葉面鎂營養促進妃子笑荔枝果皮著色的生理成因
2016-05-30 15:30:31周開兵周曉超蘇陽高丹
熱帶作物學報 2016年9期
周開兵 周曉超 蘇陽 高丹
摘 要 以妃子笑荔枝(Litchi chinensis Sonn. cv. Feizixiao)成年樹為試材,探討葉面 Mg 營養調節妃子笑荔枝果皮著色的效果及其初步原因。以葉面噴Mg為處理,以噴清水為對照(CK),分別在坐果后1 d(2013 年5月5 日)和8 d進行葉面噴肥處理,觀測果皮著色、內源激素含量和相關關鍵酶活性的動態變化。在坐果后15 d,Mg處理的果皮h值最低和果肉可溶性糖含量最高,克服了果皮“滯綠”現象。在果實生長發育過程中,Mg處理與CK的果皮生長素(IAA)和乙烯(Eth)含量、葉綠素單加氧酶(PaO)和葉綠素酶(Chlase)活性在同期間均無顯著差異。在坐果后11 d后,Mg處理在果皮赤霉素(GA3)和脫落酸(ABA)含量及類黃酮糖基轉移酶(UFGT)活性上均顯著高于同期CK,并且Mg處理使果皮 ABA/GA3(含量比)高于CK。Mg處理在坐果后15 d和18 d使其果皮花色素苷含量顯著高于CK,而使其葉綠素含量與CK無顯著差異。可見,Mg處理克服果皮“滯綠”現象歸功于果皮花色素苷含量的升高,其可能通過提高ABA/GA3值,進而刺激UFGT活性升高,從而促進果皮花色素苷合成。
關鍵詞 妃子笑荔枝;Mg;果皮著色;內源激素;酶
中圖分類號 S667.1 文獻標識碼 A
荔枝(Litchi Chinesis Sonn.)在中國主要分布于廣東、廣西、福建、海南和臺灣等省區,是華南區栽培面積最大的果樹[1],研究其果實品質發育問題對促進荔枝產業和華南區農村經濟發展有重要的意義。荔枝果皮著色轉紅和果肉風味變佳均是果實成熟的重要標志,也分別是最重要的外觀和內在品質因素。因此,商品價值高的荔枝果實應該是荔枝果皮轉紅和果肉風味變佳同步發育的,其中果肉風味品質變佳表現為果肉含糖量升高和總酸含量下降,糖酸比達到最高時果肉即已經成熟[2]。大多數荔枝品種果皮著色和果肉風味變佳同步發育,然而少數品種如妃子笑荔枝果肉含糖量最高時果皮未完全轉紅(即“滯綠”現象),果面全紅時的果肉含糖量發生下降(即“退糖”現象)[3],存在果皮著色轉紅和果肉風味變佳發育不同步的現象,在果肉風味最佳時采收則出現所謂“果皮著色不良”的問題。妃子笑荔枝的果皮“滯綠”問題和果肉“退糖”問題均使生產者蒙受經濟損失。因此,有必要開展妃子笑荔枝果實內在和外觀品質發育不同步問題的理論成因和解決辦法等研究。
由于荔枝果皮著色表現為果皮花色素苷含量升高,同時果皮葉綠素含量下降,而與果皮類胡蘿卜素含量變化關系不明顯[4-8]。妃子笑荔枝果皮花色素苷合成與類黃酮基轉移酶(UFGT)活性變化有關,隨著果皮中UFGT活性的增加,花青苷含量上升;套袋處理抑制UFGT活性的同時也抑制花青苷的合成,除袋后UFGT活性和花青苷含量都迅速增加;6-BA處理抑制UFGT酶活性的同時也抑制花青苷合成,ABA和茉莉酸處理提高了UFGT酶活性的同時也促進了花青苷的合成[9-10]。因此,要解決妃子笑荔枝果皮轉紅和果肉風味變佳發育不同步問題,可以通過調節妃子笑荔枝果皮色素代謝來使二者發育實現同步。由前人報道可見,對妃子笑荔枝果實套袋能解決其果皮“滯綠”問題,并且主要是套袋通過調節果皮UFGT活性而調節花色素苷合成和著色
筆者所在課題組最近已報道,對妃子笑荔枝在坐果后進行葉面噴施鎂肥處理,可成功地克服了其果皮“滯綠”問題[11],然而其生理學機制又是怎樣的?筆者從果皮激素含量和果皮色素代謝相關關鍵酶活性變化上對此問題展開初步探討。
1 材料與方法
1.1 材料
試驗地點為海南省農業科學院熱帶果樹研究所永發科研示范基地,該基地位于澄邁縣北部平原臺地,屬于熱帶季風氣候區,年平均氣溫23.8 ℃,年平均日照時數2 059 h,年均降雨量1 786.1 mm,且熱雨同季,終年無霜;土壤為肥沃的磚紅壤。在該基地荔枝園選16年生、生長勢一致和健壯的妃子笑荔枝/大丁香荔枝砧嫁接樹10株作為試驗材料。
1.2 方法
1.2.1 試驗設計 以葉面噴清水為對照(CK),依據MgCl2在果樹上用于葉面噴肥時的常用濃度,設置1.5%MgCl2溶液為處理(Mg),進行葉面噴肥處理時,噴施至葉面和葉背滴水。單株小區,5次重復。坐果后,在坐果后1 d(2013年5月5日)和坐果后5 d的上午9~10時進行葉面噴肥處理。
1.2.2 取樣及樣品處理 坐果后,果實生長發育即進入快速膨大期,此時假種皮剛剛包滿種子,即2013年5月5日,視作坐果后1 d,在每株樹樹冠外圍中部選5個大小基本一致的果實作為每次取樣的標準果,并掛牌標記,此后在不同時期對各樹動態采果樣時均參照這5個標準果平均大小,采集與其平均果徑一致的樣果。從果實開始膨大時開始到果皮全紅時為止,共取樣5次,即在坐果后1、8、11、15、18 d各取樣1次,其中坐果后1、8 d取樣均在葉面噴肥處理之前完成。果樣取自樹冠中部外圍,每次取樣采果30個/株。在田間先測定果實著色指標值,再將樣果剝皮,并將果皮和果肉分別及時用液氮速凍,帶回實驗室儲存于-80 ℃超低溫冰箱中備用。
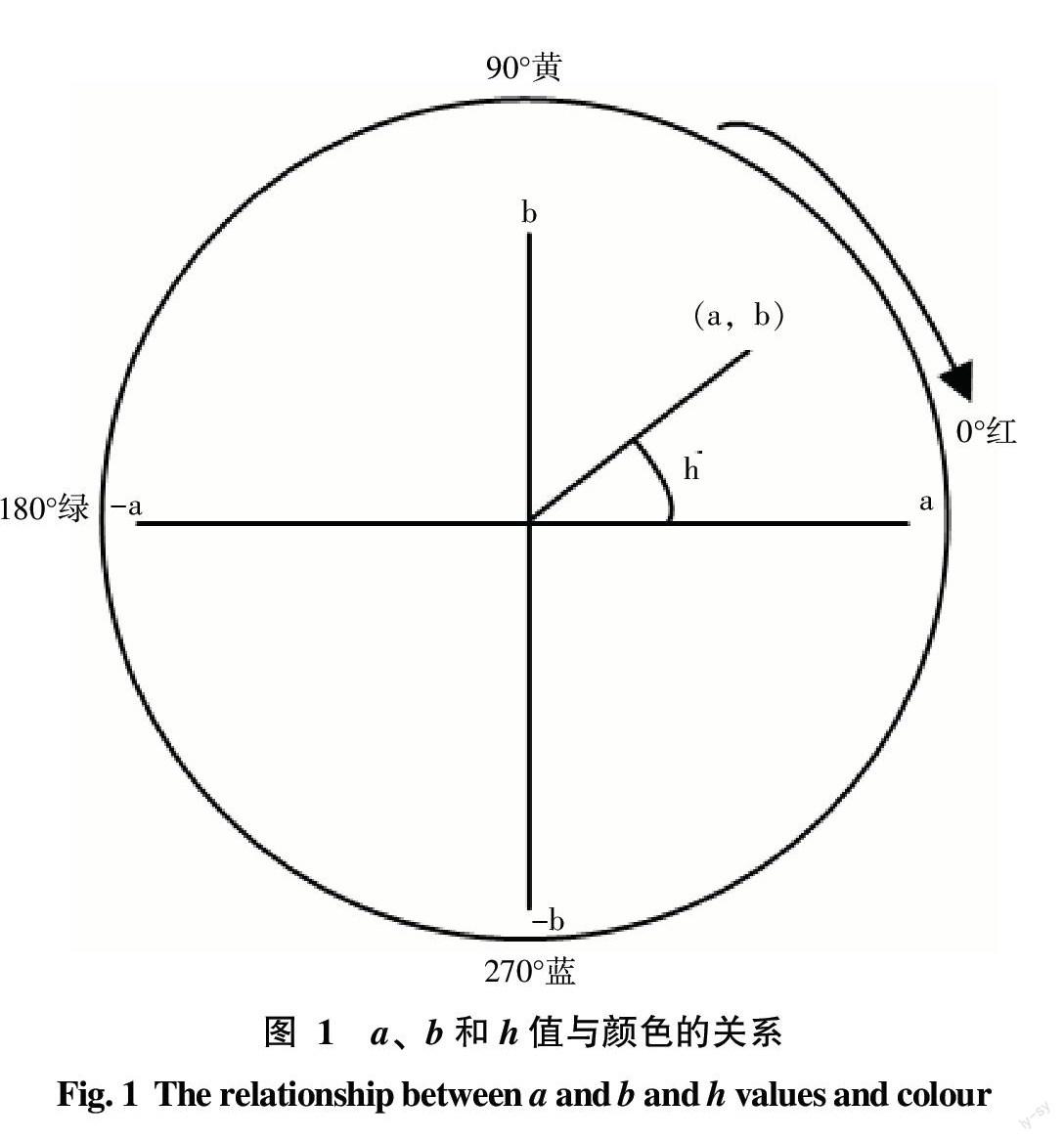
1.2.3 試驗指標觀測方法 采用日本產Minolta CR2300型全自動測色色差計測定荔枝果皮a、b值,換算出色度角(h),公式為h=tan-1(b/a),a、b和h值與顏色的關系如圖1所示,其中h值越小則果皮綜合色澤越紅,各指標以每株樣樹上30個樣果測定值的平均值作為該單株(重復)的觀測值。以下各生理生化指標測定時,每株樣樹(重復)的果皮和果肉樣品分別制備3個測樣,取其平均值作為該株樹(重復)的觀測值。采用改良Arnon法測定荔枝果皮葉綠素含量[12];采用Pirie等[13]和張昭其等[14]提出的方法測定荔枝果皮花色素苷含量;采用蒽酮比色法測定荔枝果肉可溶性糖含量[15];果皮生長素(IAA)、赤霉素(GA3)、細胞分裂素(iPAs)和脫落酸(ABA)含量測定采用酶聯免疫法(ELISA)法[16],結合南京農業大學提供試劑盒說明書測定;果皮乙烯(Eth)釋放量測定采用龐學群等優化的GC法[17]。
脫鎂葉綠酸單加氧酶(PaO)提取與活性測定參考Hortensteiner 等[18]的方法,加以改進。20 g黑暗處理5 d的荔枝果皮,放置在研缽中,加入液氮,研磨成粉,按3 mL/g FW加入提取液(0.4 mol/L蔗糖、25 mmol/L Tris-HCl 緩沖液pH8、10 mmol/L NaCl、2 mmol/L EDTA、1 mmol/L MgC12、3 mmol/L PEG600O),25 μm尼龍網過濾,濾液移進冰浴的80 mL 離心管,10 000 ×g、4 ℃ 離心5 min,棄上清液,沉淀1 mL/g FW 標準溶于上述溶液,10 000×g、4 ℃ 離心5 min,沉淀按0.5 mL/g FW標準溶解于25 mmol/L Tris-HCl pH 8緩沖液(內含1% Tritonx-100),暗中4 ℃振蕩30 min,150 000×g、4 ℃離心60 min,上清液備用。加入25 μL酶液與下列溶液(20 mmol/L Pheide a、60 mmol/L NADPH、100 mmol/L Glc6P、10 mU/μL Glc6P-DH、10 μg/μL Fd)各1 μL混勻,暗中室溫反應30 min,加入冷甲醇至終濃度70%終止反應,11 000×g離心2 min,取10 μL上清液進行HPLC分析。以HPLC色譜峰面積/min·g FW表示酶活力(或降解速率)。色譜條件:柱壓2 MPa;pH范圍:1;狹縫:10 nm;FD 激發波長320 nm,發射波長450 nm;流速0.8 mL/min;洗脫液為[67.5% 無水甲醇+32.5% PBS (pH7.0)]。
采用甘志軍等[19]的方法測定荔枝果皮葉綠素酶(Chlase)活性。將荔枝果皮在冰浴中加入液氮碾磨成粉末,加入一定量的PVP,按照0.167mL/mg標準加入緩沖溶液(0.05 mol/L pH7.5磷酸鹽緩沖液,含50 mmol/L KCl、0.24% TritonX-100),4 ℃ 抽提4 h,在4 ℃條件下10 000 r/min 離心15 min,取上清液,12 000 r/min 離心20 min,取上清液定容至一定體積,得到葉綠素酶粗酶液。用 80%丙酮從鮮綠菠菜葉片中提取色素,然后加入一定體積的石油醚萃取,葉綠體色素(主要是Chl),在醚相濃縮,醚相用冷風吹干后得到的Chl溶解于確定體積的丙酮中備用,取1 mL 色素提取液加入24 mL 丙酮,測定葉綠素OD值,然后按以下公式計算葉綠素含量:Chl含量(μmol/L)=805×OD652,Chl的分子量按900計。反應混合物3.5 mL,其中包括0.5 mL 50 mmol/L pH 7.5 PBS(內含 0.24% Triton X-100)、l mL Chl 液和2ml Chl 丙酮溶液,混合物放置40 ℃水浴中,反應30 min后,取0.5 mL混合物與4.5 mL提取液(丙酮 ∶ 石油醚=1 ∶ 2,V ∶ V)混合并充分振蕩,然后8 000 g、4 ℃、離心l0 min(若不分層,可以加入適量NaCl),取下相測定665、667、 651 nm的A值,消光系數分別按54.1、74.9、47.0 mmol-1·cm-1計算 Chl、Chla、Chlb 的反應量,酶活力(或酶促反應速度)以μmol/(min·g FW)表示。以沸水浴加熱失活的酶液同樣處理為空白對照。
UFGT 酶液的提取采用Murray[20]的方法,其活性測定采用Lister等[21]的方法。取樣品1 g,加液氮研磨后加入5 mL、20 ℃的丙酮混勻離心,棄去上清液,用4 mL -20 ℃丙酮再提取1次,沉淀用4 mL 溶液[0.1 mol/L 硼酸緩沖液(pH8.8)的5 mmol/L抗壞血酸]提取,上清液為UFGT酶的粗提液。取酶液0.5 mL加入0.5 mL的反應液[0.05 mol/L 二甘氨酸緩沖液(pH8.0)、1 mmol/L的櫟精和2.5 mmol/L的UDP-葡萄糖],反應液在34 ℃水浴30 min后,用0.75 mL 20%三氯乙酸的甲醇溶液終止反應,于5 000 r/min離心5 min,上清液貯藏于-20 ℃待測。檢測櫟精的減少量并用其表示酶活性。360 nm處的紫外分光光度法測定櫟精的含量,計算減少量,并用其表示酶活性。
1.3 數據統計分析
采用SAS軟件ANOVA過程作方差分析;采用DUNCAN法作多重比較分析;采用 TTEST過程作處理與CK差異顯著性分析;采用CORR過程作一元線性相關性分析。
2 結果與分析
2.1 Mg處理對果皮著色與果肉風味變佳發育同步性的影響
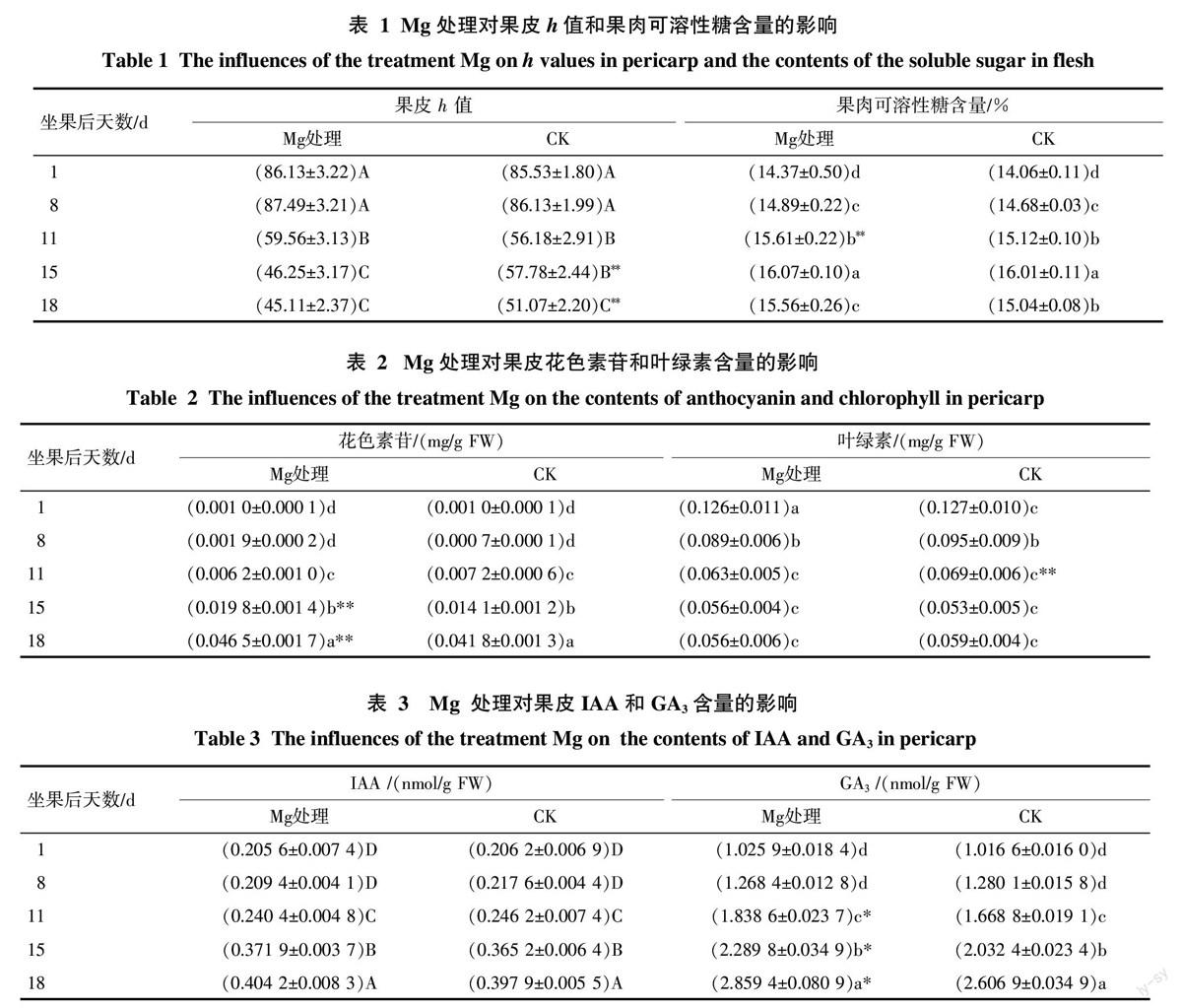
Mg處理對果皮著色和果肉可溶性糖含量的影響見表1。Mg處理與CK的果皮h值動態變化趨勢相似。二者差異出現在坐果后11 d以后,即坐果后11 d到15 d,Mg處理極顯著下降,CK則無顯著變化;坐果后15 d到18 d,Mg 處理無顯著變化,CK顯著下降。Mg處理因在坐果后15 d時h值已經降至最低且此后無顯著變化,即其已提前完全著色;CK在坐果后18 d完全著色。在坐果后15 d和18 d,Mg處理果皮h值極顯著低于CK,說明在果實成熟期Mg處理可促進果皮提早著色和著色更加紅艷。
Mg處理與CK的果肉可溶性糖含量總體變化趨勢相似;Mg處理果肉可溶性糖含量一直與CK 均差異不顯著。說明Mg處理不改變果肉可溶性糖的積累變化過程,基本上不影響在果皮全紅時的果肉可溶性糖含量。
綜上所述,表明Mg處理克服了果皮“滯綠”問題,且Mg處理果皮著色轉紅更佳,實現果皮著色轉紅和果肉風味變佳同步發育。
2.2 Mg處理對果皮色素含量的影響
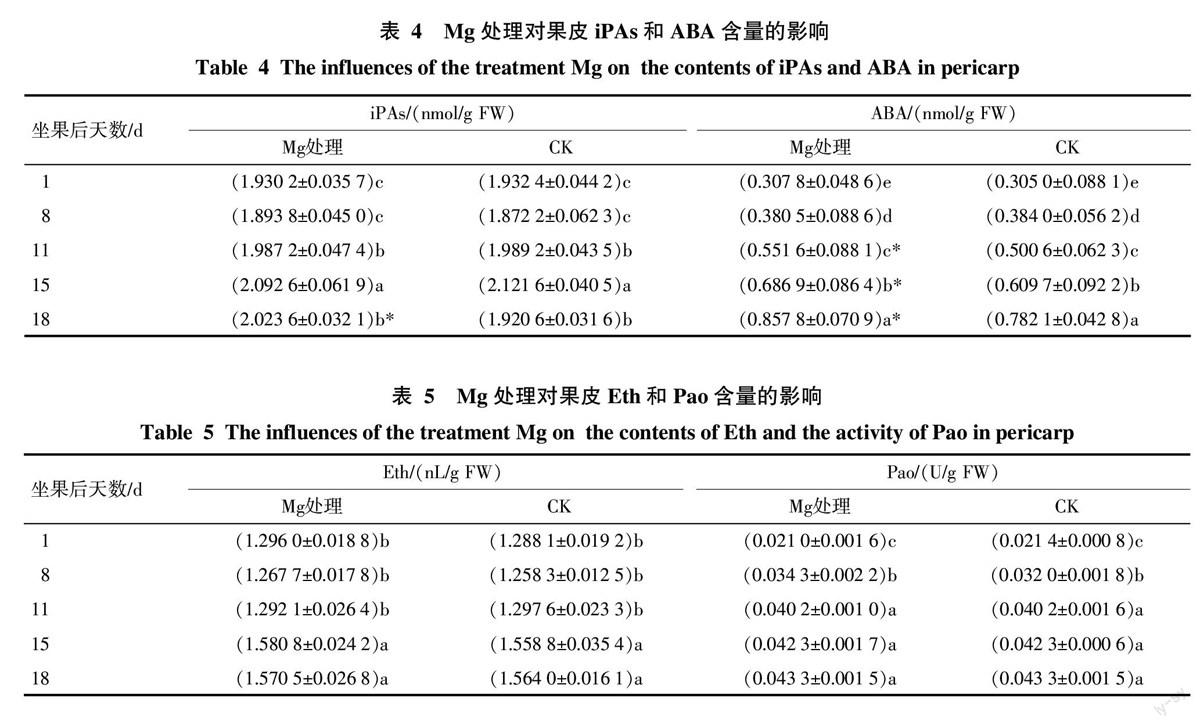
Mg處理對果皮果皮色素含量的影響見表2。Mg處理與CK的果皮花色素苷含量均呈上升趨勢,且都在坐果后18 d達到最高;Mg處理與CK的果皮葉綠素含量都呈下降趨勢。說明Mg處理不改變果皮著色的基本規律。
Mg處理與CK果皮花色素苷含量在同期間比較表明,在坐果后15 d和18 d,Mg處理極顯著高于CK。可見,Mg處理在坐果后15 d以后促進果皮花色素苷積累,即其在果實成熟期促進果皮花色素苷積累,與上述Mg處理對果皮紅色著色的影響結果一致。Mg處理果皮葉綠素含量除在坐果后11 d顯著低于CK外,在其它日期均與CK差異不顯著。說明Mg處理基本不影響果皮葉綠素代謝。總之,Mg處理調節果皮著色主要是通過調節果實成熟期時花色素苷的積累而克服“滯綠”問題。
2.3 Mg處理對果皮激素含量的影響
2.3.1 Mg處理對果皮IAA含量的影響 由表3 可見,Mg處理與CK的果皮的IAA含量都呈相似的上升趨勢,Mg處理與CK在同期果皮IAA含量間比較均無顯著差異。說明Mg處理不影響果皮IAA的代謝,即果皮IAA可能與Mg處理克服果皮“滯綠”現象無關。
2.3.2 Mg處理對果皮GA3含量的影響 由表3可見,Mg處理與CK的果皮GA3含量都呈相似的上升趨勢;除在坐果后1 d 和8 d,Mg處理與CK差異不顯著;在其余日期,Mg處理均顯著高于CK。說明Mg處理不影響 GA3含量變化趨勢,但其在果實成熟期時促進果皮GA3的積累,而此期是果皮快速轉色的關鍵時期,即果皮GA3含量的提高與Mg處理克服果皮“滯綠”現象有關。
2.3.3 Mg處理對果皮iPAs含量的影響 由表4 可見,Mg處理與CK的果皮iPAs含量呈相似的先升后降趨勢;Mg處理除在坐果后18 d顯著高于CK外,在其余時間均與CK無顯著差異。說明Mg處理基本上不改變果皮iPAs的代謝,即在本試驗中果皮iPAs可能與Mg處理克服果皮“滯綠”現象無關。
2.3.4 Mg處理對果皮ABA含量的影響 由表4 可見,Mg處理與CK的果皮ABA含量呈相似的持續顯著上升趨勢;在坐果后1 d和8 d,Mg 處理與CK差異不顯著,在其余日期,Mg處理顯著高于 CK。說明Mg處理不影響ABA含量變化趨勢,但其在果實成熟期時促進果皮ABA的積累,而此期是果皮快速轉色的關鍵時期,即果皮ABA含量的提高與Mg處理克服果皮“滯綠”現象有關。
2.3.5 Mg處理對果皮Eth含量的影響 由表5可見,在果實生長發育過程中,Mg處理與CK的果皮Eth含量呈相似的上升趨勢,Mg處理與CK在同期間比較均無顯著差異。說明Mg處理不影響果皮Eth的代謝,即在本試驗中果皮Eth可能與Mg 處理克服果皮“滯綠”現象無關。
2.4 Mg處理對與果皮著色相關的關鍵酶活性的影響
2.4.1 Mg處理對PaO活性的影響 由表5可見,Mg處理與CK的果皮的PaO活性呈相似的上升趨勢,Mg處理與CK果皮PaO活性在同期間無顯著差異。說明Mg處理不影響果皮PaO的活性變化,即在本試驗中果皮PaO可能與Mg處理克服果皮“滯綠”現象無關。
2.4.2 Mg處理對Chlase活性的影響 由表6可見,Mg處理與CK的果皮的Chlase活性都呈相似的上升趨勢,Mg處理與CK果皮Chlase活性在同期間比較均無顯著差異。說明Mg處理不影響果皮Chlase的活性變化,即在本試驗中果皮Chlase可能與Mg處理克服果皮“滯綠”現象無關。
2.4.3 Mg處理對UFGT活性的影響 由表6可見,Mg處理與CK的果皮的UFGT活性都呈上升趨勢;在坐果后1 d 和8 d,Mg處理與CK差異不顯著,在其余時間,Mg處理均顯著高于CK。說明Mg處理不影響UFGT活性的變化趨勢,但其在果實成熟期時促進果皮UFGT活性增強,而此期是果皮快速轉色的關鍵時期,與上述果皮花色素苷含量在此期快速升高一致,即果皮UFGT活性的增強與Mg處理克服果皮“滯綠”現象有關。
3 討論與結論
3.1 葉面Mg營養促進果皮花色素苷合成
對本試驗結果作線性相關性分析表明,Mg處理和CK的果皮UFGT活性與其GA3含量和ABA含量分別呈顯著正相關(GA3:Mg處理r=0.971 1*,CK r=0.995 2*;ABA:Mg處理r=0.966 1*,CK r=0.964 7*),同時其果皮花色素苷含量與果皮UFGT活性和果皮ABA含量也分別呈顯著正相關(UFGT:Mg處理r=0.910 8*,CK r=0.948 3*;ABA:Mg處理r=0.849 5*,CK r=0.843 7*)。其中,果皮花色素苷含量與果皮UFGT活性和果皮ABA含量正相關,與前人在荔枝和蘋果中的研究結果相同[9,22-,23];果皮UFGT活性與果皮GA3含量呈正相關,與蘋果中的ABA含量較高抑制GA3含量上升的結果不一致[23],但在果實成熟期,Mg處理使其果皮 ABA/GA3比值大于CK(坐果后15 d,Mg處理的0.465,CK的0.436;坐果后18 d,Mg處理的0.400,CK的0.383)。因此,從激素代謝平衡上分析表明,果皮ABA含量升高促進其UFGT活性和花色素苷含量升高。總之,果皮可能通過提高其ABA/GA3比值而提高其UFGT活性,進而促進其花色素苷合成而促進著色轉紅。在坐果后15 d 以后,Mg處理使其果皮ABA/GA3 比值在同期高于CK,從而使其果皮UFGT活性顯著高于CK,進而導致其果皮花色素苷含量顯著高于CK,最終其著色好于CK,這可能是Mg處理促進果皮著色的生理成因,同時也對妃子笑荔枝施肥技術的創新研究有一定指導意義。至于Mg如何促進果皮ABA積累,目前關于Mg營養與ABA代謝的關系未見報道,還有待于作進一步的研究。
3.2 葉面Mg營養不影響葉綠素降解
對本試驗結果作線性相關性分析還表明,Mg 處理和CK的果皮葉綠素含量與其PaO活性和Eth 含量分別均呈顯著負相關(PaO:Mg處理r=-0.995 2*,CK r=-0.993 2*;Eth:Mg處理r=-0.821 6*,CK r=-0.819 1*),與其Chlase活性變化趨勢相反。這說明果皮PaO可能也是果皮葉綠素降解的關鍵酶,與植物葉片葉綠素降解相似[24]。同時,果皮PaO和Chlase活性分別均與其Eth含量呈顯著正相關(PaO:Mg處理r=0.8130*,CK r=0.8263*;Chlase:Mg處理r=0.9057*,CK r=0.9311*);這又與前人關于乙烯促進葉綠素降解的研究結果相同[25]。總之,果皮可能是通過提高其Eth含量而提高其 PaO和Chlase活性,進而促進其葉綠素的降解而導致其葉綠素含量下降。與CK相比,Mg處理未改變果皮Eth含量的動態變化,對果皮PaO、Chlase活性和果皮葉綠素含量的動態變化也無顯著影響,這可能是果皮Eth、PaO和Chlase等與Mg處理克服果皮“滯綠”問題無關的原因。
參考文獻
[1] 倪耀媛, 吳素芬. 荔枝栽培[M]. 北京: 中國農業出版社, 1990.
[2] 張 銳, 韓麗婷, 王 娟,等. 兩個荔枝品種果皮著色和果肉糖酸積累特點的比較[J]. 福建農林大學學報, 2014, 43(4):374-378
[3] 王惠聰, 黃旭明,黃輝白. 妃子笑荔枝果實著色不良原因的研究[J]. 園藝學報, 2002, 29(5): 408-412.
[4] 張 銳, 楊 迎, 文亞迪,等. 三月紅和白糖罌荔枝果皮著色動態差異的比較[J]. 福建農業學報, 2013, 28(9): 892-896.
[5] 周開兵, 蘇 舉, 徐遠峰. 荔枝果皮著色與色素含量的關系[J]. 山地農業生物學報, 2007, 2(1): 30-33.
[6] 李 平, 陳大成, 胡桂兵,等. 荔枝果實發育過程中果皮色素的變化[J]. 熱帶亞熱帶植物學報, 1999, 71(1): 53-58.
[7] 高飛飛, 尹金華, 陳大成, 等. 荔枝果皮葉綠素、 類胡蘿卜素、 花色苷的形成規律及對果色的影響[J]. 華南農業大學學報, 2000, 21(1): 16-18.
[8] 王家保, 劉志媛, 杜中軍,等. 荔枝果實發育過程中果皮顏色形成的相關分析[J]. 熱帶作物學報, 2006, 27(2): 11-17.
[9] 王惠聰, 黃旭明, 胡桂兵,等. 荔枝果皮花青苷合成與相關酶的關系研究[J]. 中國農業科學, 2004, 37(12): 2 028 -2 032.
[10] 陳大成, 李 平, 胡桂兵,等. 套袋對妃子笑荔枝果實著色的影響[J]. 華南農業大學學報, 1999, 20(4): 65-69.
[11] 周曉超, 蘇 陽, 張 銳, 等. 葉面噴布鉀、鈣和鎂肥對妃子笑荔枝果皮著色的調節效果[J]. 西南農業學報, 2015, 28(4):1 713-1 718.
[12] 齊秀娟, 徐善坤, 李作軒,等. 山楂果實發育期礦質元素和色素含量的動態變化及相關關系[J]. 果樹學報, 2006, 32(2):214-217.
[13] Pirie A, Mullins M G. Changes in anthocaynin and phenolic content of grapevine leaf and abscisic acid[J]. Plant Phys,1976, 58: 468-472.
[14] 張昭其, 龐學群, 段學武,等. 荔枝采后果皮花色素苷的降解與花色素苷酶活性變化[J]. 中國農業科學, 2003, 36(8): 945-949.
[15] 翁雪成, 袁 紅. 蒽酮-硫酸法測定荔枝核中可溶性多糖含量[J]. 杭州師范學院學報, 2007(27): 1 007-1 008.
[16] 吳頌如, 陳婉芬, 周 燮. 酶聯免疫法(ELISA)測定內源植物激素[J]. 植物生理學通訊, 1988(5): 53-57.
[17] 龐學群, 張昭其, 段學武, 等. 乙烯與1-甲基環丙烯對荔枝采后果皮褐變的影響[J]. 華南農業大學學報, 2001, 22(4): 11-14.
[18] Hortensteiner S, Vicentini F, Matile P. Chlorophyll breakdown in senescent cotyledons of rape, Brassica napus L.: enzymatic cleavage of phaeophorbide a in vitro[J]. New Phytol, 129: 237-246.
[19] 甘志軍, 王曉云, 鄒 琦. 小麥葉綠素酶生化動力學特性研究[J]. 西北植物學報, 2003, 23(5): 750-754.
[20] Murray J R, Hackett W P. Dihydroflavonol reductase activity in relation to differential anthocyanin accumulation in juvenile and mature phase Hedera helix L. [J]. Plant Physiology,1991, 97: 343-351.
[21] Lister C E, Lancaster J E. Developmental changes in enzymes of flavonoid biosynthesis in the skins of red and green apple cultivars[J]. Journal Science of Food Agriculture, 1996, 71:313-320.
[22] Ju Z G, Liu C L, Yuan Y B, et al. Coloration potential, anthocyanin accumulation, and enzyme activity in fruit of commercial apple cultivars and their F1 progeny[J]. Scientia Horticulturae, 1999, 79: 39-50.
[23] 李 明, 郝建軍, 于 洋, 等. 脫落酸(ABA)對蘋果果實著色相關物質變化的影響[J]. 沈陽農業大學學報, 2005, 36(2):189-193.
[24] Tanaka A, Tanaka R. Chlorophyll metabolism[J]. Curr Opin Plant Biol., 2006, 9(3): 248-255.
[25] Huicong Wang, Huibai Huang, Xuming Huang. Differential effects of abscisic acid and ethylene on the fruit maturation of Litchi chinensis Sonn[J]. Plant Growth Regul, 2007, 52: 189-198.
