半紅樹植物黃槿的氣體交換特性
2016-05-27 01:45:19蔡水花周光霞陳貽竹葉萬輝
廣西植物 2016年4期
蔡水花, 李 婷, 周光霞, 陳貽竹, 韋 霄, 葉萬輝, 沈 浩*
( 1. 中國科學院退化生態系統植被恢復與管理重點實驗室, 中國科學院華南植物園, 廣州 510650;2. 中國科學院大學, 北京 100049; 3.廣西壯族自治區中國科學院廣西植物研究所, 廣西 桂林 541006 )
?
半紅樹植物黃槿的氣體交換特性
蔡水花1,2, 李婷1,2, 周光霞1,2, 陳貽竹1, 韋霄3, 葉萬輝1, 沈浩1*
( 1. 中國科學院退化生態系統植被恢復與管理重點實驗室, 中國科學院華南植物園, 廣州 510650;2. 中國科學院大學, 北京 100049; 3.廣西壯族自治區中國科學院廣西植物研究所, 廣西 桂林 541006 )
摘要:半紅樹植物是一類既可在海岸潮間帶又可在陸地生長和繁殖的海陸兩棲植物。夏季強光和高溫是海岸帶生境突出的特點,半紅樹植物如何在這樣的自然生境中生存和繁衍是值得探討的問題。該研究以具有較高應用價值的錦葵科(Malvaceae)木槿屬 (Hibiscus)的常綠半紅樹植物黃槿(H. tiliaceus)作為研究對象,采用LI-6400便攜式光合測定系統(Li-Cor Inc., USA)對南亞熱帶氣候條件下的廣東省珠海淇澳島紅樹林自然保護區自然生長的黃槿在夏季高溫季節的葉片氣體交換特性進行測定,探討了其在自然生境下的光合行為和相關的形態解剖特征,揭示其生境適應性的光合生理生態機制。結果表明:(1) 黃槿凈光合速率、氣孔導度、胞間二氧化碳濃度和蒸騰速率的日變化均表現為單峰型曲線,中午未出現明顯光抑制現象。(2) 黃槿較高的凈光合速率與其較高的氣孔導度相關,而后者則可能與其較大的氣孔密度和具有表皮毛等葉片解剖結構特征有關。(3) 黃槿光飽和點、光補償點、最大凈光合速率和日均凈光合速率均符合陽生植物特征,因而是一種陽生性樹種。以上結果說明黃槿在自然環境中具有的快速生長特性可能與其較強和較穩定的光合能力有關,該研究結果為深入揭示黃槿以及其他半紅樹植物適應海岸帶高溫強光生境的生理生態機制提供了基礎數據,為黃槿的合理栽培及造林實踐提供了理論參考。
關鍵詞:黃槿, 半紅樹, 氣體交換, 生境適應性
Gas exchange characteristics in the mangrove associateHibiscustiliaceus
CAI Shui-Hua1,2, LI Ting1,2, ZHOU Guang-Xia1,2, CHEN Yi-Zhu1,WEI Xiao3, YE Wan-Hui1, SHEN Hao1*
( 1.KeyLaboratoryofVegetationRestorationandManagementofDegradedEcosystems,SouthChinaBotanicalGarden,ChineseAcademyofSciences, Guangzhou 510650, China; 2.UniversityofChineseAcademyofSciences, Beijing 100049, China; 3.GuangxiInstituteofBotany,GuangxiZhuangAutonomousRegionandChineseAcademyofSciences, Guilin 541006, China )Abstract: Mangrove associates are a category of plants that can grow and reproduce on both intertidal coastal zones and land. As subtropical coastal zones are characterized by high sun light and temperature, how mangrove associates adapt to such an environment is an important issue. Some researches show that mangrove associates are similar to land plant as well as thalassophyte in certain morphological structures and biochemical characteristics. However, there are few studies focusing on plant physiological characteristics of mangrove associates, especially in leaf gas exchange characteristics.Hibiscustiliaceus(Malvaceae) is a pantropical mangrove associate that usually occurs in coastal zones. It is widely used for the conservation of sea banks, architecture and medicine. In order to disentangle the photosynthetic physiological mechanims underlying its habitat adaptation to high light and temperature under south subtropical climate, leaf gas exchange characteristics ofH.tiliaceuswere measured using LI-6400 portable photosynthesis system (Li-Cor Inc, USA) at Mangrove Natural Reserve of Qi’ao Island of Zhuhai City, Guangdong Province during July and August of 2013. The nature reserve (22°23′40″ N, 113°36′40″ E) is characterized by a typical south subtropical monsoon climate. The diurnal changes in photosynthetic rate (Pn), stomatal conductance (Gs), intercellular CO2concentration (Ci) and transpiration rate (E) inH.tiliaceusall exhibited a mono-peak modal, and the peak value ofPnoccurring at noon was around 26.8 μmol CO2·m-2·s-1without obvious photoinhibition during the daytime. The high photosynthetic rate inH.tiliaceuswas associated with high stomatal conductance, which was probably related to anatomical features of leaves, such as high stomatal density and dense trichomes in the lower leaf epidermis. Analysis of light response curves revealed that its light saturation point was 800 μmol photons·m-2·s-1and its light compensation point was 26 μmol photons·m-2·s-1. The net photosynthesis at saturating irradiances was high at 27.8 μmol CO2·m-2·s-1. These photosynthesis parameters, along with high daily mean photosynthetic rate (19.6 μmol CO2·m-2·s-1) inH.tiliaceussuggested that this mangrove associate was characterized as a typical sun plant species. The results showed that the fast growth ofH.tiliaceusmight be attributed to its high and stable photosynthetic capacity, thus providing basic data for further revealing the eco-physiological mechanism underlying habitat adaptation ofH.tiliaceusand other mangrove associates to coastal environment with high temperature and high light in summer. These results can also provide theoretical supports for its planting and reforestation practices.
Key words:Hibiscus tiliaceus, mangrove associate, gas exchange, habitat adaptation
紅樹植物是一類生長在熱帶、亞熱帶海岸潮間帶(最高潮線以下及平均高潮線以上)的喬木、灌木或草本植物,對海岸潮間帶的特殊生境(如高且多變的鹽濃度、缺氧、長時間的強光照和高的葉表面溫度)有很強的適應能力(Tomlinson,1986)。紅樹林生態系統覆蓋了世界上60%~75%的海岸線,是生產力水平最高的4個海洋生態系統之一。紅樹包括84個種,分屬16科24屬(Wang et al,2003),由于環境變化和陸地海洋演變,紅樹植物群落形成梯度和帶狀分布,造成了種的分化,這些種的許多生理和結構特征彼此不完全相同(Cheesman et al,2004)。紅樹植物通常分為真紅樹植物和半紅樹植物,前者只出現在河口潮間帶,具有為適應環境而演化出的氣生根及胎生現象等,以紅樹科(Rhizophoraceae)植物為代表;后者既能在潮間帶生長也能延伸到陸地生態系統,如行道樹常選用的黃槿(Hibiscustiliaceus)、海芒果(Cerberamanghas)等。
黃槿(Hibiscustiliaceus)為錦葵科(Malvaceae)木槿屬 (Hibiscus)常綠灌木或喬木,作為泛熱帶的半紅樹植物,在熱帶、亞熱帶的沿海堤岸和內陸均有分布,廣布于印度洋和西太平洋沿岸,以太平洋群島尤其多見(Tomlinson,1986;王伯蓀等,2003)。在我國產于臺灣地區、廣東、福建等省(中國植物志,1984)。黃槿具有較高的應用價值,木材可以制成獨木舟和工具(Tan,2001);樹皮中的粗纖維可加工成繩子;葉子和花具有藥用價值,葉子可止咳化痰,花可治療炎癥和化膿(Rosa et al, 2006);根系發達,可防風固沙。目前對黃槿的研究還相對較少,主要集中在形態結構、生理生化和遺傳結構方面,例如,黃槿葉面積大,枝角大,能充分利用空間資源,獲取更多的光照(張偉偉等,2012);黃槿葉片產生含低分子的親和溶質甲基化季胺,甜菜堿和醣類等滲透調節物(Parida & Jha,2010),提高滲透調節能力以跨越生長在鹽和淡水生境 (Nichol et al,2006);黃槿系統地理學特征和遺傳結構研究表明黃槿廣泛分布于熱帶地區可能是由于種子漂流習性的原因(Takayama et al,2006)。然而,關于黃槿生理生態學特征方面的研究不多,且大多局限于溫室幼苗的研究,對自然條件下生長的黃槿研究較少。當光合作用被基質鹽抑制時,黃槿能通過提高對光合組織的分配減緩鹽的負作用使得生長速率無凈差異(Santiago et al,2000);在干旱環境下,鹽度≥40%,一周內,黃槿全部死亡(Youssef,2007)。
海岸帶生境突出的特點是夏季強光和高溫,植物如何在這樣的自然生境中生長和繁殖,其內在光合生理生態機制是一個值得探討的問題。為此,本文試圖通過對南亞熱帶氣候條件下自然生長的黃槿進行葉片氣體交換特性的研究,旨在探討其在自然生境下的光合行為和相關的適應形態解剖特征,為其合理栽培及造林實踐提供理論參考。
1材料與方法
1.1 研究區概況
研究樣地位于廣東省珠海市淇澳島紅樹林自然保護區,地理位置為113°36′40″~113°39′15″ E,22°23′40″~22°27′38″ N,屬南亞熱帶海洋性季風氣候。區內年均氣溫22.2 ℃,7月平均氣溫27.9 ℃,極端最高氣溫35.0 ℃;1月平均氣溫為14.9 ℃,極端最低氣溫2.6 ℃,基本無霜期。年降水量為1 875.7 mm,4-10月為雨季,占全年降水量的84%,11月至翌年3月為旱季,占全年降水量的16%。保護區受降雨、江河徑流和潮汐的影響,海水鹽度在3.31‰~7.05‰變化,海域潮汐屬不正規半日潮。
1.2 實驗材料
于2013年7月下旬至8月上旬,在樣地內選取自然生長的健康、成熟黃槿植株4株,株高為2.5~3.0 m,胸徑4~6 cm,冠幅(1.5~2)m×(1.5~2)m。于植株冠層外側向陽方位選取葉位、大小和成熟度相近的健康成熟葉片,進行形態解剖特征和光合氣體交換指標的觀測。
1.3 觀測指標與方法
1.3.1 葉片解剖結構特征將葉片于4%戊二醛溶液中固定24 h以上(4 ℃),再經0.1 mol·L-1磷酸緩沖液處理,酒精梯度脫水(30%、50%、70%、80%、90%、100%),最后在叔丁醇中冷凍,JFD-310真空冷凍干燥儀干燥后,采用雙面碳膠帶固定在樣品臺上,JFC-1600型離子濺射儀(JEOL Ltd,Tokyo,Japan)鍍金膜,在JSM-6360LV型掃描電子顯微鏡(JEOL Ltd,Tokyo,Japan)下觀察和拍照。觀測指標為氣孔密度(每1 mm2內的氣孔個數)、葉片總厚度以及上、下表皮柵欄組織和海綿組織厚度。
1.3.2 氣體交換特性選擇晴朗天氣,使用LI-6400便攜式光合作用測定系統(LI-Cor Inc,Lincoln NE,USA) 進行黃槿植株葉片的光合光響應曲線和氣體交換日變化的活體測定。
(1)光響應曲線測定:在上午8:00-11:00進行,設定葉溫30 ℃,參比室CO2濃度360 μmol·mol-1,用連體葉片分別在以下光量子通度密度(photosynthetic photon flux density,PPFD)測定光合作用光響應動態為500、800、1000、1 500、1 800、2 000、200、100、50、20、0 μmol photons·m-2·s-1(內置6400-02B型紅藍光源提供)。設置最大和最小等待時間后,由儀器自動記錄數據。每次開始新的測量時,將待測葉片先在500 μmol photons·m-2·s-1下適應20 min,再開始測定。根據光響應曲線取得的數據,以非直角雙曲線(nonrectangular hyperbola)公式(Lambers et al,1998) 計算出表觀量子效率、暗呼吸速率、曲度系數、最大凈光合速率等氣體交換參數。
式中,Pn為凈光合速率(μmolCO2·m-2·s-1);I為光量子通度密度(PPFD,μmolphotons·m-2·s-1);Φ為光響應曲線起始斜率,即表觀量子效率(apparentquantumyield,molCO2·mol-1photons);Rd為暗呼吸速率(darkrespirationrate,μmol·m-2·s-1);Pmax為光飽和點時凈光合速率;θ為曲度系數(無量綱,0<θ<1)。利用葉片在PPFD≤200μmolphotons·m-2·s-1范圍內的PPFD(X軸)和Pn(Y軸)進行直線回歸,得出回歸直線,其與X軸的交點數值即為光補償點(LCP,μmolphotons·m-2·s-1)。將非直線雙曲線模型計算出的Pmax值和X軸作平行直線,兩條直線的交點在X軸上的數值則為光飽和點(LSP,μmolphotons·m-2·s-1)(Walker,1989)。
(2)氣體交換日變化測定:分別在8:00、9:00、10:00、11:00、12:00、13:00、14:00、16:00和18:00這9個時間點,在自然光下測定連體生長葉片的氣體交換指標及環境參數。在每個測定時間點均固定選取3~5片生長狀況和生長位置相似的葉片重復測定,取其平均值作為所測植株的指標,再以4株的平均值作為該物種的相應指標。測定項目包括葉片的凈光合速率、蒸騰速率(E,mmolH2O·m-2·s-1)、氣孔導度(Gs,molH2O·m-2·s-1)、胞間CO2濃度(Ci,μmolmol-1)等氣體交換指標以及PPFD、氣溫(Ta,℃)、葉溫(Tl,℃)和空氣相對濕度(RH,%)等微氣象參數。以Pn與E的比值計算瞬時水分利用效率(WUE,μmolCO2·mmol-1H2O)。
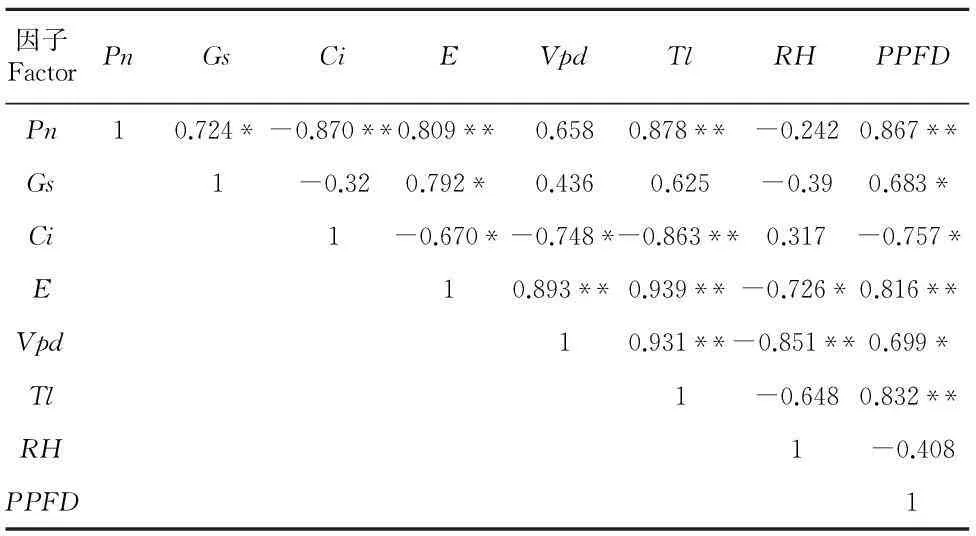
1.3.3 數據分析所測數據采用Excel計算平均值和標準誤,用軟件SPSS16.0(SPSSInc.,Chicago,IL,USA)進行統計分析, 用SigmaPlot10.0軟件作圖。采用Pearson相關分析方法對氣體交換日變化研究結果中的氣體交換參數、微氣象因子之間的相關關系進行分析。

圖版 Ⅰ 黃槿葉片解剖結構掃描電鏡圖 A. 黃槿葉片上表皮,示角質層; B. 黃槿葉片上表皮,示角質層和蠟質層; C. 黃槿葉片下表皮,示表皮毛; D. 黃槿葉片下表皮,示氣孔; E. 黃槿葉片縱切面,示海綿組織、柵欄組織、內皮層、角質層和表皮毛等。 S. 氣孔; St. 海綿組織; Pt. 柵欄組織; Cu. 角質層; Hy. 內皮層。標尺:1,3,5 =100 μm; 2,4=20 μm。Plate Ⅰ Anatomical structure of Hibiscus tiliaceus under scanning electron microscope A. Upper cuticle in the upper epidermis; B. Upper cuticle and wax layer in the upper epidermis of leaf; C. Trichomes in the lower epidermis of leaf; D. Stomata in the lower epidermis of leaf; E. Palisade tissue, upper hypodermis, spongy tissue, upper cuticle and wax layer, transverse sections of leaf. S. Stomata; St. Spongy tissue; Pt. Palisade tissue; Cu. Cuticle; Hy. Upper hypodermis. Scale bars:1,3,5 =100 μm; 2,4=20 μm.
2結果與分析
2.1 黃槿葉片的解剖結構特征
研究樣地內黃槿葉片的氣孔密度每1 mm2為(1 112.77±145.55)個,是大亞灣黃槿的三倍(繆紳裕等,2001)。葉片上表皮的外壁有一層角質層(圖版I:1,5),角質層上面覆蓋顆粒狀的蠟質(圖版I:2);上表皮下具有內皮層,內皮層細胞體積大于上表皮細胞,葉肉柵欄組織和海綿組織明顯(圖I:5);下表皮密生星狀表皮毛 (圖I:3,圖I:5)。氣孔呈橢圓形,腎形的保衛細胞周圍有一圈副衛細胞,部分氣孔被表皮毛覆蓋(圖I:4)。
2.2 黃槿葉片的光響應特征
從表1可以看出,廣東珠海淇澳島黃槿居群的光飽和點(LSP)和光補償點(LCP)略低于海南東寨港黃槿海岸種群(李妮亞等,2011),高于夏威夷瓦胡島河口黃槿種群(Santiago et al,2000);但表觀量子效率是三個種群中最高的,并且最大凈光合速率也遠大于其他兩個種群。另外,從光響應曲線來看,淇澳島黃槿LSP在800 μmol photons·m-2·s-1左右(圖1),因為從光響應曲線估算出的光飽和點比用經典方程計算出的LSP更接近真實值(陳根云等,2006;云嵐,2009),所以,淇澳島黃槿種群的光飽和點也是三個種群中最大的,這和其較高的表觀量子效率和凈光合速率是一致的。
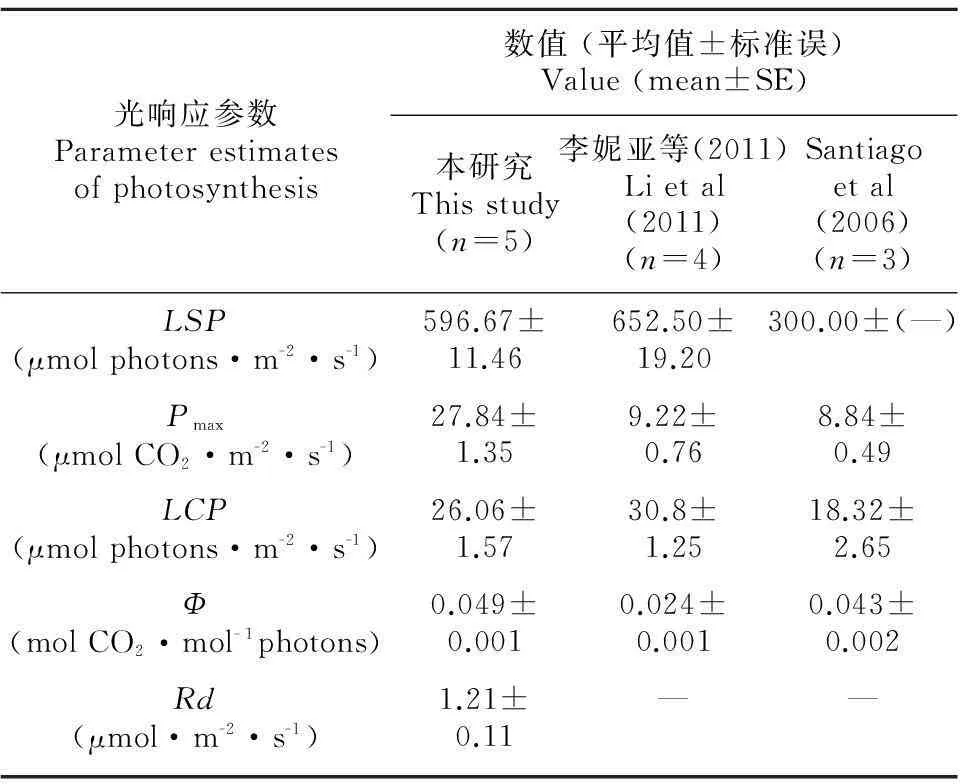
表 1 黃槿葉片的光響應曲線參數
注: LSP. 光飽和點; Pmax. 光飽和點時的凈光合速率; LCP. 光補償點; Φ. 表觀量子效率; Rd. 暗呼吸速率。
Note: LSP. Light saturation point; Pmax. Net photosynthesis at saturating irradiances; LCP. Light compensation point; Φ. Apparent quantum yield; Rd. Respiration rate.
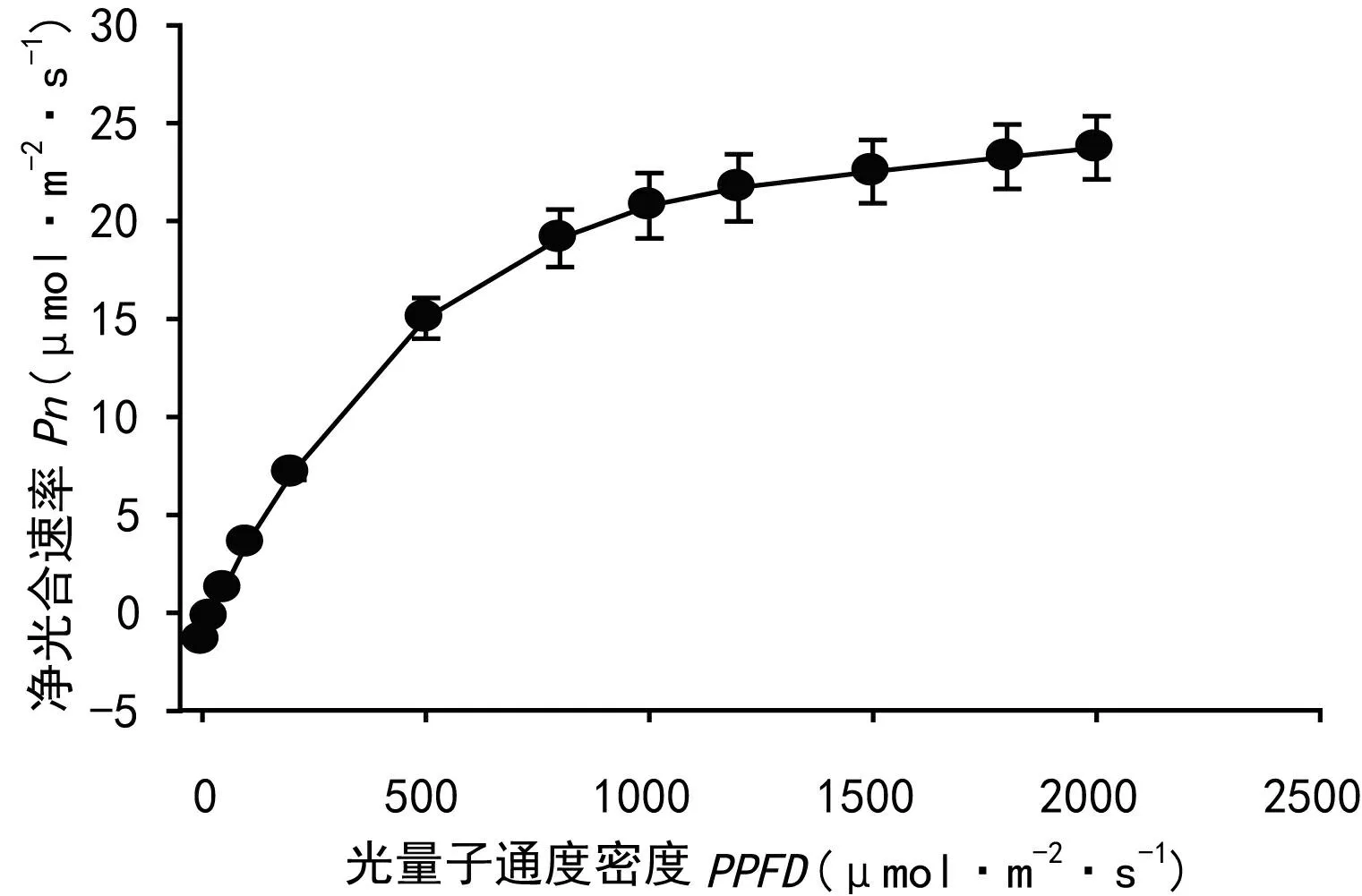
圖 1 黃槿葉片氣體交換對光強的響應曲線Fig.1 Response of net photosynthetic CO2 assimilation (Pn) to photosynthetic photon flux densities (PPFD) in the leaves of H. tiliaceus
2.3 黃槿葉片氣體交換日變化特征
2.3.1 實驗樣地小環境 從圖2可以看出,在實測的時間范圍內,PPFD的變化范圍為850~1 500 μmol photons·m-2·s-1。上午10:00 前后光強達最大值,18:00最低,但仍為850 μmol photons·m-2·s-1;氣溫(Ta)日變化范圍在35~40 ℃;空氣相對濕度(RH,% ) 從上午8:00至10:00呈線性下降趨勢,至11:00有小幅上升,其后一直到下午14:00呈線性下降,14:00以后又呈線性上升。CO2濃度(Ca)的日變化在369~381 μmol·m-2·s-1之間,基本保持相對穩定。
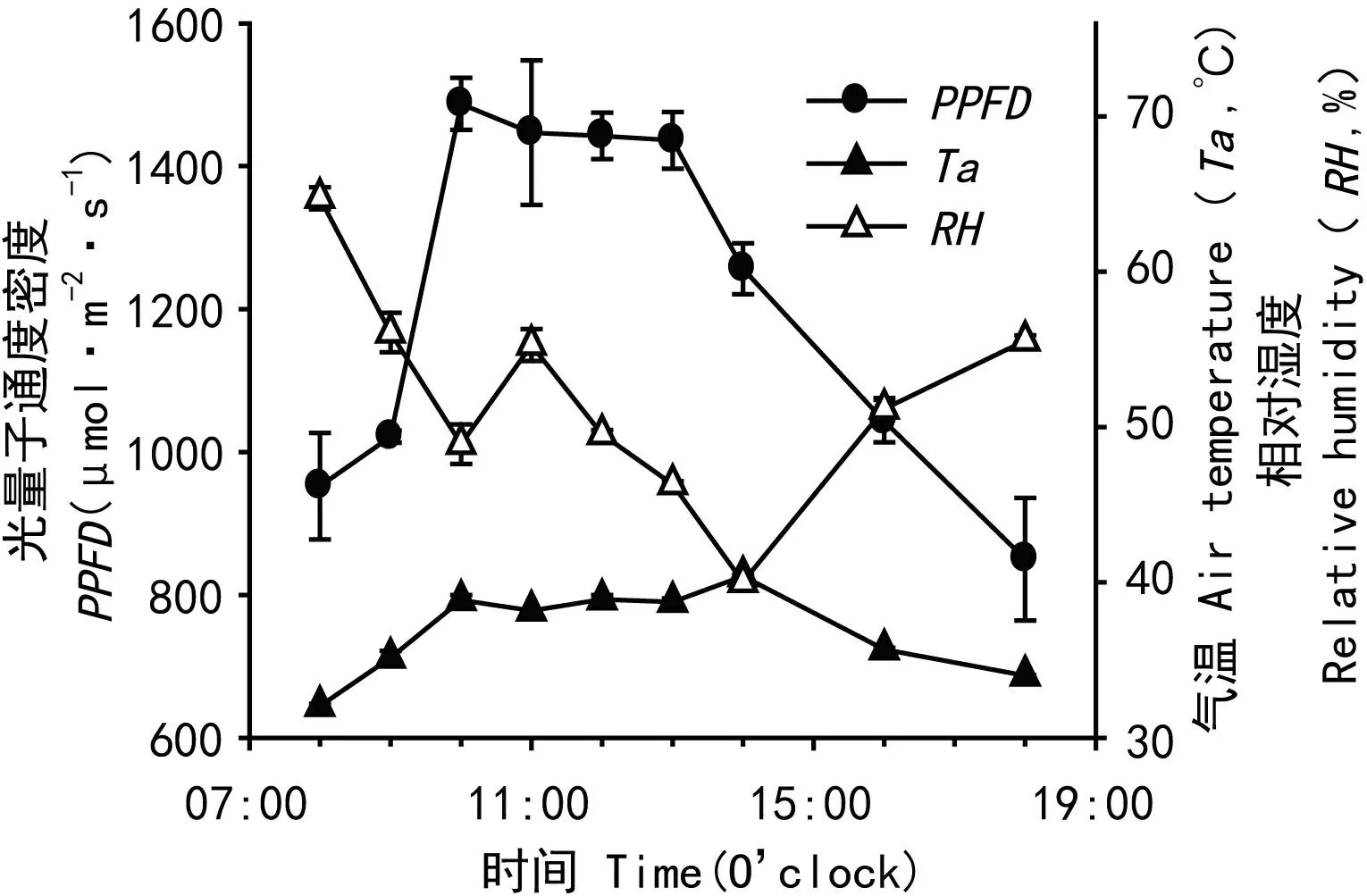
圖 2 樣地輻射強度、氣溫和相對濕度的日變化Fig.2 Diurnal changes in photosynthetic photon flux density (PPFD), air temperature (Ta) and relative humidity (RH) at the experimental site
2.3.2 氣體交換參數日變化在自然生境條件下,黃槿葉片的Pn8:00開始隨著PPFD的線性上升亦呈線性上升趨勢,至11:00達到最大值26.78 μmol CO2·m-2·s-1(圖3:A),且10:00~14:00一直保持較高的凈光合速率,未出現明顯下降。下午16:00以后Pn隨著光強下降而降低。黃槿葉片的日均凈光合速率達19.61 μmol CO2·m-2·s-1,高于東南沿海5種紅樹植物的最大凈光合速率(Pmax<15 μmol CO2·m-2·s-1)(黃敏參等,2012)和廣東省4種海岸帶迎風坡面植物的最大凈光合速率(Pmax<14 μmol CO2·m-2·s-1)(廖凌娟等,2011)。從一天中Pn的動態變化來看,黃槿的凈光合速率日變化呈單峰曲線,未出現明顯的光抑制。
黃槿葉片Gs的日變化趨勢與其Pn的日變化基本同步。11:00 凈光合速率達到最高值時(圖3:A),Gs高達0.49 mol H2O·m-2·s-1(圖3:B),此后,11:00~13:00,Gs沒有下降反而上升36%,達到一天中最大值,0.67 mol H2O·m-2·s-1。這在其他植物的光合日變化研究中比較少見,可能是黃槿中午保持較高的光合能力而未出現光抑制的原因之一。
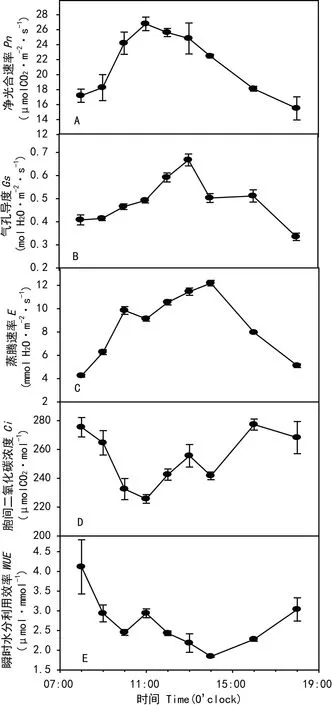
圖 3 氣體交換參數的日變化 黃槿葉片凈光合速率(A)、氣孔導度(B)、 蒸騰速率(C)、胞間二氧化碳濃度(D)、瞬時水分利用效率(E)Fig. 3 Diurnal changes of H. tiliaceus leavesNet photosynthetic rate (Pn,A), Stomata conductance (Gs, B), transpiration rate (E, C), intercellular CO2 concentration (Ci, D) and instantaneous water use efficiency (WUE, E)
圖3:C顯示,黃槿葉片E的日變化動態與Pn和Gs的相似,峰值(12.19 mmol H2O·m-2·s-1)出現在14:00,最低值出現在8:00 (4.33 mmol H2O·m-2·s-1)。但在12:00到14:00,E出現了輕微上升,這與此期間氣溫的上升(圖2)和Gs的上升有關。
黃槿葉片的Ci日變化曲線(圖3:D)與Pn相比總體呈相反趨勢。從圖3:E可以看出,除了14:00,黃槿一天的WUE有較高的水平。日均WUE為2.7 μmol·mmol-1,接近東南沿海5種紅樹植物的最大瞬時水分利用效率(2.5 表 2 黃槿的氣體交換參數及其與微氣象因子的相關關系 注: *, **分別表示在0.05和0.01概率水平上的相關性; Pn. 凈光合速率; Gs. 氣孔導度; Ci. 胞間二氧化碳濃; E. 蒸騰速率; Vpd. 葉面大氣蒸汽壓虧缺; Tl. 葉面溫度; RH. 大氣相對濕度; PPFD. 光量子通度密度。 Note:*, ** show significant correlations atP=0.05 andP=0.01; Pn. Net photosynthetic rate; Gs. Stomatal conductance; Ci. Intercellular CO2concentration; E. Transpiration rate; Vpd. Air vapor pressure deficit of leaf; Tl. Leaf temperature; RH. Relative humidity; PPFD. Photosynthetic photon flux density. 2.3.3 氣體交換參數及其與微氣象因子的關系從表2可以看出,4個氣體交換參數(Pn,Gs,Ci,E)之間除了Gs和Ci之外,均兩兩顯著相關。Pn與環境中Tl、PPFD呈顯著正相關。Gs只與環境因子中PPFD呈顯著正相關,Ci則除了RH之外與其他參數均顯著相關;E則與所有氣體交換參數及微氣象因子均顯著相關。因此,黃槿Pn的大小與葉片的生理生化因子和外界的氣象因子相關,各氣體交換參數主要受溫度和輻射強度的影響較大。 3討論與結論 在海岸帶強光高溫的鹽生環境中,多數真紅樹植物、半紅樹植物和其他海岸帶植物一般具有較低的LSP和Pmax,有些植物種類中午出現明顯的光抑制。例如,南美洲伯利茲珊瑚礁真紅樹植物的LSP為300~500 μmol·m-2·s-1(Cheeseman & lovelock,2004);東寨港幾種半紅樹植物的Pmax為6~15 μmol CO2·m-2·s-1(李妮亞等,2011);海岸帶迎風坡面幾種陽生樹種的Pmax為8~16 μmol CO2·m-2·s-1(廖凌娟等,2006)。福建泉州3種真紅樹植物出現明顯的光抑制現象(劉榮成等,2006)。本研究表明,在夏季高溫強光環境中,廣東省珠海淇澳島紅樹林自然保護區的黃槿居群具有較強的光合能力,整個白天保持較高的凈光合速率,且中午未出現明顯的光抑制。針對本研究中黃槿不同于其他研究中紅樹的光合行為,以下從其解剖結構的適應特征、氣孔導度和陽生植物特性等方面進行分析。 (1)黃槿葉片上表皮具有豐富的角質層和蠟質層,可減少水分的蒸發。葉肉組織分化為海綿組織和柵欄組織,增加貯水機能和提高生理生化反應速度。下表皮毛和真紅樹植物白骨壤(Avicenniamarina)一樣發達,不僅可以保護葉肉組織,還可以部分遮蓋分布在表皮的氣孔,從而減少水分從氣孔的蒸發,過濾部分過剩光能,進而減少高溫和強光的傷害(李元躍,2006)。本研究發現,黃槿葉片下表皮的氣孔密度每1 mm2高達1 112.77 個,而秋茄、木欖等幾種真紅樹植物的氣孔密度每1 mm2為100~300 個 (李元躍,2006);長在海岸帶坡面的紅錐等5種喬木的氣孔密度每1 mm2為100~700個 (廖凌娟等,2011);氣孔密度越大,氣孔導度越高,有利于蒸騰作用的進行,這使其能夠在強烈光照下保持植物體內水分平衡,提高水分利用效率,從而有效增加生物量(張偉偉等,2012)。 (2)Youssef (2007)針對高鹽和干旱環境下黃槿的光合作用氣體交換研究表明,氣孔在限制凈光合速率方面比生化因子起著更為關鍵的作用。本研究針對光響應曲線和光合日變化研究發現,Pn和Gs均呈顯著正相關關系(相關系數分別為r=0.978**,r=0.724*),進一步針對日變化研究中氣體交換參數、微氣象因子相關性分析發現,Pn和PPFD呈正相關可能間接與Gs和PPFD的顯著正相關有關,推測Gs是限制黃槿Pn的主要內在因素。另外,黃槿的氣孔導度不僅高于一般紅樹植物的氣孔導度,而且與具有光抑制的紅樹植物中午出現Gs的下降顯著不同的是黃槿在中午時段Gs表現出小幅升高。因此,黃槿中午時段維持較高Gs可能是光合日變化未出現明顯光抑制的原因之一。 (3)按照Larcher(1980)的觀點,光補償點低于10 μmol photons·m-2·s-1、光飽和點低于180 μmol photons·m-2·s-1的植物一般為陰生植物;而光補償點在20 μmol photons·m-2·s-1以上、光飽和點在500 μmol photons·m-2·s-1以上的植物一般為陽生植物。本研究結果表明,黃槿的光補償點和光飽和點在陽生植物范圍內,最大凈光合速率高于一般木本植物(5~10 μmol CO2·m-2·s-1),且接近于草本C3植物(20~25 μmol CO2·m-2·s-1), 這與許大全(2002)的研究結果一致。因此,總體來說黃槿為典型的陽生植物。另外,在光合日變化和光響應曲線中Pn和Ci成極顯著負相關(相關系數分別為r=-0.870**,r=-0.988**),表明擴散到葉綠體的CO2被羧化的效率較高,亦符合陽生植物特性。從氣體交換日變化動態可以看出,黃槿在白天能維持較大的Pn,且日均凈光合速率接近其最大光合能力,因此可以保持較高的光合生產能力,有利于其快速生長和種群的擴展。 參考文獻: CHEESEMAN JM,LOVELOCK CE, 2004. Photosynthetic characteristics of dwarf and fringeRhizophoramangleL. in a Belizean mangroves [J]. Plant Cell Environ, 27: 769-780.CHEN GY,YU GL,CHEN Y,et al, 2006. Exploring the observation methods of photosynthetic responses to light and carbon dioxide [J]. J Plant Physiol Mol Biol,32(6): 691-696. [陳根云,俞冠路,陳悅,等, 2006. 光合作用對光和二氧化碳響應的觀測方法探討 [J]. 植物生理與分子生物學學報,32(6): 691-696.] FENG GM, 1984. Flora of China [M]. Beijing: Science Press,49(2): 64. [馮國楣, 1984. 中國植物志 [M]. 北京: 科學出版社,49(2): 64.] HUANG MS,DU XN,LIAO MM,et al, 2012. Photosynthetic characteristics and water use strategies of coastal shelterbelt plant species in Southeast China. [J]. Chin J Ecol,31(12): 2 996-3 002. [黃敏參,杜曉娜,廖蒙蒙,等, 2012. 東南沿海潮間帶防護林主要樹種的光合特性及水分利用策略 [J]. 生態學雜志,31(12): 2 996-3 002.] LAMBERS H,CHAPIN III FS,PONS TL, 1998. Plant physiological ecology [M]. New York: Springer-Verlag:540. LARCHER W, 1980. Physiological plant ecology [M]. Beijing: Academic Press: 57. LI NY,HAN SM,LIU Q,et al, 2011. Ion accumulation, photosynthetic characteristics of mangrove associates in divergent habitats [J]. J Hainan Norm Univ: Nat Sci Ed,24(4): 429-438. [李妮亞,韓淑梅,張俊清,等, 2011. 不同生境中半紅樹植物鹽離子積累與光合特性的研究 [J]. 海南師范大學學報·自然科學版,24(4): 429-438.] LI YY, 2006. Studies on leaf anatomy of some mangrove species [D]. Xiamen: Xiamen University: 153. [李元躍, 2006. 幾種紅樹植物葉的解剖學研究 [D]. 廈門: 廈門大學: 153.] LIAO LJ,LI Q,CHEN YZ,et al, 2011. Effects of environmental disturbance on leaf morphological traits,stomata,and water use efficiency of species for afforestation on a windward slope in a coastal zone [J]. Plant Sci J, 29(5): 613-624. [廖凌娟,黎清,陳貽竹,等, 2011. 海岸帶迎風坡環境干擾對造林植物葉片解剖、氣孔和水分利用效率的影響 [J]. 植物科學學報, 29(5): 613-624.] LIU RC,WU YY,FU WG,et al, 2009. Chlorophyll fluorescence characteristics of several plant species in Quanzhou Bay estuarine wetland [J]. J Fujian Coll For,29(4): 374-379. [劉榮成,吳沿友,付為國,等, 2009. 泉州灣河口濕地幾種植物葉綠素熒光特性 [J]. 福建林學院學報,29(4): 374-379.] MIU SY,WANG HL, 2001. Stomatal characteristics and its development of leaves in mangroves and coastal plants at Daya Bay [J]. J Oceanogr Taiwan Strait,20(2): 251-258. [繆紳裕,王厚麟, 2001. 大亞灣紅樹林與海岸植物葉片氣孔特征及其發育 [J]. 臺灣海峽,20(2): 251-258.] NICHOL CJ,RASCHER U,MATSUBARA S, 2005. Assessing photosynthetic efficiency in an experimental mangrove canopy using remote sensing and chlorophyll fluorescence [J]. Trees, 20(1): 9-15. PARIDA AK,JHA B, 2010. Salt tolerance mechanisms in mangroves: a review [J]. Trees, 24(2): 199-217.ROSA RM,MELECCHI MI,DA COSTA HALMENSCHLAGER R,et al, 2006. Antioxidant and antimutagenic properties ofHibiscustiliaceusmethanolic extract [J]. J Agr Food Chem,54: 7 324-7 330. SANTIAGO LS, LAU TL, MELCHER PJ,et al, 2000. Morphological and physiological responses of three populations of HawaiianHibiseustiliaeeusto light and substrate salinity [J]. Int J Plant Sci,161: 14-20.TAKAYAMA K,KAJITA T,MURATA J,et al, 2006. Phylogeography and genetic structure ofHibiscustiliaceus-speciation of a pantropical plant with sea-drifted seeds [J]. Mol Ecol,15(10): 2 871-2 881. TAN R, 2001. SeaHibiscusmangrove and wetland wild life at Sungei Buloh Nature Park [EB/OL]. http://www. naturia. per. sg/buloh/plants/sea_hibiscus. htm. TOMLINSON PB, 1986. The botany of mangroves [M]. Cambridge: Cambridge University Press: 374-381. WALKER DA,JARVIS PG,FARQUHAR GD,et al, 1989. Automated measurement of leaf photosynthetic O2evolution as a function of photon flux density [J]. Phil Trans RSoc Lond B Biol Sci, 323: 313-326. WANG BS,LIANG SC,ZHANG WY,et al, 2003. Mangrove flora of the world [J]. Acta Bot Sin,45(6): 644-653. [王伯蓀,梁士楚,張煒銀,等, 2003. 世界植物紅樹區系 [J]. 植物學報,45(6): 644-653.] XU DQ, 2002. The Efficiency of photosynthesis [M]. Shanghai: Shanghai Science and Technology Press:46. [許大全, 2002. 光合作用效率 [M]. 上海: 上海科學技術出版社:46.] YOUSSEF T, 2007. Stomatal,biochemical and morphological factors limiting photosynthetic gas exchange in the mangrove associateHibiseustiliaeeusunder saline and arid environment [J]. Aquat Bot, 87: 292-298. YUN L, 2009. Polyploidy induction and cytological analyses of russian wild ryegrass [D]. Hohehot: Inner Mongolia Agricultural University: 67. [云嵐, 2009. 新麥草多倍體誘導及細胞學研究 [D]. 呼和浩特: 內蒙古農業大學: 67.] ZHANG WW,LIU N,WANG J,et al, 2012. Ecological and biological characteristics ofHibiscustiliaceus,a mangrove associate in China [J]. Guihaia, 32(2): 198-202. [張偉偉,劉楠,王俊,等, 2012. 半紅樹植物黃槿的生態生物學特性研究 [J]. 廣西植物, 32(2): 198-202.] 中圖分類號:Q945.79 文獻標識碼:A 文章編號:1000-3142(2016)04-0397-08 作者簡介:蔡水花(1987-),女,四川簡陽人,碩士,主要從事植物生理生態學研究,(E-mail)celery-99@163.com。*通訊作者: 沈浩,博士,副研究員,主要從事保護生態學和植物生理生態學研究,(E-mail)shenhao@scbg.ac.cn。 基金項目:國家科技支撐計劃項目(2012BAC07B04)[Supported by National Key Technology R & D Porgram of China(2012BAC07B04)]。 *收稿日期:2014-10-28修回日期: 2014-12-10 DOI:10.11931/guihaia.gxzw201403011 蔡水花,李婷,周光霞,等. 半紅樹植物黃槿的氣體交換特性 [J]. 廣西植物, 2016, 36(4):397-404 CAI SH,LI T,ZHOU GX,et al. Gas exchange characteristics in the mangrove associateHibiscustiliaceus[J]. Guihaia, 2016, 36(4):397-404