蓖麻根際土壤解鉀菌的篩選鑒定及發酵條件的優化
2016-05-30 10:48:04張妙宜陳宇豐周登博起登鳳高祝芬張錫炎
熱帶作物學報 2016年12期
張妙宜 陳宇豐 周登博 起登鳳 高祝芬 張錫炎

摘 要 為了提高鉀肥的利用效率,根據解鉀菌的特性及分離方法,從藥用植物蓖麻根際土壤中分離出6株高效解鉀菌,測定并分析它們的解鉀效率。結果表明:獲得1株最高效、穩定的解鉀細菌,將其命名為MY-1。通過形態觀察、生理生化特性和16S rDNA基因序列分析,初步鑒定菌株MY-1是嗜線蟲賽雷氏菌(Serratia nematodiphila)。并對菌株MY-1的發酵條件進行優化,確定其在溫度37 ℃,時間24 h,鉀長石粉量為10 g,pH值為5,搖床轉速為250 r/min,以蔗糖和硫酸銨為碳、氮源時,發酵效果最佳,發酵液中可溶性鉀含量最高,達65.04 mg/L。菌株MY-1有穩定且較強的解鉀能力,將為今后微生物菌肥研究、開發及土壤肥力優化提供理論依據。
關鍵詞 蓖麻;根際土壤;微生物;解鉀;發酵條件優化
中圖分類號 S565.6 文獻標識碼 A
蓖麻系蓖麻屬大戟科一年或多年生草本喜溫作物,原產地非洲東部,適種于熱帶地區[1],其綜合利用價值高,是世界上十大油料作物之一,具有一定的藥用價值。它周身是寶,從葉片、莖稈到根部均可入藥,根系發達,呈圓錐形,主根入土可深達2~4 m,側根橫向分布可達1.5~2 m,耐旱性和抗貧瘠能力極強[2]。在含鉀量高的鹽堿地中,蓖麻的另一“耐鹽”特性能運用自身的耐鹽性機制從根部向地上部運輸,提高鉀的吸收效率,維持鉀、鈉離子的平衡[3]。
地殼礦質元素中排名第7位的鉀元素含量為2.47%,是作物生長的重要營養要素之一[4],對促進作物生長發育、提高代謝能力、增強酶活性和作物光合作用,以及增強作物抗病、抗凍、抗旱、抗鹽等能力起重要作用[5]。然而鉀在土壤中有95%是以礦物鉀的形態存在于鉀長石和云母這兩大礦物中,無法被作物直接吸收利用,在農業生產過程中使用化肥鉀肥補充作物生長所需的鉀元素,見效雖快,卻破壞了土壤結構、造成有機質含量逐年下降[6],不利于生態平衡與農業生產持續健康穩定的發展。
硅酸鹽細菌(Silicate bacteria)俗稱“解鉀菌”,大量存在于土壤中,1912年被Bassalik首次分離于蚯蚓腸道中,1939年被科研人員從土壤中分離并被正式命名,1950年其釋放鋁硅酸鹽礦物中鉀元素的能力被前蘇聯學者亞歷山大羅夫發掘[7],之后人們開始嘗試研究利用其改善土壤中可溶性鉀嚴重缺失的狀況。解鉀菌能分解土壤中鉀長石等不溶性無機礦物質,將其轉化為可供作物直接吸收利用的有效鉀,溶磷固氮、改善作物氮磷營養[8],促進作物生長,改良土壤環境。
近年來更有研究結果表明,根際解鉀菌比普通解鉀菌更能產生對土壤微生物活性有促生作用的分泌物[9]。根際土壤解鉀菌通過刺激作物生長激素分泌,增強作物對環境的脅迫耐性和對病原菌的抗病性[10]。史靜靜[11]從棉花根際分離篩選出5株(K1111、K1114、K2115、K3105和K3205)具有親和性的促生效果明顯的優良PGPR菌株;羅娜等[12]從降香黃檀、檀香根際土壤中分離篩選出解鉀率高達221.18%的惡臭假單胞菌JT-K21。茍志輝[13]從油茶根際分離純化出73株解鉀細菌,其中菌株K50的活性達121.71 mg/kg;陳宇豐[14]從香蕉根際土壤中篩選出16株高效解鉀菌,并最終獲得解鉀率約為20%的陜西鏈霉菌(Streptomyces shaanxiensis)M3-4。但受氣候、土壤類型、作物品系等綜合因素的影響,作物根際土壤解鉀菌的解鉀能力在一定程度上受到制約。
本研究以鉀長石粉為唯一鉀源對蓖麻根際土壤進行解鉀細菌的分離與篩選,獲得6株解鉀能力較強的細菌,選取其中1株抗逆性較好,生長能力較強,且解鉀能力最佳的解鉀細菌MY-1作為研究對象,通過觀察菌株的形態、生理生化特征,分析16S rDNA序列;優化發酵條件,測定該解鉀菌株的解鉀能力,為研究開發具有穩定高效解鉀能力的作物根際解鉀菌提供理論基礎,以緩解當前土壤“少鉀”現狀。
1 材料與方法
1.1 材料
1.1.1 樣品采集 土壤樣品分別采自海南省臨高縣南寶鎮、美臺鎮、皇桐鎮3地的蓖麻植株根際土壤。采用五點交叉取樣法,共采集土樣8份,分別放入無菌封口袋中混勻、封口、編號,裝入冰盒內保存后,除去根系、石塊等雜物,4 ℃保存備用。
1.1.2 儀器設備 超凈工作臺:SW-CF-1F,蘇州蘇潔凈化設備有限公司;生化培養箱:SPX-150,北京恒瑞天創機電設備有限公司;冷凍離心機:ST16R,德國Thermo公司;PCR儀:9701,賽飛(中國)有限公司;水平電泳槽:HR/022,北京恒瑞天創機電設備有限公司;凝膠成像儀:BG-gds Auto,北京百晶生物技術有限公司;恒溫水浴鍋:HHS-11-2,杭州匯爾儀器;紫外可見分光光度計:UV1000,上海天美科學儀器有限公司。
1.1.3 主要試劑 分析純試劑:磷酸二氫鉀、硫酸鎂、氯化鈉、無水硫酸鈣、碳酸鈣、磷酸氫二鈉、氯化鐵、蔗糖等;生化試劑:酵母粉、胰蛋白胨、瓊脂、四苯硼鈉等。
1.1.4 培養基 分離培養基:葡萄糖10.0 g,碳酸鈣5 g,磷酸二氫鉀0.2 g,硫酸鎂0.2 g,無水硫酸鈣0.2 g,氯化鈉 0.2 g,瓊脂15 g,去離子水1 000 mL,pH7.2;篩選培養基:蔗糖5 g,磷酸氫二鈉2 g,硫酸鎂0.5 g,碳酸鈣0.1 g,氯化鐵0.005 g,土壤礦物1 g,瓊脂20 g,蒸餾水1 000 mL,pH7.0~7.5;LB營養培養基:酵母粉5 g,胰蛋白胨10 g,氯化鈉l0 g,瓊脂20 g,去離子水1 000 mL,pH7.2~7.5。
1.2 方法
1.2.1 根際土壤解鉀菌的分離、篩選 (1)初篩:將新鮮土樣(10 g)放入無菌水(90 mL)中,利用磁力攪拌器充分混合均勻后,在無菌操作條件下,采用稀釋涂布平板法分別配制10-4、10-5、10-6三個濃度梯度的土壤懸液備用,各吸取100 μL懸液涂布于分離培養基上(每個梯度設3個重復),37 ℃下培養1~3 d,觀察細菌菌落生長和形態特征,運用平板劃線法純化直至獲得純培養。
(2)復篩:將初篩獲得的純化菌株轉接到以鉀長石為唯一鉀源的篩選培養基上,37 ℃下培養2 d,采用四苯硼鈉滴定法測定鉀,菌落周圍呈現亮藍色,初步確定含有解鉀細菌,將其純化得到6種菌株,分別編號MY-1、MY-2、MY-3、MY-4、MY-5、MY6,并保存至LB斜面培養基上擴大培養。
(3)解鉀能力測定:將測定取出的培養液樣品倒入蒸發皿中,用恒溫水浴鍋干燥蒸餾至10 mL,加入4 mL H2O2繼續蒸發除去菌體及殘渣至無粘稠物,4 ℃,4 000 r/min,離心10 min,收集上清液至容量瓶(50 mL)中定容。同時設置空白對照(未接種),每個處理重復3次,利用火焰原子吸收光度計測定鉀含量(mg/L)。
1.2.2 解鉀菌的鑒定 (1)菌株形態學和生理生化鑒定:將保藏處理的菌種活化后,37 ℃下培養1~3 d,觀察其菌落形態特征,然后挑取菌株分別進行細菌革蘭氏、莢膜、芽孢染色鏡檢觀察,并參考《常見細菌鑒定手冊》[15]、《伯杰氏細菌鑒定手冊》[16]對菌株進行生理生化測定。
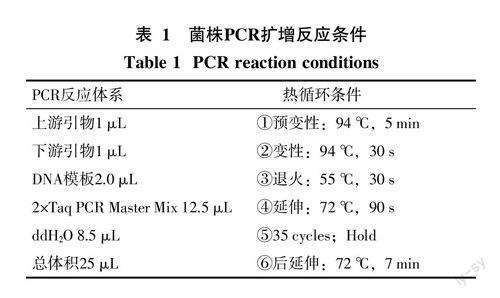
(2)菌株16S rDNA分子生物學鑒定:將活化后的菌種接入LB液體培養基中,選用16S rDNA通用引物27F(5′-AGAGTTTGATCCTG-GCTCAG-3′)和1492R(5′-GGTTACCTTGTTACGACTT-3′),建立PCR擴增體系進行擴增(表1)。
擴增處理后的DNA樣品,經1%瓊脂糖凝膠電泳預檢測后,送往北京六合華大基因股份有限公司純化測序,獲得DNA序列后輸入GenBank,用Blast程序與數據庫中的所有序列進行比對,使用鄰接法Neighbor Joining method[17]選取最相似的模式菌株作為參比對象,運用MEGA5.05軟件構建系統發育樹。
1.2.3 解鉀細菌發酵條件優化 解鉀菌自身的遺傳特性、種類成分決定了它對難溶性鉀礦石有高效的解鉀作用,為盡可能地使菌株MY-1的解鉀量達到最大化,本研究通過優化菌株的培養時間、pH值、鉀長石粉量、搖床轉速、碳源種類、氮源種類共6個因素,設定每個因素適合的水平數,并通過單因素方差設計,分析在不同培養條件下解鉀菌MY-1的解鉀量變化,得出MY-1的最佳發酵條件。
(1)培養時間對解鉀量的影響:在LB培養基上將已篩選出的解鉀菌株培養24 h,然后接種到無菌水中,置于搖床上充分震蕩,制成懸液備用;另配制解鉀培養基(無鉀),取100 mL分裝于已加有1 g鉀長石粉的三角瓶(250 mL)內,121 ℃,25 min滅菌后,接入1%接種量的備用懸液,37 ℃,150 r/min,分別在培養12、24、36、48、60、72 h后取樣。
(2)pH值對解鉀量的影響:配制解鉀菌備用懸液和無鉀解鉀培養基(步驟同上),調節培養液pH值分別為3、4、5、6、7、8、9,培養2 d后取樣。
(3)鉀長石粉量對解鉀量的影響:配制解鉀菌備用懸液和無鉀解鉀培養基(步驟同上),分別加入0.5、1、1.5、2.5、5、10 g的鉀長石粉,培養2 d后取樣。
(4)搖床轉速對解鉀量的影響:配制解鉀菌備用懸液和無鉀解鉀培養基(步驟同上),分別置于50、100、150、200、250 r/min轉速的搖床上振蕩,培養2 d后取樣。
(5)不同碳源種類對解鉀量的影響:配制解鉀菌備用懸液和無鉀解鉀培養基(步驟同上),分別以葡萄糖、蔗糖、乳糖、果糖、淀粉、麥芽糖為碳源,培養2 d后取樣。
(6)不同氮源種類對解鉀量的影響:配制解鉀菌備用懸液和無鉀解鉀培養基(步驟同上),分別以氯化銨、硝酸銨、硫酸銨、硝酸鈉、酵母粉、蛋白胨為氮源,培養2 d后取樣。
1.3 數據處理
采用WPS和SAS9.1統計軟件DUI對實驗數據進行分析及多重比較。
2 結果與分析
2.1 根際土壤解鉀菌的篩選
本研究對蓖麻根際土壤樣品進行涂布處理,初步篩選得到83株菌,通過分離、初篩等步驟挑取6株生長效率高、具高效解鉀能力的菌株,再經過優化發酵條件、采用過氧化氫灰化法[18]等進一步地優化鑒定菌株,并使用火焰原子吸收分光光度計測定這6株解鉀細菌的可溶性解鉀含量,得出其解鉀能力均較強。其中接種MY-1菌株的培養液所產生的速效鉀含量最高,解鉀效果較其他菌株更加明顯,因此,確定以MY-1菌株為實驗研究對象,進行生理生化特性鑒定和發酵條件優化,分析其解鉀特性的變化為后續研究奠定基礎。
2.2 解鉀菌MY-1的生理生化鑒定
由表2可知,菌株能使明膠液化,接觸酶、硝酸鹽還原、甲基紅等試驗呈現陽性;不能水解淀粉、纖維素,丙二酸、乙酰甲基甲醇、硫化氫等試驗呈現陰性。
在碳源利用方面:菌株MY-1可利用α-乳糖、D-纖維二糖、D-果糖、D-半乳糖、D-葡萄糖、D-甘露糖、D-山梨醇、D-海藻糖、D-木糖、L-阿拉伯糖、L-苯丙氨酸、棉子糖、木聚糖、肌醇、松三糖、鼠李糖、水楊苷、可溶性淀粉和蔗糖,但不能利用核糖。
在氮源利用方面:菌株MY-1可利用L-精氨酸、L-苯基丙氨酸、甘氨酸、L-乙硫氨酸、纈氨酸、組氨酸和乙酸銨作為唯一氮源,但不能利用L-絲氨酸、蛋氨酸、L-羥基脯氨酸、L-半胱氨酸、硝酸銨、氯化銨、硫酸銨、草酸銨和四水合鉬氨酸。
菌株MY-1的最適生長溫度為37 ℃,pH值為5;且能生長在NaCl含量小于10%的培養基上。
2.3 菌株16S rDNA分子生物學鑒定
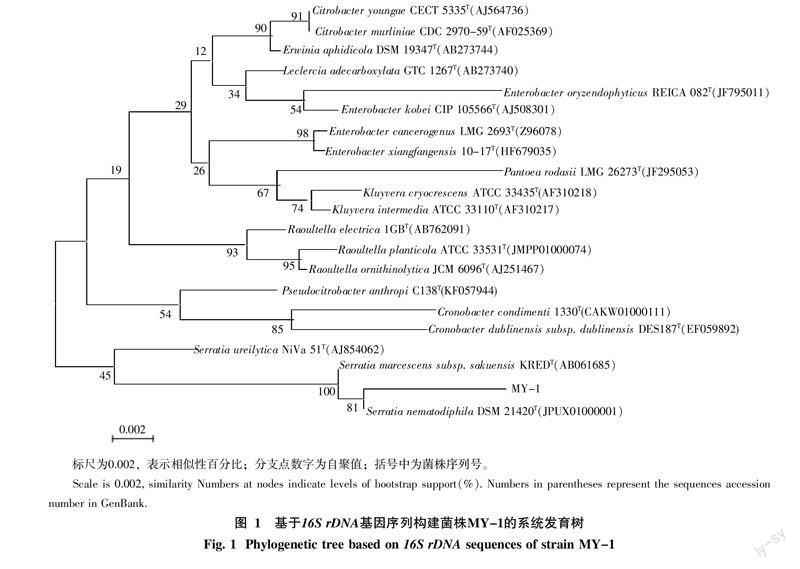
對菌株MY-1進行測序,獲得1 530 bp的序列片段,將菌株的序列分別在GenBank和EzBioCloud數據庫中進行同源性比對,并選取20株同源性較高的標準菌株序列,與靶標菌株MY-1的基因序列一起,應用MEGA 5.1軟件的相關功能構建鄰接距離矩陣法系統發育關系進化樹(圖1)[19]。結果顯示,菌株MY-1與鏈霉菌屬(Streptomyces)的同源性均較高,大多數的序列聚集在同一節點處,并且具有較近的遺傳距離。MY-1與嗜線蟲賽雷氏菌Serratia nematodiphila DSM 21420T(JPUX01
000001)和粘質沙雷氏菌Serratia marcescens subsp.sakuensis KREDT(AB061685)親緣關系最近,相似率均達100%。根據系統發育樹相似性和同源性分析結合形態特征、培養特征和生理生化特征,鑒定此菌株為嗜線蟲賽雷氏菌(Serratia nematodiphila)。
2.4 解鉀菌MY-1發酵條件的優化研究
2.4.1 不同培養時間對解鉀菌解鉀量的影響 由圖2可知,在72 h內,菌株MY-1對鉀長石的解鉀能力變化顯著。當培養時間從12 h增加至36 h時,解鉀量總體呈上升趨勢,并在36 h時達到峰值,解鉀量為44.57 mg/L;在36 h后,48、60、72 h的解鉀量略有下降并保持較平穩狀態,分別為34.95、35.61、34.62 mg/L。這表明菌株MY-1在培養了36 h后的解鉀能力最強,在36 h后因受代謝產物的影響,其解鉀能力略有降低但基本處于平穩狀態。這對今后開發具有穩定解鉀能力的菌肥生產具有十分重要的意義。
2.4.2 不同pH值條件對解鉀菌解鉀量影響 由圖3可知,調節培養pH值可使菌株MY-1的解鉀能力發生顯著變化.設定pH值從3~9,菌株解鉀能力總體為先升后降,在pH為5時達到峰值,解鉀量為31.30 mg/L,比pH3時的解鉀量飆升了40.32%;將pH值調至6,開始明顯下降,降至8.72 mg/L;并在pH值為7、8時保持穩定,均為7.06 mg/L;當pH9時解鉀量降至最低值4.74 mg/L。這表明改變pH值對菌株MY-1的解鉀量變化有顯著影響,菌株MY-1最適合酸性環境下生長,這為改良土壤肥力提供有利依據。
2.4.3 不同鉀長石粉量對解鉀菌解鉀量影響 由圖4可知,在鉀長石粉量為0.5、1.0 g時,解鉀量變化甚小,分別為42.58、46.23 mg/L;當增加到1.5、2.5、5.0 g時,菌株解鉀能力呈穩定增加趨勢,解鉀量分別為73.46、94.04、112.30 mg/L;當加入10.0 g的鉀長石粉時,菌株的解鉀能力最強,解鉀量為183.01 mg/L。這表明菌株MY-1的解鉀能力隨著鉀長石粉量的增加而遞增,不溶性鉀礦石含量越高,解鉀能力越高,這對充分利用土壤鉀元素有著重要的意義。
2.4.4 不同搖床轉速對解鉀菌解鉀量的影響 由圖5可知,從50~250 r/min為觀察周期,搖速每間隔50 r/min提取1次培養液測定其解鉀量,在50、100、150、200 r/min時,解鉀量穩定增加,分別為36.61、50.88、55.53、63.17 mg/L;當搖床轉速為250 r/min時,菌株解鉀量達到峰值為63.50 mg/L,比轉速為200 r/min時略高,但基本保持恒定。這表明不同搖床轉速對菌株MY-1的解鉀能力產生顯著影響,搖速增大時,解鉀量也隨之增加,并在250 r/min轉速時,解鉀量最佳。
2.4.5 不同碳源種類對解鉀菌解鉀量的影響 由圖6可知,不同碳源種類對菌株解鉀量影響的強弱順序為:蔗糖>葡萄糖>乳糖>果糖>淀粉>麥芽糖。以蔗糖為碳源時,接種了菌株MY-1的培養液解鉀量最大,達32.62 mg/L,其次是葡萄糖;以麥芽糖碳源時,解鉀效果最差,解鉀量降至1.56 mg/L。這表明該解鉀菌發酵培養時,以蔗糖、葡萄糖、乳糖為碳源,發酵效果最好,解鉀量最大,其中蔗糖是發酵時的最佳碳源。
2.4.6 不同氮源種類對解鉀菌解鉀量的影響 由圖7可知,不同的氮源種類對解鉀菌解鉀量的影響強弱順序為:硫酸銨>氯化銨>硝酸銨>硝酸鈉>酵母粉>蛋白胨,以硫酸銨為氮源時,解鉀量達42.0 mg/L,以蛋白胨為氮源時,解鉀效果最差,解鉀量為2.33 mg/L。這表明解鉀菌MY-1發酵培養時,對氮源的利用以有機氮為主,有機氮利用效率優于無機氮,且以硫酸銨為氮源,發酵效果最好。
3 討論
解鉀菌能分解鉀長石、云母等不溶硅鋁酸鹽類無機礦物,大量存在于土壤中,并將土壤中的固態鉀分解轉化為可溶性速效鉀。已有大量研究結果表明,從作物根際土壤中分泌出來的解鉀菌更能產生對土壤微生物活性有促生作用的分泌物,劉璇等[20]從青島即墨煙草根際土壤中篩選到的8株解鉀菌并最終確定其中1株變棲克雷伯氏菌(Klebsiella variicola)產生的速效鉀能力最強;張朝輝等[21]從環神農架煙區烤煙根際土壤分離出1株使鉀有效性提高約27%的解鉀菌K03;羅華元等[22]從云煙高端品牌直煙區根際土壤中分離出77株解鉀菌株,并高溫篩選獲得1株解鉀活性達10.6 mg/L的側孢芽孢桿菌(Bacillus Lateraporus)K77解鉀菌。目前以藥用植物根際土壤解鉀菌為研究對象的報道還相對較少,從慕東艷等[23]在黑龍江省6個地區栽培的藥用植物根際土壤樣品中,分離和鑒定出真菌35屬86種的試驗可以看出,藥用植物根際土壤的菌類多樣性可研究性。油料作物蓖麻,具有很高的藥用價值,蓖麻根醇提物具有體外抗乙肝病毒和抑菌作用[24],另一方面,蓖麻對土壤重金屬污染修復和微生物的強化作用也不容小覷,從蓖麻根際土壤中分離篩選出的解鉀菌具有較高的解鉀活性,可為將來的農業生產利用奠定基礎。本研究是針對蓖麻根際土壤進行試驗分析,從蓖麻根際土壤中分離得到6株高效解鉀菌,通過解鉀量的測定和菌株鑒定,利用分離篩選培養基獲得1株高效解鉀細菌MY-1,在經過發酵培養36 h后,解鉀量可達44.57 mg/L,與同類文獻相比[7],發酵周期較短,這有利于加快對土壤鉀的調控速度。通過形態觀察、生理生化試驗和16S rRNA基因序列分析對菌株進行鑒定,初步鑒定該菌株為嗜線蟲賽雷氏菌(Serratia nematodiphila),目前研究較廣泛的高效解鉀硅酸鹽細菌多為膠質芽孢桿菌B. mucilaginosus,如Karavaiko等[25]的研究,還未見從藥用作物根際土壤中分離出嗜線蟲賽雷氏菌的報道,菌株MY-1在不同的鉀長石粉量培養條件下,不溶性鉀礦石含量越高,解鉀能力越高,當加入10 g的鉀長石粉時,解鉀量可為183.01 mg/L,絕對解鉀含量值較高,這有利于提高對根際土壤鉀元素的利用率。
解鉀菌解鉀量的多少是影響菌株性能的重要因素,本研究從海南省臨高縣蓖麻種植區根際土壤中采樣篩選出1株具有高效解鉀能力的解鉀菌株MY-1,可溶性鉀含量高達65.04 mg/L,初步鑒定為嗜線蟲賽雷氏菌(Serratia nematodiphila),通過單因素發酵條件優化結合方差分析統計得出,MY-1菌株在以蔗糖和硫酸銨為碳氮源時的最佳發酵條件為:培養溫度為37 ℃,pH值為5,搖床轉速為250 r/min,培養時間為24 h,鉀長石粉量為10 g。本研究著重利用蓖麻自身能從根部向地上部運輸鉀離子的耐鹽性機制,提取根部土壤解鉀菌,并進行發酵條件優化,使該解鉀菌的解鉀量達到最大化,為今后減少工業化肥,改善植株抗逆性、提高代謝能力,平衡土壤肥力和改善土壤環境提供可行性措施,進而為今后開展有益微生物菌肥田間試驗和農業生產提供理論依據。
參考文獻
[1] 張良波, 李昌珠, 宋 煒. 能源植物系列講座(8)世界十大油料作物之一——蓖麻[J]. 太陽能, 2009(8): 17-19.
[2] 張寶賢,譚德云. 中國蓖麻的發展優勢與加快能源化利用的決策[J]. 中國農村小康科技, 2009(12): 12-16.
[3] 孫振鈞. 蓖麻產業發展:從種植到利用[J]. 中國農業大學學報,2012, 17(6), 204-214.
[4] 伍善東, 雷 平, 郭照輝, 等. 1株高效解鉀菌的分離、鑒定及培養條件優化[J]. 貴州農業科學, 2016, 44(5): 77-80.
[5] 李新新, 高新新, 陳 星, 等. 一株高效解鉀菌的篩選、鑒定及發酵條件的優化[J]. 土壤學報, 2014, 51(2): 381-388.
[6] 周毅峰, 羅云霞, 劉華中. 解鉀菌的篩選[J]. 湖北民族學院學報(自然科學版), 2009, 27(3): 285-288.
[7] 陳 易, 程永毅, 郭 濤, 等.一株具紫色土親和性解鉀菌的篩選及促生效應[J]. 西南大學學報(自然科學版), 2016, 38(5): 58-65.
[8] 陳廷偉.膠質芽孢桿菌分類名稱及特性研究(綜述)[J].土壤肥料, 2002, 5(4): 5-10.
[9] Sugumaran P, Janarthanam B. Solubilization of potassium containing minerals by bacteria and their effect on plant growth[J]. Wor J Agr Sci, 2007, 3(3): 350-355.
[10] Sturz A V, Christie B R, Nowak J. Bacterial endophy tes,Potential role in developing sustainable system of cropproduction[J]. Citical Reviews in Plant Sciences, 2000, 19(1): 1-30.
[11] 史靜靜. 棉花根際解鉀菌的生理活性和促生效果評價[D]. 合肥: 安徽農業大學, 2012.
[12] 羅 娜, 周德明, 徐 睿, 等. 降香黃檀、檀香根際解鉀菌的篩選與活性研究[J]. 熱帶作物學報, 2016, 37(5): 964-970.
[13] 茍志輝. 油茶根際功能菌株的組合優化及菌肥研究[D]. 株洲:中南林業科技大學, 2010: 1-51.
[14] 陳宇豐. 香蕉根際土壤解鉀放線菌的篩選鑒定及解鉀特性研究[J]. 生物技術通報, 2015, 31(6): 129-137.
[15] 東秀珠, 蔡妙英. 常見細菌系統鑒定手冊[M]. 北京: 科學出版社, 2001: 364-398.
[16] Buchanan R E, Gibbons N E. 伯杰細菌鑒定手冊[M]. 北京: 科學出版社, 1984.
[17] Hong K, Gao A H, Xie Q Y, et al. Actino-mycetes for marine drug discovery isolated from man-grove soils and plants in China[J]. Mar Drugs,2009,7(1):24-44.
[18] 陳華癸. 微生物學實驗[M]. 北京: 農業出版社,1962.
[19] Wang F, Xu X X, Qu Z, et al. Nonomuraea wenchangensis Sp. Nov,isolated from mangrove rhizosphere soil[J]. International Journal of Systematic and Evolutionary Microbiology, 2011, 61(7): 1 304-1 308. Doi, 10.1099/ijs.0.025742-0.
[20] 劉 璇, 孔凡玉, 張成省, 等. 煙草根際解鉀菌的篩選與鑒定[J]. 中國煙草科學, 2012, 33(3): 28-31.
[21] 張朝輝, 王豹祥, 席淑雅,等. 一株烤煙根際解鉀細菌的鑒定及其在烤煙生產中的應用[J]. 浙江農業學報, 2011, 23(3):553-558.
[22] 羅華元, 常壽榮, 王紹坤,等. 云煙高端品牌植煙區根際土壤高效解鉀菌的篩選[J]. 西南農業學報, 2011, 24(5): 1 813-1 817.
[23] 慕東艷, 呂國忠, 孫曉東,等. 黑龍江省藥用植物根際土壤真菌多樣性[J]. 生態學報, 2013, 33(1): 229-237.
[24] 唐祖年,徐雅娟,馮 梅,等. 蓖麻根提取物對HepG2 2 15HBsAg和HBeAg表達和抗菌作用的研究[J].時珍國醫國藥,2010, 21(11): 3 006-3 008.
[25] Karavaiko G I. Role of microoganisms and some physico-chemical factors of the medium in quartz destruction[J]. Mikrobiologiya, 1984, 53(6): 976-981.
