甘蔗與斑茅割手密復合體雜交后代的染色體遺傳分析
2016-05-30 10:48:04黃玉新羅霆劉昔輝周珊楊翠芳高軼靜楊翠鳳段維興周會張革民李楊瑞
熱帶作物學報 2016年2期
黃玉新 羅霆 劉昔輝 周珊 楊翠芳 高軼靜 楊翠鳳 段維興 周會 張革民 李楊瑞

摘 要 本研究的目的是明確甘蔗與斑割(斑茅割手密)復合體雜交后代F1材料的染色體核型構成和遺傳行為,為有效利用甘蔗野生種質提供細胞遺傳學的理論依據。采用甘蔗根尖細胞酶解去壁低滲法分別對3個不同甘蔗品種(系)與斑割復合體GXAS07-6-1的雜交后代進行染色體數目鑒定和核型分析。結果表明:各親本和后代材料染色體數目在不同細胞中不恒定,染色體變幅為6~11條。粵糖93-159與GXAS07-6-1的F1材料GXASF108-1-1染色體核型公式為 2n=94=91 m+3 sm;桂糖01-53與GXAS07-6-1的F1材料GXASF108-2-28 染色體核型公式為2n=98=88 m+10 sm(1SAT);桂糖02-761與GXAS07-6-1的F1材料GXASF108-3-7 染色體核型公式為2n=92=87 m+5 sm;3個F1材料的核型均為2B型。推斷甘蔗與斑割復合體雜交親子間的染色體基本按 “n+n”方式傳遞,同時可能存在部分染色體加倍現象,它們的雜種F1核型均為較原始的染色體2B類型。
關鍵詞 甘蔗;斑茅割手密復合體;核型分析;染色體傳遞
中圖分類號 S566.1 文獻標識碼 A
Genetic Analysis of Chromosomes for the Progeny Between
Sugarcane (Saccharum spp.)and Intergeneric Hybrid
Complex(Erianthus arundinaceus ×Saccharum spontaneum)
HUANG Yuxin1,2, LUO Ting2,3, LIU Xihui2,3, ZHOU Shan2,3, YANG Cuifang2,3,
GAO Yijing2,3, YANG Cuifeng1,2, DUAN Weixing2,3,
ZHOU Hui2,3, ZHANG Gemin2,3*, LI Yangrui2,3*
1 Agricultural College,Guangxi University,Nanning,Guangxi 530005,China
2 Guangxi Key Laboratory of Sugarcane Genetic Improvement,Nanning,Guangxi 530007,China
3 Sugarcane Research Institute,Guangxi Academy of Agricultural Sciences/Key
Laboratory of Sugarcane Biotechnology and Genetic Improvement(Guangxi),
Ministry of Agriculture,Nanning ,Guangxi 530007,China
Abstract To incorporate genes from wild germplasm for laying a good cytogenetical foundation of sugarcane, karyotype composition and genetic behavior of chromosome were explored for the F1 progeny between sugarcane (Saccharum spp.)and intergeneric hybrid(Erianthus arundinaceus×Saccharum spontaneum). Chromosome number calculation and karyotype analysis were carried out for the progeny between 3 clones(Saccharum spp.)and GXAS07-6-1(Erianthus arundinaceus×Saccharum spontaneum)by the means of cell wall degradation hypotonic smear method. The somatic chromosome numbers of all accessions were non-constant and the amplitude was 6-11. The somatic chromosome karyotypic types of F1 progeny for 3 crosses were 2n=94=91 m+3 sm, 2n=98=88 m+10 sm(1SAT),and 2n=92=87 m+5 sm,respectively. The “n+n”transmission and 2B karyotype of F1 progeny from sugarcane and intergeneric hybrid were indicated. The chromosomes of sugarcane and intergeneric hybrid complex(Erianthus arundinaceus×Saccharum spontaneum)were transmitted in the mode “n+n”but doubling possibly existed. The karyotypes of these three F1 progenies belong to the primitive 2B type.
Key words Sugarcane;Intergeneric hybrid complex;Karyotype analysis;Chromosome transmission
doi 10.3969/j.issn.1000-2561.2016.02.002
現代甘蔗品種由于原始親本有限,長期的雜交或定向選擇,致使育成品種難以獲得突破[1-2]。通過遠緣雜交導入野生種質血緣或優異新基因是甘蔗突破性育種的重要途徑。利用斑茅和割手密雜交(Erianthus arundinaceus×Saccharum spontaneum)獲得的野生雜種F1材料斑茅割手密復合體GXAS07-6-1兼具雙親的優點,其抗逆性、生勢等具有明顯的超親優勢[3],再利用斑割復合體GXAS07-6-1與甘蔗進行雜交或回交,改變育種者幾十年來對斑茅和割手密野生種質“單元化”雜交利用的策略,有可能同時聚合斑茅和割手密的優異野生基因,以改良現有甘蔗品種,提高品種的綜合性狀水平。
甘蔗是高度雜合的異源多倍體或非整倍體植物,同一品種體細胞的染色體數目在不同個體及同一個體不同細胞之間有所差異[4];同時,由于甘蔗的染色體組成及傳遞方式較為復雜和獨特,在甘蔗遠緣雜交過程中,雙親的染色體在后代材料中的傳遞方式存在“不平衡現象”[5]。鄧祖湖等[6]對斑茅、Badila 及甘蔗與斑茅的雜交后代進行染色體計數與核型分析,結果表明,海南斑茅92-105和92-77核型屬于原始的1A型,參試的甘蔗無性系的核型均屬2B型,甘蔗斑茅雜交的染色體遺傳以“n+n”的方式進行。王先宏等[7]利用GISH技術對16份甘蔗與蔗茅屬間的雜交后代F1染色體進行研究,并對其中3份材料的中期染色體進行核型分析,結果表明子代材料中具有7~10條不等的父本染色體,甘蔗×蔗茅雜種后代F1的染色體組成為“n+n”及“2n+n”,3份F1材料核型類型分別為2C、2C和2B。
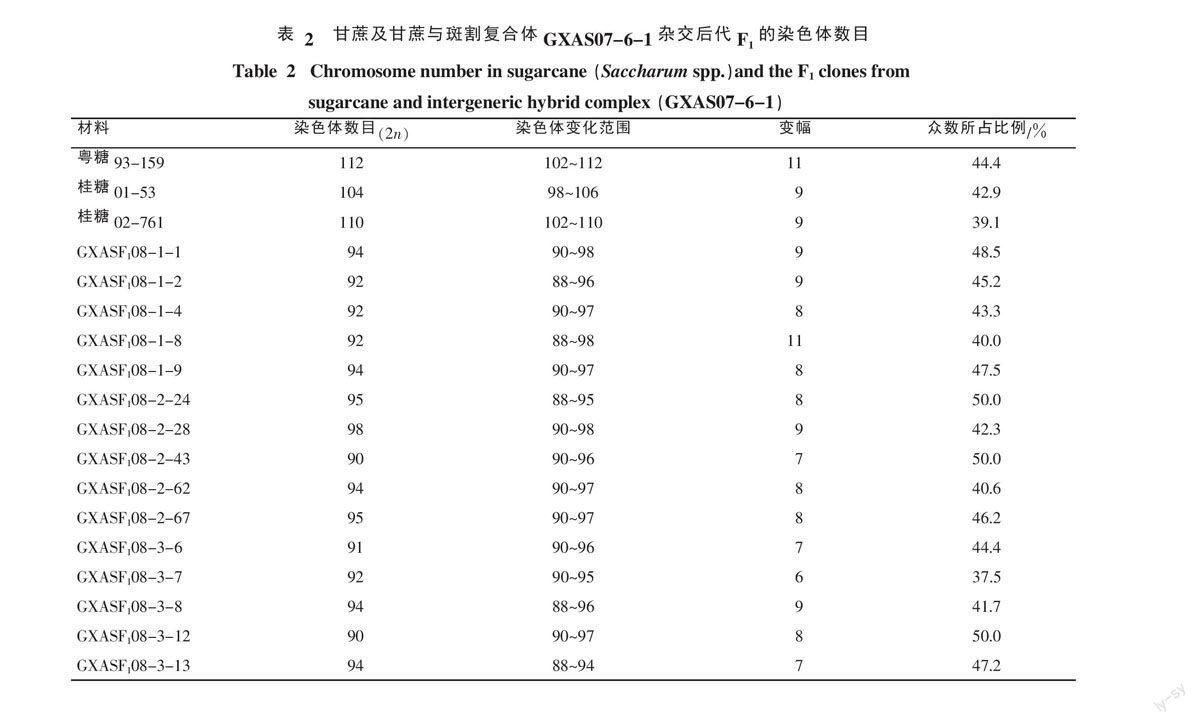
本課題組于2006年通過利用廣西斑茅87-36(2n=60)和廣西割手密79-9(2n=64)雜交獲得斑割復合體GXAS07-6-1(2n=62)以來,對GXAS07-6-1與甘蔗雜交利用過程中的分子遺傳學和細胞遺傳學相關基礎進行了研究,并獲得一批聚合斑茅和割手密雙親優良性狀(基因)的雜種后代F1[8-10]。為了有效地利用斑割復合體改良甘蔗品種,需要了解斑茅割手密染色體在雜交利用過程中的傳遞動態和遺傳規律。本研究對3個甘蔗品種(系)粵糖93-159、桂糖01-53、桂糖02-761及其與斑茅割手密復合體(GXAS07-6-1)的15個雜交后代F1材料進行染色體數目鑒定,并對其中3個F1(GXAS F108-1-1、GXAS F108-2-28、GXAS F108-3-7)進行核型分析,以期探討斑割復合體在雜交利用過程中的染色體傳遞動態和遺傳規律,為甘蔗野生資源斑茅與割手密的復合雜交利用提供細胞遺傳學的理論基礎。
1 材料與方法
1.1 材料
以斑茅割手密復合體GXAS07-6-1為父本,分別與3個不含斑茅及廣西割手密血緣的常用甘蔗親本粵糖93-159(廣州甘蔗糖業研究所選育,含熱帶種、爪哇割手密、印度割手密血緣)、桂糖01-53(廣西甘蔗研究所選育的品系,含熱帶種、爪哇割手密、印度割手密血緣)、桂糖02-761(廣西甘蔗研究所選育,含熱帶種、爪哇割手密、印度割手密、云南割手密及大莖野生種血緣)進行雜交,雜種的真偽經SRAP和SSR分子標記鑒定[9],從中選取真雜種F1(GXASF1)15份(表1)進行染色體計數和核型分析。
1.2 方法
染色體觀察與計數及核型分析。染色體制片參考黃東益等[11]、劉昔輝[8]等的方法,略有修改。將帶雙芽的莖段在28 ℃條件下恒溫培養,待其根長到2 cm,取幼嫩、健壯的根尖,置于0.002 mol/L 8-羥基喹啉溶液中預處理3~5 h (室溫),后轉入固定液(甲醇 ∶ 冰乙酸=3 ∶ 1)于4 ℃冰箱固定12 h 以上,低滲30~60 min,用70%的酒精于4 ℃保存備用;將根尖置于3%纖維素酶和1%果膠酶的混合液中酶解6~9 h(酶解時間視根尖的粗細老嫩而定),用改良苯酚卡寶品紅溶液染色10~15 min,于空氣中干燥,在尼康正立熒光顯微鏡80i下觀察,每個材料取30個分散好的完整細胞,以眾數進行染色體計數。
1.3 數據處理
用Image-Pro Plus 5.51(Media Cybernetics, Inc.)軟件測量染色體長度,Microsoft Excel 2010 軟件分析作圖,繪制核型模式圖。染色體核型參數分析參考李懋學等[12]的標準。甘蔗為異源多倍體,染色體不進行同源配對。染色體核型排列按從大到小、短臂向上、長臂向下、著絲點在一條線上進行排列。
2 結果與分析
2.1 甘蔗及甘蔗×斑割復合體后代F1材料的染色體數目和傳遞方式推斷
從表2看,各親本及其后代材料的體細胞染色體數目在不同細胞中均不恒定。粵糖93-159染色體2n=112(眾數,下同),染色體變化范圍為102~112,變幅為11條,眾數所占比例為44.4% ;桂糖01-53染色體2n=104,染色體變化范圍為98~106,變幅為9條,眾數所占比例為42.9%;桂糖02-761染色體2n=110,染色體變化范圍為102~110,變幅為9條,眾數所占比例為39.1%。粵糖93-159×GXAS07-6-1后代GXASF108-1-1、GXASF108-1-2、GXASF108-1-4、GXASF108-1-8、GXASF108-1-9染色體數目為92~94,同一材料不同細胞染色體變幅為8~11條,眾數所占比例為40.0%~48.5%;桂糖01-53×GXAS07-6-1后代中GXASF108-2-24、GXASF108-2-28、GXASF108-2-43、GXASF108-2-62、GXASF108-2-67染色體數目為90~98,同一材料不同細胞染色體變幅為7~9條,眾數所占比例為40.6%~50.0%;桂糖02-761×GXAS07-6-1后代中GXASF108-3-6、GXASF108-3-7、GXASF108-3-8、GXASF108-3-12、GXASF108-3-13染色體數目為90~94,同一材料不同細胞染色體變幅為6~9條,眾數所占比例為37.5%~50.0%。15份甘蔗與斑割復合體的后代F1染色體眾數都為90~98條,平均為93,子代的體細胞染色體數目大于雙親各自單倍的配子體染色體數目之和(87、83、86),由此推測甘蔗斑割染色體基本按“n+n”方式傳遞。
2.2 3份甘蔗與斑割復合體的后代F1材料核型分析結果
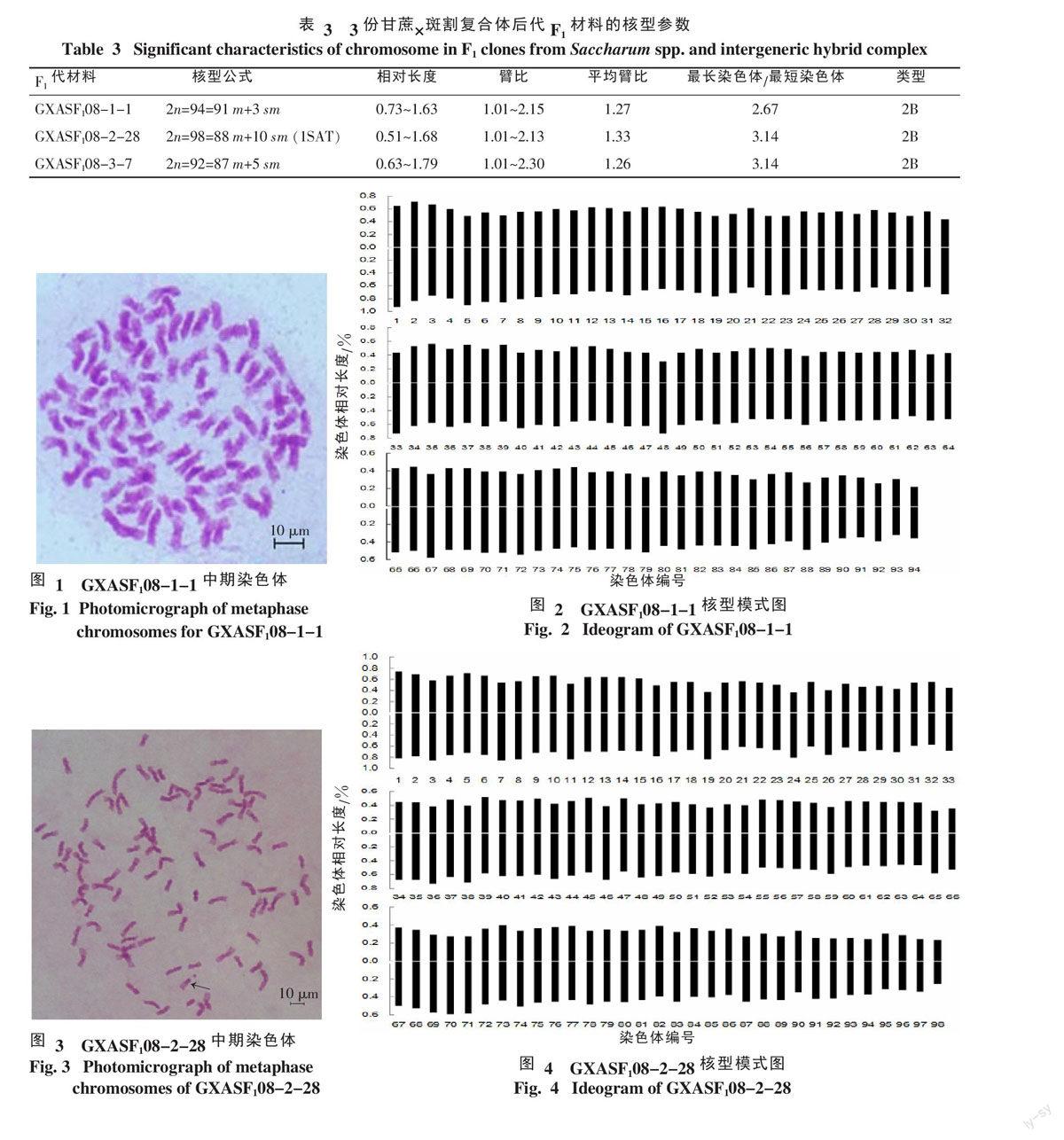
2.2.1 GXASF108-1-1的核型分析結果 GXASF108-1-1的核型參數見表3,其體細胞染色體數為2n=94,染色體相對長度0.73~1.63,臂比為1.01~2.15,平均臂比為1.27,最長染色體/最短染色體為2.67,核型分類為2B,核型不對稱系數為55.5。94條染色體中有91條為中部著絲粒染色體,3條為近中部著絲粒染色體,核型公式為2n=94=91 m+3 sm。染色體形態和核型模式見圖1、2。
2.2.2 GXASF108-2-28的核型分析結果 GXASF108-2-28的核型參數見表3,其體細胞染色體數為2n=98,染色體相對長度為0.51~1.68,臂比為1.01~2.13,平均臂比為1.33,最長染色體/最短染色體為3.14,核型分類為2B,核型不對稱系數為56.2。98 條染色體中有88條為中部著絲粒染色體,10條為近中部著絲粒染色體,90號染色體有一條隨體(圖3箭頭所示),核型公式為2n=98=88 m+10 sm(1SAT)。染色體核型模式見圖4。
2.2.3 GXASF108-3-7的核型分析結果 GXASF108-3-7的核型參數見表3,其體細胞染色體數為2n=92,染色體相對長度為0.63~1.79,臂比為1.01~2.30,平均臂比為1.26,最長染色體/最短染色體為3.14,核型分類為2B,核型不對稱系數為55.1。92 條染色體中有87條為中部著絲粒染色體,5條為近中部著絲粒染色體,核型公式為2n=92=87 m+5 sm。染色體形態和核型模式見圖5、圖6。
3 討論與結論
現代甘蔗品種由于親本遺傳基礎狹窄,長期雜交或是定向選擇,育成品種異質性低。通過遠緣雜交引進野生種質優異基因、擴寬甘蔗基因庫血緣成為育種家的共識。由于熱帶種和斑茅遠緣雜交獲得的F1大多無花粉或花粉量少且不育,其回交后代蔗糖分改良相對較慢,而且伴有側芽多、氣根多、難脫葉、蒲心大、57號毛群發達等缺點[13-14],所以盡管通過甘蔗斑茅種質的研究與利用已獲得BC4代材料,但至今尚未育成含斑茅血緣的甘蔗品種,即可供生產或育種應用的品種[15]。通過以割手密為載體或橋梁親本,與斑茅雜交獲得生勢好、莖蒲心程度相對較輕、無57號毛群、莖徑大、莖汁錘度高,花粉育性高的聚合雙親優點的斑茅割手密復合體材料,再利用該復合體與甘蔗雜交或回交,就有可能創造出新的突破性優良種質。
由于甘蔗遺傳背景復雜,染色體傳遞呈現多種形式,有“n+n”,“n+2n”,“2n+n”和“2n+2n”等方式,且甘蔗遠緣雜交中,減數分裂過程染色體配對異常,出現單價體、三價體、落后染色體以及微核等現象。鄭成木[4]的研究認為,甘蔗細胞染色體數目的變化很可能是由于祖親(不同物種)的細胞周期不同,因而在雜種中來自不同物種的染色體的行為不同步,導致細胞分裂過程中染色體分配不均衡。王先宏等[16]對甘蔗與蔗茅的雜交后代F2、BC1進行染色體遺傳分析,其結果也顯示各材料的體細胞染色體數目均不恒定,變幅為3~7條;雙親染色體在F2子代以“2n+n”方式傳遞,而在F2BC1子代則以“n+n”方式傳遞。本研究中GXAS07-6-1雙親分別是不同屬的甘蔗野生種質廣西斑茅GXA87-36(2n=60)和廣西割手密GXS79-9(2n=64),染色體按“n+n”方式傳遞[8]。粵糖93-159、桂糖01-53、桂糖02-761 3個母本的染色體數分別為112、104、110條。15份甘蔗與斑割復合體的雜交后代F1材料中,同一組合的不同后代染色體數目存在一定差異,染色體數目都為90~98條;F1代的體細胞染色體數目比雙親各自單倍的配子體染色體數目之和多,染色體按不嚴格的“n+n”方式傳遞,由此推斷,同時很可能存在部分染色體加倍現象。Bremer[17]的研究也表明,雜種中的染色體數目比預期的雙親各自單倍的配子體染色體數目之和要多。
染色體核型分析能夠了解分類關系、遺傳變異和物種的進化起源。本研究對3份甘蔗與斑割復合體的F1后代GXASF108-1-1、GXASF18-2-28、GXASF18-3-7進行核型分析,發現其染色體由大多數中部著絲點和少數近中部著絲點組成,都屬于2B型。劉文榮等[18]、鄧祖湖等[6]、黃永吉等[19]對甘蔗與斑茅雜交后代的核型分析結果表明,其F1材料的核型主要屬于2B型。Stebbins[20]認為,高等植物核型進化的基本趨勢是由對稱向不對稱發展的,較原始的植物核型具對稱的中部著絲點染色體較多,而較進化的植物核型具中部著絲點染色體較少,因此,3份甘蔗與斑割復合體的F1染色體較原始。由核型的不對稱系數來看,GXASF108-1-1的核型不對稱系數為55.5,GXASF108-2-28的核型不對稱系數為56.2,GXASF108-3-7的核型不對稱系數為55.1。3個材料的核型不對稱系數相差不大,但GXASF108-2-28較GXASF108-1-1,GXASF108-3-7不對稱,這可能與其母本的進化程度相關。
參考文獻
[1] Lu Y H, D'hont A, Walker D, et al. Relationships among ancestral species of sugarcane revealed with RFLP using single copy maize nuclear probes[J]. Euphytica, 1994, 78(1-2): 7-18.
[2] Irvine J E. Saccharum species as horticultural classes[J]. Theoretical and Applied Genetics, 1999, 98(2): 186-194.
[3] 張革民, 劉昔輝, 方鋒學, 等. 斑茅割手密復合體創制及其與甘蔗雜交F1的染色體計數和形態表現[C]//2009年中國作物學會學術年會論文摘要集. 2009.
[4] 鄭成木. 甘蔗核型及其染色體數目變化的研究[J]. 熱帶作物學報, 1993, 14(2): 47-51.
[5] Bremer G. A cytological investigation of some cultivated kinds and of their parents[J]. Genetica, 1924, 6: 497-525.
[6] 鄧祖湖, 李玉蟬, 劉文榮, 等. 甘蔗和斑茅遠緣雜交后代的染色體遺傳分析[J]. 熱帶作物學報, 2007, 28(3): 62-67.
[7] 王先宏, 楊清輝, 李富生, 等. 基于GISH的甘蔗與蔗茅屬間雜交F1后代染色體組成及核型分析[J].中國農業科學, 2011, 44(6): 1 085-1 091.
[8] 劉昔輝, 方鋒學, 高軼靜, 等. 斑茅割手密雜種后代真實性鑒定及遺傳分析[J]. 作物學報, 2012, 38(5): 914-920
[9] 高軼靜, 方鋒學, 劉昔輝, 等. 甘蔗與斑茅割手密復合體雜交后代的分子標記鑒定[J]. 植物遺傳資源學報, 2012, 13(5):912-916.
[10] 劉許輝. 甘蔗與斑茅割手密復合體雜交后代的遺傳分析[D]. 南寧: 廣西大學, 2013.
[11] 黃東益, 鄭成木, 莊南生, 等. 甘蔗染色體組構成系統演化的研究[J]. 熱帶作物學報, 2000, 21(1): 43-51.
[12] 李懋學, 陳瑞陽. 關于植物核型分析的標準化問題[J]. 武漢植物學研究, 1985, 3(4): 297-302.
[13] 劉少謀, 符 成, 陳勇生. 近十年海南甘蔗育種場斑茅后代回交利用研究[J]. 甘蔗糖業, 2007(2): 1-6.
[14] 王麗萍, 蔡 青, 范源洪, 等. 甘蔗(Saccharum)與斑茅(Erianthus arundtnaceus)遠緣雜交利用研究[J]. 西南農業學報, 2007, 20(4): 721-726.
[15] 吳嘉云. 甘蔗與斑茅后代染色體遺傳分析及抗性初步評價[D]. 福州: 福建農林大學, 2013.
[16] 王先宏, 李富生, 何麗蓮, 等. 甘蔗與蔗茅雜交F2BC1代的染色體遺傳分析[J]. 中國農學通報, 2014, 30(15): 81-85.
[17] Bremer G. Problems in breeding and cytology of sugarcane[J]. Euphytica, 1961, 10(1): 59-78
[18] 劉文榮, 鄧祖湖, 張木清, 等. 甘蔗斑茅的雜交利用及其雜種后代鑒定系列研究Ⅲ. 甘蔗斑茅遠緣雜交后代細胞遺傳分析[J]. 作物學報, 2004, 30(11): 1 093-1 096.
[19] 黃永吉, 吳嘉云, 劉少謀, 等. 基于GISH的甘蔗與斑茅F1染色體遺傳與核型分析[J]. 植物資源遺傳學報, 2014, 15(2): 394-398.
[20] Stebbins G L. Chromosomal evolution in higher plants[M]. London: Edward Arnold, 1971: 87-89.
