現有差異,進入果腔的雌蜂個體最大,這說明苞片通道的過濾作用對傳粉者和欺騙者是有差異的,欺騙者更容易通過。其次,比較了傳粉榕小蜂和欺騙性小蜂未產卵時孕卵量與個體大小的關系,兩種小蜂均呈現出:個體越大的小蜂孕卵量越多的正相關關系。進一步再比較兩種小蜂產卵前后,卵巢中卵量的差異,發現單只蜂在一個雌花果內產卵,有充足的雌花資源,但產卵后,榕小蜂卵巢中都遺留著一些卵,說明榕小蜂并未產下所有卵。當傳粉榕小蜂和欺騙性小蜂單獨進入雌花期果時,進兩只蜂的產卵和傳粉效率均比進一只蜂的果低,這是種內競爭產卵位點、食物或空間導致的。當雌花期果內進入一只傳粉者和一只欺騙者時,欺騙性小蜂產下的卵較多,傳粉榕小蜂產卵及傳粉效率遭受極大抑制,這表明:存在種間產卵競爭時,欺騙性小蜂更具有繁殖優勢。這些研究結果揭示了高榕傳粉榕小蜂和欺騙性小蜂之間一些尚未報道的繁殖特點和差異。
傳粉榕小蜂通常需要在其短暫的成蟲壽命期(通常1d)從它的出生果飛出、尋找到其他正在處于雌花期的同種隱頭果[18],飛行將消耗能量,只有那些個體較大的雌蜂能飛行到達接受樹,由于樹間飛行距離不同,到達接受樹的小蜂個體樹間有顯著差異,需長距離飛行的小蜂可能個體變異更大[15]。到達接受樹的小蜂必須通過頂生苞片通道才能到達果腔,接觸雌花傳粉和產卵,而榕樹隱頭花序的苞片結構有螺旋型和線型的結構,不同種類苞片的形狀、數量和排列交叉程度也不相似[19]。苞片較多、螺旋排列、重疊緊密的種類,榕小蜂進入頂生苞片很容易被夾死[20],而苞片少結構松散的種類,進入果腔的榕小蜂還能再出來[21],苞片通道結構與傳粉榕小蜂的頭部形態適應性演化,已相當于鎖與鑰匙的關系[22]。在本研究中,進入高榕隱頭果內的小蜂未見再出來現象,并且苞片對傳粉榕小蜂和欺騙性小蜂的篩選過濾作用有差異,個體大的傳粉榕小蜂有更多機會到達接受樹,但不容易進入果腔;而欺騙性小蜂則是到達果腔內的雌蜂個體最大,這一差異可能與兩種小蜂的頭寬有關,欺騙者頭部窄,進入苞片通道受到的篩選壓力可能較小。
傳粉榕小蜂成蟲壽命較短,進入到雌花期果內傳粉和產卵的時間約1天,已知傳粉榕小蜂在長花柱雌花上傳粉,短花柱雌花上產卵,但傳粉和產卵的先后順序目前仍不清楚[7]。本研究檢查了傳粉、產卵前后的傳粉榕小蜂,發現到達接受樹的傳粉榕小蜂100%攜帶著花粉,一只傳粉榕小蜂進果傳粉、產卵后,有66.67%的雌蜂傳完所攜帶的花粉,余下小蜂的花粉筐中或多或少遺留著一些花粉。而產卵后傳粉榕小蜂卵巢中遺留卵量,僅1只蜂(3%)產完所有的卵,且大部分小蜂遺留的卵量都較多;從這個對比數據可看出傳粉工作完成更快,而產卵工作可能由于過程復雜(從柱頭插入產卵器,卵產于子房中)導致小蜂不能產下更多的卵。研究結果還進一步揭示了傳粉越成功的榕小蜂,產卵越成功,導致的原因可能有兩種,一是個體大的傳粉小蜂帶粉量和孕卵量都多,因此繁殖更有效;其次是與果腔內傳粉榕小蜂的活力有關,健康活力強的榕小蜂傳粉和產卵更有效。一只欺騙性小蜂產卵量與傳粉小蜂相當,但還沒有發現把所有卵產完的個體。當同種的兩只小蜂進入同一果內繁殖時,由于種間競爭,兩種小蜂的產卵量下降了約0.5倍,而傳粉量減少了1倍,其原因可能是競爭空間和食物資源導致的。當雌花期果內同時進入一只傳粉者和一只欺騙者時,兩種小蜂競爭同樣的雌花資源,只產卵的欺騙者明顯占優勢,比傳粉者產下顯著多的卵;而傳粉榕小蜂不僅產卵量受限制,傳粉效率也降低,僅44%的雌蜂傳完所攜帶的花粉。這些對比研究結果說明,在高榕隱頭果內,傳粉榕小蜂和欺騙性小蜂產卵或干擾競爭種間大于種內,這與兩種小蜂在雌花期果壁上的打抖競爭不一致,在競爭進入苞片通道時,兩種小蜂的同種種內均存在激烈打斗,勝利者才能進入果腔,而種間沒有打斗行為,兩種小蜂在果壁上相遇,通常傳粉榕小蜂先行進入苞片通道,而欺騙性小蜂有膽怯、躲避傳粉榕小蜂的行為,待傳粉榕小蜂進入后才跟隨著進入[13]。這種不同階段種間及種內競爭上表現出來的差異可能與榕小蜂獲取繁殖資源的難易程度有關。
參考文獻(References):
[1]Wiebes J T.Co-evolution of figs and their insect pollinators.Annual Review of Ecology and Systematics,1979,10(1):1-12.
[2]Cruaud A,R?nsted N,Chantarasuwan B,Chou L S,Clement W L,Couloux A,Cousins B,Genson G,Harrison R D,Hanson P E,Hossaert-Mckey M,Jabbour-zahab R,Jousselin E,Kerdelhué C,Kjellberg F,Lopez-Vaamonde C,Peebles J,Peng Y Q,Pereira R A S,Schramm T,Ubaidillah R,van Noort S,Weiblen G D,Yang D R,Yodpinyanee A,Libeskind-Hadas R,Cook J M,Rasplus J Y,Savolainen V.An extreme case of plant-insect co-diversification:figs and fig-pollinating wasps.Systematic Biology,2012,61(6):1029-1047.
[3]Hossaert-McKey M,Soler C,Schatz B,Proffit M.Floral scents:their roles in nursery pollination mutualisms.Chemoecology,2010,20(2): 75-88.
[4]Dunn D W,Yu D W,Ridley J,Cook J M.Longevity,early emergence and body size in a pollinating fig wasp-implications for stability in a figpollinator mutualism.Journal of Animal Ecology,2008,77(5):927-935.
[5]Zhang Y,Yang D R,Peng Y Q,Compton S G.Costs of inflorescence longevity for an Asian fig tree and its pollinator.Evolutionary Ecology,2012,26(3):513-527.
[6]Wieblen G D.How to be a fig wasp.Annual Review of Entomology,2002,47:299-330.
[7]Nefdt R T,Compton S G.Regulation of seed and pollinator production in the fig-fig wasp mutualism.Journal of Animal Ecology,1996,65(2): 170-182.
[8]West S A,Herre E A.The ecology of the New World fig-parasitizing wasps Idarnes and implications for the evolution of the fig-pollinator mutualism.Proceedings of the Royal Society of London:Biological sciences,1994,258(1351):67-72.
[9]Dunn D W,Jansen-González S,Cook J M,Yu D W,Pereira R A S.Measuring the discrepancy between fecundity and lifetime reproductive success in a pollinating fig wasp.Entomologia Experimentalis et Applicata,2011,140(3):218-225。
[10]Zhang Y,Peng Y Q,Compton S G,Yang D R.Premature attraction of pollinators to inaccessible figs of Ficus altissima:A search for ecological and evolutionary consequences.PLoS One,2014,9(1):e86735.
[11]Peng Y Q,Duan Z B,Yang D R,Rasplus J Y.Co-occurrence of two Eupristina species on Ficus altissima in Xishuangbanna,SW China.Symbiosis,2008,45(1/3):9-14.
[12]Peng Y Q,Compton S G,Yang D R.The reproductive success of Ficus altissima and its pollinator in a strongly seasonal environment: Xishuangbanna,Southwestern China.Plant Ecology,2010,209(2):227-236.
[13]Zhao J B,Peng Y Q,Quinnel R J,Compton S G,Yang D R.A switch from mutualist to exploiter is reflected in smaller egg loads and increased larval mortalities in a‘cheater’fig wasp.Acta Oecologica,2014,57:51-57.
[14]段柱標,彭艷瓊,楊大榮,徐磊.高榕隱頭果內小蜂群落的動態變化.生態學報,2005,25(10):2589-2594.
[15]Liu C,Yang D R,Peng Y Q.Body size in a pollinating fig wasp and implications for stability in a fig-pollinator mutualism.Entomologia Experimentalis et Applicata,2011,138(3):249-255.
[16]Jandér K C,Herre E A.Host sanctions and pollinator cheating in the fig tree-fig wasp mutualism.Proceedings of the Royal Society B:Biological Sciences,2010,277(1687):1481-1488.
[17]Herre E A.Coevolution of reproductive characteristics in 12 species of New World figs and their pollinator wasps.Experientia,1989,45(7): 637-647.
[18]Harrison R D,Rasplus J Y.Dispersal of fig pollinators in Asian tropical rain forests.Journal of Tropical Ecology,2006,22(6):631-639.
[19]Verkerke W.Structure and function of the fig.Experientia,1989,45(7):612-622.
[20]Liu C,Yang D R,Compton S G,Peng Y Q.Larger fig wasps are more careful about which figs to enter-with good reason.PLoS One,2013,8(9): e74117.
[21]Suleman N,Raja S,Quinnell R J,Compton S G.Putting your eggs in several baskets:oviposition in a wasp that walks between several figs.Entomologia Experimentalis et Applicata,2013,149(1):85-93.
[22]van Noort S,Compton S G.1996.Convergent evolution of Agaonine and Sycoecine(Agaonidae,Chalcidoidea)head shape in response to the constraints of host fig morphology.Journal of Biogeography,1996,23(4):415-424.
Reproductive characteristics of pollinator and cheater wasps that utilize the female flowers of Ficus altissima
XU Rui1,2,ZHANG Yuan3,PENG Yanqiong1,*,YANG Darong1
1 Xishuangbanna Tropical Botanical Garden,Chinese Academy of Sciences,Kunming 650223,China
2 University of Chinese Academy of Sciences,Beijing 100049,China
3 Yunnan Academy of Biodiversity,Southwest Forestry University,Kunming 650224,China
Abstract:The interaction between pollinating fig wasps(Agaonidae)and their host fig trees(Ficus)is a striking example of obligate pollination mutualism.Ficus and its pollinating fig wasps rely on each other,and the coevolutionary relationship may date back to 75 Myr.Very rarely,the pollinating fig wasp evolves cheating in the fig-fig wasp mutualism,in which the pollinator loses pollination ability but still lays eggs in female flowers.Ficus altissima is a monoecious fig species in which seeds and wasps are produced in the same figs.The figs of F.altissima are occupied by two Eupristina species in the Xishuangbanna region:the pollinating fig wasp Eupristina altissima and the cheater Eupristina sp.The two species share similar biological characteristics but have different reproductive strategies.In this study,we compared the reproductive differences between the two wasp species during emergence,oviposition,and pollination.The results for both speciesbook=1135,ebook=247showed that newly emerging females were small on average,but some smaller females died during dispersal so that larger females arrived at receptive trees.When entering the figs,however,some larger pollinators were trapped in the ostiole so that the pollinators arriving in the fig cavity were smaller.The cheater,in contrast,easily passed through the ostiole so that the females arriving in the fig cavity were large.This suggests that the two species have different abilities to pass through the ostiole.In both E.altissima and Eupristina sp.females,larger wasps carried more eggs before oviposition,and there was no significant difference in average egg loads between the two species.When only one female was introduced into a fig,there were enough female flower resources for oviposition and pollination.However,E.altissima and Eupristina sp.did not lay all their eggs so some remained in the ovaries.The numbers of eggs in their ovaries were significantly reduced after oviposition,but the number of eggs remaining did not differ between the two species.Only some pollinating fig wasps used all the pollen grains they carried for pollination,and the wasps that pollinated more successfully,also laid more eggs.The pollinator performed pollination more quickly than oviposition.When two females of the same species were introduced to a fig,the numbers of eggs that both wasp species laid decreased,and the pollination efficiency of the pollinator was also reduced as a result of the intraspecific competition.When one pollinator and one cheater were introduced,the cheater laid eggs more successfully than the pollinator and both pollination and oviposition of the pollinator were restricted as a result of the interspecific competition.These results imply that the pollinating fig wasps only utilized part of the female resources for pollination and oviposition and that the efficiency of pollination was higher than that of oviposition.This could be a mechanism maintaining the fig-fig wasp mutualism.However,the cheater was a stronger competitor when it coexisted in the same figs with the pollinator,and significantly influenced seed and pollinator production.Therefore,for the cheater to stably coexist in the fig-fig wasp mutualism,it needs to reduce direct competition with the pollinator.A good reproductive strategy would be that the pollinator and cheater reproduce separately in different figs,or even in different trees.
Key Words:Ficus;pollinating fig wasps;mutualism;cheater;reproductive difference
*通訊作者
Corresponding author.E-mail:pengyq@xtbg.ac.cn
收稿日期:2014-06-25;網絡出版日期:2015-07-09
基金項目:國家自然科學基金項目(31372253,31120002);云南省中青年學術技術帶頭人后備人才培養項目(2011HB041)
DOI:10.5846/stxb201406251310
