中國特有瀕危植物白豆杉生長地的土壤性狀分析
2016-05-17 11:24:57王楨鄧琦蘇應(yīng)娟
生態(tài)科學 2016年5期
王楨, 鄧琦, 蘇應(yīng)娟,*
中國特有瀕危植物白豆杉生長地的土壤性狀分析
王楨1, 鄧琦2, 蘇應(yīng)娟2,*
1. 南京農(nóng)業(yè)大學生命科學學院, 南京 210095
2. 中山大學生命科學學院, 廣州 510275
白豆杉(Pseudotaxus chienii)是我國特有珍稀瀕危植物, 其生長地的土壤表現(xiàn)出明顯的異質(zhì)性。為此, 對分布在浙江龍門頂、廣西大明山、湖南張家界、江西三清山及福建武夷山的白豆杉種群的生長地進行了土壤取樣和成分分析。共測定了包括水分、pH值、有機質(zhì)、全N、P等在內(nèi)的19個土壤因子, 并對其展開主成分和聚類分析。結(jié)果顯示, 白豆杉生長地的土壤呈酸性; 除Zn和Si外, 其他因子在各個生長地的含量變異不大。確定pH值、有機質(zhì)、電導率、土壤新鮮水含量、土壤干燥水含量、K、Si和P為影響白豆杉生長的土壤因子。浙江龍門頂、福建武夷山、廣西大明山和湖南張家界的土壤相似性高, 而江西三清山的土壤與它們相差較遠。這些結(jié)果為進一步探究白豆杉對異質(zhì)性土壤的適應(yīng)性奠定了基礎(chǔ)。
白豆杉; 生長地的土壤; 性狀分析; 主成分分析; 聚類分析
1 前言
白豆杉(Pseudotaxus chienii (Cheng) Cheng)是我國特有珍稀瀕危植物, 為第三紀孑遺物種, 具典型的“古稀有種”特征[1]。它隸屬于裸子植物紅豆杉科(Taxaceae Gray)白豆杉屬(Pseudotaxus Cheng)[2]。白豆杉為常綠灌木或小喬木, 葉背面有兩條白色氣孔帶, 種子生于白色肉質(zhì)杯狀假種皮中[2–3]。該植物主產(chǎn)于廣東、廣西、湖南、江西及浙江[3]。因其長期呈星散分布, 個體稀少, 再加之雌株往往無法正常受粉, 故天然更新極其困難, 已被列為我國首批珍稀瀕危保護植物[4]。白豆杉一般生長在山地林下,棲息地的土壤表現(xiàn)出明顯的異質(zhì)性。白豆杉主要分布地的成土母質(zhì)各異, 其生長地的土壤一般屬山地黃壤, 肥力較高; 但在裸露程度、土層厚度及土壤成分含量迥異的懸崖峭壁和裸巖地段也有其植株生長[5]。
土壤條件是影響植物生長的關(guān)鍵因素[6]。土壤不僅能為植物生長提供必需的水、礦物質(zhì)和有機質(zhì),還能影響種子萌發(fā)、微氣候、排水模式以及局部水循環(huán)。植物從土壤吸收的金屬元素, 例如Cu和Zn,參與了包括光合作用、色素合成、蛋白質(zhì)代謝和維持生物膜完整在內(nèi)的多種生理生化過程[7]。此外, 土壤成分的組成還決定著植物對棲息地的偏好[8]。
迄今, 圍繞白豆杉開展的研究主要包括資源調(diào)查[5]、分類學[3,9]、分子系統(tǒng)學[10–11]、胚胎學[12]、地理分布和潛在分布區(qū)估計[13]、花粉形態(tài)[14]、種群結(jié)構(gòu)與群落特性[15]、篩分子質(zhì)體[16]、化學成分[17]、染色體核型[18]、同工酶[19]、種群遺傳分化[20]以及SSR引物的開發(fā)與利用[1]等方面。然而, 有關(guān)白豆杉種群生長地土壤成分的研究則還未見報道。
本研究針對白豆杉分布在浙江龍門頂、廣西大明山、江西三清山、湖南張家界和福建武夷山的天然種群, 對其生長地的土壤進行了取樣, 測定了土壤樣品的pH值與電導率以及水分、有機質(zhì)、全C、全N、P、S、Si和金屬元素的含量; 比較了不同生長地土壤成分的差異, 并借助主成分分析確定出對白豆杉生長起主導作用的土壤成分。在此基礎(chǔ)上,又通過聚類分析探討了白豆杉生長地土壤性質(zhì)間的相似性。
2 材料與方法
2.1實驗材料
2012年12月對白豆杉分布在浙江龍門頂、廣西大明山、湖南張家界和江西三清山以及2013年3月對福建武夷山的天然種群, 采集其生長地地表下方10—15 cm處的土壤樣品, 用封口膠袋保存(表1)。
2.2樣品分析方法
將新鮮土樣和通過0.2 mm尼龍篩的自然風干土樣105 ℃烘烤2 h, 分別測定土壤新鮮和風干后的水含量。土壤pH值和電導率以水土比5︰1震蕩混勻后, 用pH計和電導率測定儀測定。過0.2 mm篩后的土壤樣品, 加入K2Cr2O7和濃H2SO4, 利用稀釋熱法測定有機質(zhì)。土樣經(jīng)HNO3、HCl和HF微波消解后, 樣品送中山大學測試中心ICP室, 用電感耦合等離子體發(fā)射光譜儀(ICP-OES)測定消化液中P、S、Si、K、Ca、Na、Mg、Al、Fe、Mn、Zn和Cu元素濃度(mg·L-1)。將0.5000 g細篩土樣于定氮管中, 加入混合加速劑2.0 g (硫酸鉀:硫酸銅質(zhì)量比10︰1)和濃硫酸5 mL, 390℃加熱2 h, 冷卻, 放入全自動凱式定氮儀測定全N含量。采用高溫灼燒法測定全C含量。上述分析中, 每份土樣做三次平行重復(fù)測定。
2.3數(shù)據(jù)分析
采用NTSYS-pc 2.0進行主成分分析(Principal Component Analysis, PCA)。PCA是一種降維的方法,能將多個變量進行線性變換而選出較少的重要變量。通過計算土壤各成分的特征值和特征向量, 依據(jù)貢獻率, 確定關(guān)鍵性的主成分。另外, 還基于確定的主成分對取自不同生長地的土壤樣品進行了聚類分析。

表1 白豆杉種群的分布及土壤樣品Tab. 1 Distribution of Pseudotaxus chienii populations and soil samples
3 結(jié)果
3.1土壤pH值
5個白豆杉生長地土壤樣品的pH值介于3.21—4.31之間, 均低于5, 為強酸性土壤(表2)。其中, 張家界的土壤酸性最強(pH=3.21), 三清山的酸性較弱(pH=4.31)。
3.2土壤成分
在白豆杉種群的各生長地, 19個土壤因子呈現(xiàn)出一定程度的變異(表2)。土壤新鮮水的含量是福建武夷山最高(59.14%), 江西三清山最低(37.06%); 而土壤風干水的含量則是浙江龍門頂最高(11.36%),江西三清山最低(6.43%)。大明山土壤電導率最高(210.2 μS·cm-1), 龍門頂最低(173.4 μS·cm-1)。白豆杉種群生長地的有機質(zhì)、全C和全N含量較高。值得注意的是, 浙江龍門頂?shù)耐寥烙袡C質(zhì)、全C和全N含量最高(32.76%、36.66%和1.54%), 而福建武夷山的土壤有機質(zhì)含量最低(16.55%), 張家界全C含量最低(17.77%), 大明山土壤的全N含量最低(1.01%)。P含量方面, 張家界的土壤最高(0.831%),大明山的最低(0.397%)。江西三清山土壤的K含量較高(9.102 mg·g-1), 而浙江龍門頂土壤K含量較低(0.553 mg·g-1)。湖南張家界土壤Zn含量最高(1.137 mg·g-1), 而福建武夷山土壤的最低(0.028 mg·g-1)。此外, 武夷山的土壤的S含量也最低(0.970 mg·g-1)。
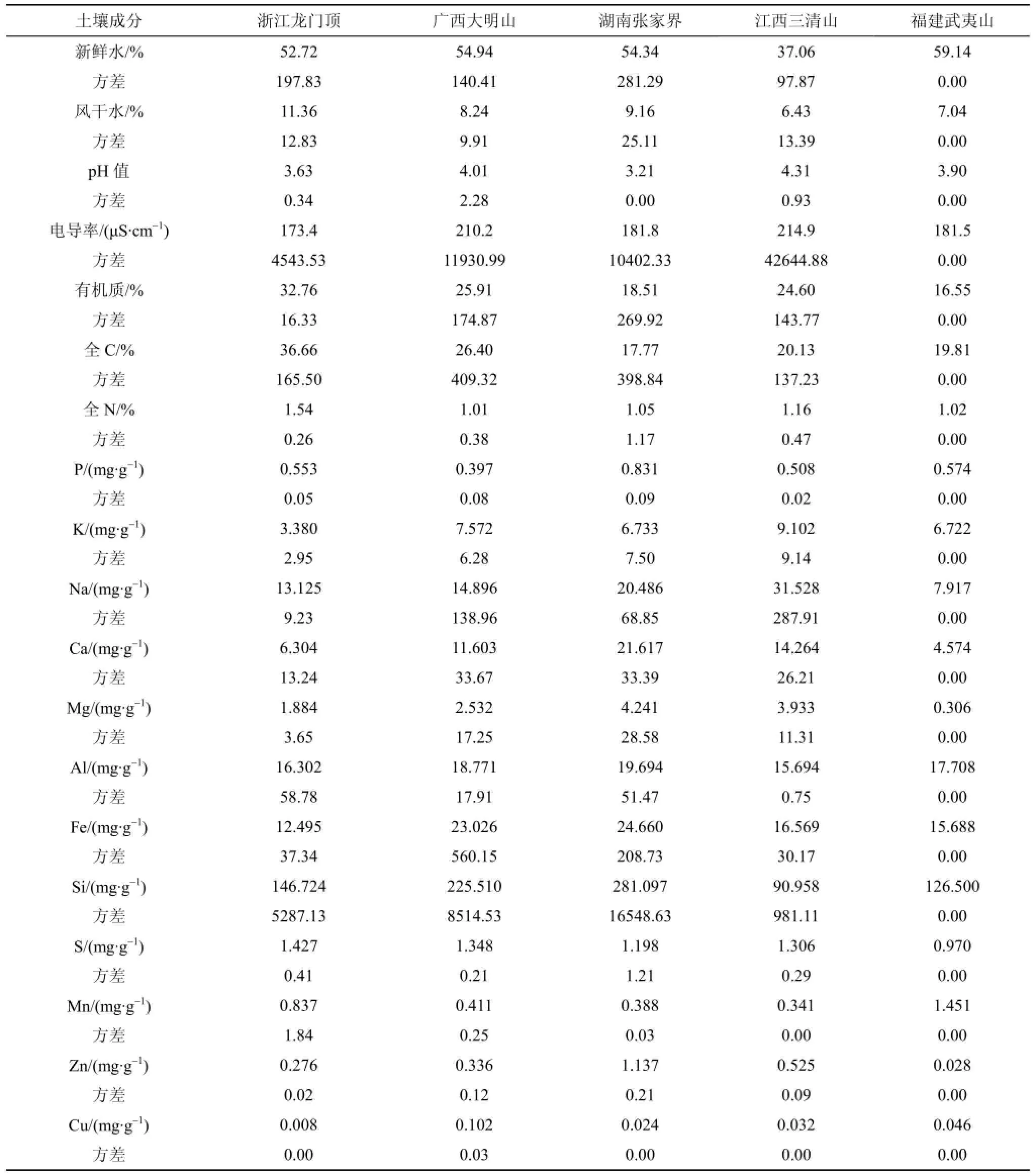
表2 白豆杉生長地土壤性狀Tab. 2 Soil characteristics in habitats of Pseudotaxus chienii
3.3主成分分析
通過PCA主成分分析, 確定了土壤因子主成分的特征值和特征向量(表3)。白豆杉生長地的3個主成分(特征值: 6.74+5.322+4.424=16.486個變量)能反映出全部19個因子86.8%的信息; 提示這3個主成分能反映土壤成分對白豆杉生長的影響。主成分1的方差貢獻率為35.5%, 電導率以及P、Si、K、Ca、Na、Mg、Al、Fe、Zn和Cu的主成分載荷較高; 主成分2的方差貢獻率為28.0%, 土壤新鮮水和干燥水以及全C、全N、P、Si、Ca、Al、Fe、Mn和Zn的主成分載荷較高, 尤以土壤干燥水為最高(0.715)。Mn和土壤新鮮水在第3主成分上有較高的載荷, 分別為0.712和0.386。根據(jù)主成分累計貢獻率, 計算土壤因子的貢獻率, 最終確定pH值、有機質(zhì)、電導率、土壤新鮮水含量、土壤干燥水含量、K、Si及P含量為影響白豆杉生長的重要土壤因子(表4)。
3.4聚類分析
基于主成分的聚類分析探究了白豆杉生長地彼此間在土壤性質(zhì)方面的相似性。浙江龍門頂和福建武夷山、廣西大明山和湖南張家界首先各自聚為一類, 顯示龍門頂和武夷山、大明山和張家界的土壤相似度較高。江西三清山與其他四者最后聚合, 提出其土壤性質(zhì)同其他生長地存在更大差異(圖1)。
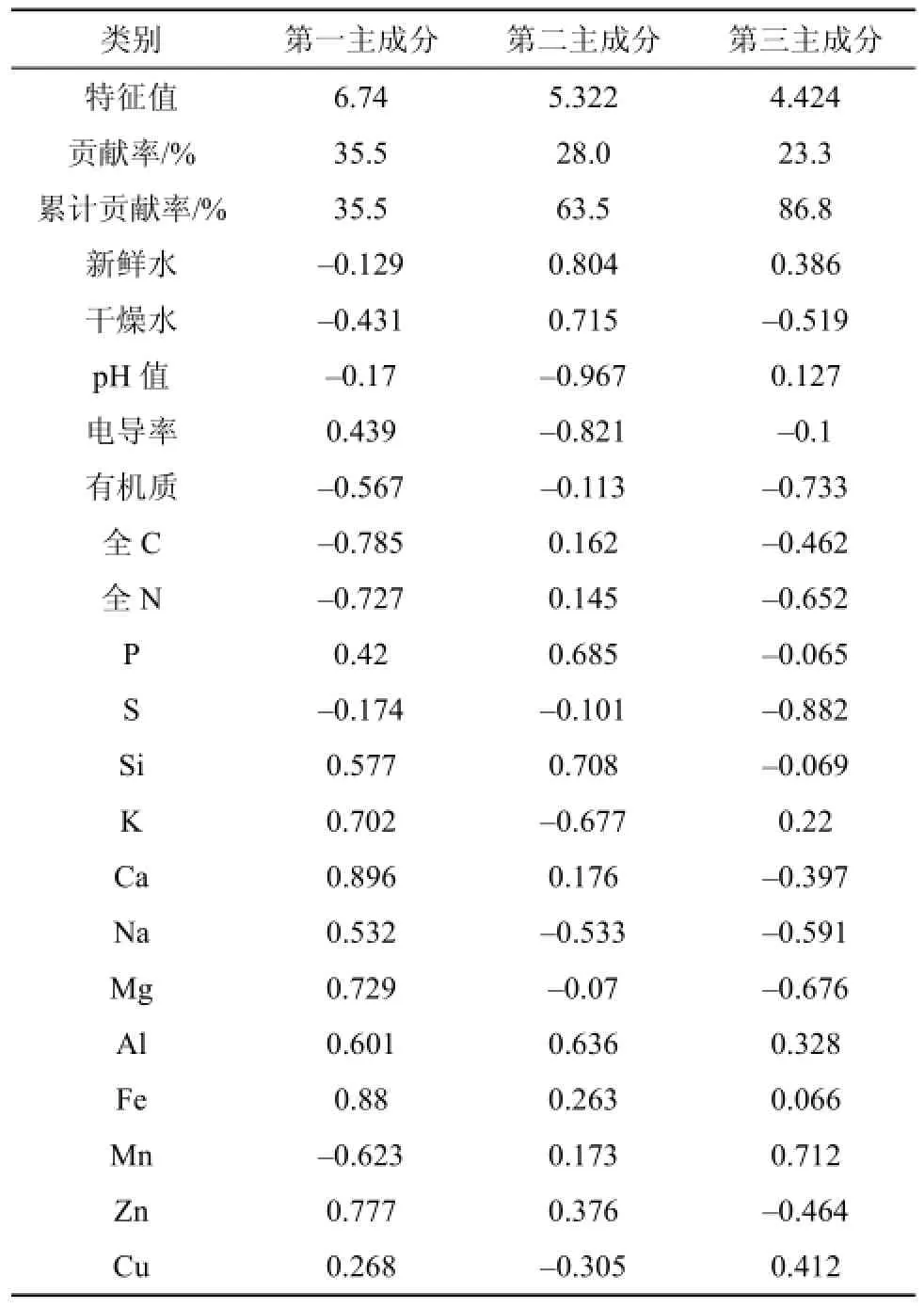
表3 白豆杉生長地19個土壤因子的主成分分析Tab. 3 Principal component analysis of 19 soil factors in habitats of Pseudotaxus chienii

表4 主成分分析中土壤因子的貢獻率Tab. 4 The contribution of soil factors to PCA analysis

圖1 基于主成分的白豆杉不同分布點的聚類圖Fig. 1 Dendrogram of different habitats for Pseudotaxus chienii based on principal components
4 討論
主成分分析顯示pH值、土壤新鮮水含量、土壤干燥水含量、有機質(zhì)、電導率、K、Si和P為影響白豆杉生長的關(guān)鍵土壤因子。白豆杉生長在酸性土壤, 生長地土壤的pH值均小于5。pH值的變化能影響Si和P的存在形式。在酸性條件下, Si和P分別以單硅酸(H4SiO4)和一價磷酸根(H2PO4-)的形式存在; 另外, 還會引發(fā)Al、Fe和土壤中的磷酸發(fā)生反應(yīng)。這些都會影響植株對Si和P的吸收, 進而改變它們在土壤中的含量。硅可通過改善氧化物酶(POD)、超氧化物歧化酶(SOD)及過氧化氫酶(CAT)的活性, 促使植物對氣候和土壤脅迫做出有效響應(yīng)[21]。磷是核苷酸和膜脂的組成成分, 涉及包括光合作用和呼吸作用在內(nèi)的多種重要代謝過程。我們的結(jié)果進一步提示: 硅和磷對于深入理解白豆杉種群的生長分布式樣和生理生態(tài)適應(yīng)具重要意義。
與其他裸子植物相比[22–23], 白豆杉生長地土壤的有機質(zhì)、全C和全N含量較高, 這與其土壤存在大量的腐殖質(zhì)和林下枯枝落葉較多有關(guān)。與之相對,生長地K的含量則相對偏低。此外, 還注意到, 白豆杉生長地土壤的Zn含量, 雖然其變異幅度高達40倍, 但卻不屬于PCA確定出的決定白豆杉生長的關(guān)鍵因子。K和Zn在保證酶的催化活性、維持胞內(nèi)離子平衡以及實現(xiàn)生物分子的高效合成等方面都起作用。然而, 在種群水平, 它們對白豆杉生長的相對貢獻卻可能存在較大差異。
白豆杉在浙江衢縣的分布最為集中, 面積最大、個體數(shù)量也最多, 龍門頂種群即位于該縣。該地的土壤特性表現(xiàn)為: 土層厚, 呈酸性; 有機質(zhì)含量高。然而, 白豆杉在其他省份的種群則相對稀疏[24]。本研究顯示, 浙江龍門頂和福建武夷山、廣西大明山及湖南張家界的土壤相似性高; 而江西三清山的土壤與其他幾個生長地相差較遠。這種差別體現(xiàn)在三清山的土壤Si含量偏低, 與含量最高的張家界土壤相差3倍。鑒于生長在異質(zhì)環(huán)境的白豆杉種群會隨環(huán)境的改變而發(fā)生遺傳分化[20]; 并且形態(tài)上, 白豆杉也呈現(xiàn)出漸變?nèi)禾卣鱗9]。我們建議, 在制定白豆杉遷地保育策略時, 應(yīng)充分考慮其天然種群生長地在土壤性質(zhì)上存在的差異, 優(yōu)先在龍門頂、武夷山、大明山及張家界種群間移栽個體。
5 結(jié)論
本研究得出以下主要結(jié)論: 1)白豆杉生長地的土壤呈酸性。除Zn和Si外, 土壤的水分、有機質(zhì)、P、全C、全N、S、K、Ca、Na、Mg、Al、Fe、Mn和Cu含量以及pH值和電導率在各生長地的變異不大; 2) pH值、有機質(zhì)、電導率及土壤新鮮水、干燥水、K、Si和P含量是影響白豆杉生長的關(guān)鍵土壤因子; 3)浙江龍門頂、福建武夷山、廣西大明山和湖南張家界的土壤相似性高, 而江西三清山的土壤與前四者相差較遠; 4)研究結(jié)果為白豆杉遷地保育策略的制定提供了依據(jù)。
致謝:感謝黃升對土壤樣品采集的大力支持,感謝中山大學林怡輝和孟繁博協(xié)助土壤成分分析,感謝中山大學測試分析中心提供的幫助。
[1] DENG Qi, SU Yingjuan, WANG Ting. Microsatellite loci for an old rare species,Pseudotaxus chienii, and transferability inTaxus wallichianavar.mairei(Taxaceae)[J]. Applications in Plant Sciences, 2013, 1(5): 1200456.
[2] 鄭萬鈞, 傅立國. 紅豆杉科, 中國植物志[M]. 北京: 科學出版社, 1978: 437.
[3] FU Liguo, LI Nan, MILL R R. Taxaceae, Flora of China[M]. Beijing: Science Press and St. Louis: Missouri Botanical Garden Press, 1999: 89–96.
[4] 傅立國, 金鑒明. 中國植物紅皮書—稀有瀕危植物(第一冊)[M]. 北京: 科學出版社, 1992.
[5] 胡紹慶, 陳征海, 孫孟軍. 浙江省白豆杉資源調(diào)查研究[J]. 浙江大學學報: 農(nóng)業(yè)與生命科學版, 2003, 29(1): 97–102.
[6] HANCOCK A M, WITONSKY D B, ALKORTAARANBURU G, et al. Adaptations to climate-mediated selective pressures in Humans[J]. PLoS Genetics, 2011,7(4): e1001375.
[7] YANG Qingwei, LAN Chongyu, SHU Wensheng. Copper and zinc in a paddy field and their potential ecological impacts affected by wastewater from a lead/zinc mine, P. R. China[J]. Environmental Monitoring and Assessment, 2008, 147(1–3): 65–73.
[8] GALLOWAY L F. Maternal effects provide phenotypic adaptation to local environmental conditions[J]. New Phytologist, 2005, 166(1): 93–100.
[9] 王康, 楊永. 紅豆杉科白豆杉屬植物的分類學研究[J].植物分類學報, 2007, 45(6): 862–869.
[10] 汪小全, 舒艷群. 紅豆杉科及三尖杉科的分子系統(tǒng)發(fā)育—兼論竹柏屬的系統(tǒng)位置[J]. 植物分類學報, 2000, 38(3): 201–210.
[11] CHENG Yuchang, NICOLSON R G, TRIPP K, et al. Phylogeny of Taxaceae and Cephalotaxaceae genera inferred from chloroplastmatK gene and nuclear rDNA ITS region[J]. Molecular Phylogenetics and Evolution, 2000, 14(3): 353–365.
[12] 陳祖鏗, 王伏雄. 白豆杉的胚胎發(fā)育及其系統(tǒng)位置的商榷[J]. 植物分類學報, 1978, 16(2): 1–10.
[13] 徐曉婷, 楊永, 王利松. 白豆杉的地理分布及潛在分布區(qū)估計[J]. 植物生態(tài)學報, 2008, 32(5): 1134–1145.
[14] 席以珍. 中國紅豆杉科花粉形態(tài)的研究[J]. 植物分類學報, 1986, 24(4): 247–252.
[15] 楊旭, 于明堅, 丁炳揚. 鳳陽山白豆杉種群結(jié)構(gòu)及群落特性的研究[J]. 應(yīng)用生態(tài)學報, 2005, 16(7): 1189–1194.
[16] 王艇, 蘇應(yīng)娟, 張宏達. 11種裸子植物篩分子質(zhì)體的超微結(jié)構(gòu)及分類[J]. 中山大學學報: 自然科學版, 1996, 35(5): 82–86.
[17] 馬忠武, 何關(guān)福, 印萬芬. 中國特有種子植物白豆杉主要化學成分的研究[J]. 植物學報, 1982, 24(6): 554–557.
[18] 管啟良, 林立. 白豆杉的核型和性染色體的研究[J]. 遺傳學報, 1993, 20(2): 155–158.
[19] 胡志昂, 王洪新, 劉長江, 等. 裸子植物的生化系統(tǒng)學(三)—從種子蛋白多肽和針葉過氧化物酶探討紅豆杉科的系統(tǒng)位置[J]. 植物分類學報, 1986, 24(4): 260–263.
[20] SU Yingjuan, WANG Ting, OUYANG Puyue. High genetic differentiation and variation as revealed by ISSR marker inPseudotaxus chienii(Taxaceae), an old rare conifer endemic to China[J]. Biochemical Systematics and Ecology, 2009, 37(5): 579–588.
[21] BALAKHNINA T, BORKOWSKA A. Effects of silicon on plant resistance to environmental stresses: review[J]. International Agrophysics, 2013, 27(2): 225–232.
[22] 普瓊, 薛會英. 西藏紅豆杉適生地土壤養(yǎng)分特性初步研究[J]. 西藏科技, 1999, 83(1): 20–21.
[23] 曾思齊, 劉發(fā)林. 6種經(jīng)營模式的木荷南方紅豆杉次生林土壤理化性質(zhì)[J]. 中南林業(yè)科技大學學報, 2014, 34(7): 9–18.
[24] 傅金堯, 胡紹慶. 浙江省白豆杉資源調(diào)查及植物區(qū)系研究[J]. 華東森林經(jīng)理, 2002, 16(2): 44–47.
Habitat soil properties of the natural population of Pseudotaxus chienii
WANG Zhen1, DENG Qi2, SU Yingjuan2,*
1.School of Life Sciences,Nanjing Agricultural University,Nanjing210095,China
2.School of Life Sciences,Sun Yat-sen University,Guangzhou510275,China
Pseudotaxus chienii(Cheng) Cheng (Taxaceae) is an endangered conifer endemic to China. Its soils in different habitats exhibit significant heterogeneity. In this study, 19 soil factors were measured for theP. chieniihabitats located in Longmending (Zhejiang), Damingshan (Guangxi), Zhangjiajie (Hunan), Sanqingshan (Jiangxi), and Wuyishan (Fujian). They included pH, electrical conductivity, and contents of fresh water, air-dried water, organic matter, total N, P, etc. A principal component analysis together with a cluster analysis was then conducted. The results showed that the soil was acidic. All the factors except Zn were little fluctuated in different habitats. Electrical conductivity, pH, and contents of fresh water, air-dried water, organic matters, K, Si, and P were revealed to be the principal soil factors that affected the growth ofP. chienii. Habitat soil was similar in Longmending, Wuyishan, Damingshan, and Zhangjiajie, and they showed higher similarity than with Sanqingshan. This study has provided valuable information for further exploring the adaptation ofP. chieniito heterogeneous soils.
Pseudotaxus chienii; habitat soil; soil characters; principal component analysis; cluster analysis
10.14108/j.cnki.1008-8873.2016.05.028
Q69
A
1008-8873(2016)05-208-06
王楨, 鄧琦, 蘇應(yīng)娟. 中國特有瀕危植物白豆杉生長地的土壤性狀分析[J]. 生態(tài)科學, 2016, 35(5): 208-213.
WANG Zhen, DENG Qi, SU Yingjuan. Habitat soil properties of the natural population ofPseudotaxus chienii[J]. Ecological Science, 2016, 35(5): 208-213.
2015-12-02;
2016-01-07
國家自然科學基金項目(31570652); 廣東省自然科學基金(2016A030313320)
王楨(1997—), 男, 山西晉城人, 在讀本科, 生物學, E-mail: 1257110928@qq.com
*通信作者: 蘇應(yīng)娟, 女, 博士, 教授, 主要從事植物進化遺傳學研究, E-mail: suyj@mail.sysu.edu.cn
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
