海南飛機草病原真菌鑒定、致病力測定及其寄主范圍分析
2016-05-30 13:38:11李雨龍楊葉韋誠符旖晴
熱帶作物學報 2016年1期
李雨龍 楊葉 韋誠 符旖晴

摘 要 對海南多個市縣的飛機草發病情況進行調研發現,飛機草葉斑病受自然環境及病菌種類的影響,從發病樣品上分離獲得46株病原真菌。經過形態學觀察與分子鑒定比對,確定飛機草的重要病原真菌分別為:鏈格孢Alternaria alternata、新月彎孢Curvularia lunata和尖孢鐮刀菌Fusarium oxysporu。通過離體葉片針刺法和活體接種法對病源真菌致病力進行測定,結果顯示,強致病力菌株12個(占26.09%),中等致病力菌株13個(占28.26%),弱致病力菌株19個(占41.30%),無致病力菌株2個(占4.3%),其中鏈格孢菌株LGB100501、LGB100401和彎孢菌株WB110216表現出較高的致病力。利用11個高致病力菌株針對8種海南常見雜草進行寄主范圍測試,結果表明,不同菌株致病力存在差異,僅有13.6%的供試雜草對這些菌株表現出高度和中度感病。
關鍵詞 海南;飛機草;病原真菌;致病力;寄主范圍
中圖分類號 S451.1 文獻標識碼 A
Abstract The investigation and sampling of the infection situation of Chromolaena odorata in some regions of hainan revealed that the leaf spot was influenced by natural environment. 46 pathogenic fungi were isolated. Based on morphological characteristics and sequence alignments of rDNA ITS region,the most important pathogen fungi of C. odorata were identified as Alternaria alternata, Curvularia lunata and Fusarium oxysporu. The virulence of the strains were determined by inoculation in vitro and in vivo. The results showed that the high-pathogenic strains were 12(26.09%),the medium-pathogenic were 13(28.26%),the low-pathogenic were 19(41.30%),and non-pathogenic 2(4.3%). The virulence of LGB100501, LGB100401 and WB110216 were very high. High virulence of 11 strains to 8 weeds showed that different strains had differences in virulence,highly susceptible and moderately susceptible accounted for only 13.6%.
Key words Hainan;Chromolaena odorata;Pathogenic fungi;Virulence;Host range
doi 10.3969/j.issn.1000-2561.2016.01.029
飛機草(Chromolaena odorata L)是菊科(Compositae)澤蘭屬(Eupatorium)多年生雜草,學名香澤蘭[1],是IUCN 確定的全球100種最具破壞力的入侵生物之一,該草遍布海南全島,對海南農林牧業的生產、物種多樣性和生態系統安全造成了嚴重危害。越來越多的研究表明,篩選和利用本土天敵控制入侵生物是一條簡捷安全的途徑。迄今為止,已有不少應用病原真菌防治入侵雜草的成功實例,并產生不少商品化的生物制劑[1-3]。
目前,國內對飛機草的研究主要集中在入侵生態學方面,并取得了突破性進展,而有關飛機草生物防除的相關研究工作較為落后,尤其是有關病原微生物控制與應用的研究報道較少。海南作為國內發現飛機草較早且危害最嚴重的地區,野外存在自然感病的植株,這為篩選和利用本土天敵控制入侵惡性雜草的研究工作提供了有利條件。但是,目前還沒有研發出能夠成功用于飛機草防治的真菌除草劑。已報道的飛機草致病菌主要有從葉片上分離的澤蘭尾孢Cerospora eupatorii(又名Mycovellosiella perfoliata)、 假尾孢Pseudocercospora eupatorii-formosani、鏈格孢Alternaria alternata、Anhellia niger和Colletotrichum sp.、Septoria ekmaniana等[4-6],從花序上分離到的Aureobasidium pullulans對飛機草也具有極高的生防潛力[7]。筆者于2013~2014年對海南地區的飛機草真菌病害進行調查,經分離獲得大量致病菌株,并對其展開致病力及寄主范圍的測定,對其潛在的生防價值進行初步評估。
1 材料與方法
1.1 材料
飛機草:采集自海南各個地區自然發病的飛機草;接種材料:從海南大學環植學院教學基地及其周邊采集健康無癥狀的飛機草葉片及幼苗。
PDA培養基:1 000 mL蒸餾水,200 g馬鈴薯,20 g瓊脂。
1.2 方法
1.2.1 田間調研 調查時間:2013年9~10月、2014年4~6月。調查地區:海口、東方、澄邁、臨高、儋州、昌江、樂東、三亞等市縣。調查方法:以田間、村莊等為主,以5 km為調查單位,沿途記錄飛機草種群、分布情況、生長環境等。采集發病的葉片,記錄飛機草發病癥狀及病害發生程度。
1.2.2 致病菌的分離與形態學鑒定 從野外采集自然感病的飛機草,記錄典型的發病癥狀后,采用常規的組織分離法對病菌進行分離培養、純化并將其接種到原雜草[8];確定是致病菌后再作屬或種的鑒定[9]。
1.2.3 重要病原菌的分子鑒定 待菌株在PDA培養基上培養7 d后收集菌絲,用CTAB法提取菌株DNA,PCR擴增引物為真菌ITS序列通用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)。對rDNA的ITS區域進行PCR擴增,擴增體系:Buffer(含Mg2+)2.5 μL,引物ITS1(10 μmol/L)以及ITS4(10 μmol/L)各0.5 μL,2.5 mmol/L dNTPS 1μL,Taq聚合酶0.2 μL,模板DNA 0.5 μL,用雙蒸滅菌水將體積至25 μL。PCR反應程序:94 ℃預變性4 min后,94 ℃變性45 s,55 ℃復性45 s,72 ℃延伸1 min,進行30個循環,最后72 ℃延伸10 min,于4 ℃保存。將PCR產物在瓊脂糖凝膠中電泳分離后,于EB溶液(0.5 μg/mL)中染色15 min,通過凝膠成像系統檢查電泳結果。將PCR產物送至上海生工生物技術有限公司進行測序,采用B1ast程序將其與 GenBank上的序列進行相似性比較。
1.2.4 致病力測定 (1)菌絲對離體葉片的致病力測定。根據柯赫氏法則,采用離體葉片針刺法。取健康、大小均一的飛機草葉片,先用4%次氯酸鈉溶液消毒1 min,后用無菌水沖洗3次,再用解剖針穿刺造成輕微傷口,以刺破下表皮而上表皮完好為標準;在培養7 d的菌落邊緣打菌塊并將其接在刺傷處,以無菌瓊脂塊為對照,每處理接種3個葉片,共重復3次,置于28 ℃恒溫箱中保濕培養;3 d后觀察并記錄發病的癥狀,統計發病率及進行病情分級。
病害分級標準如下: 0級為不發病;1級為僅刺傷點有侵染,病斑直徑小于1 mm;2級為傷點有侵染,病斑直徑1~5 mm;3級為傷點有侵染,病斑直徑5~10 mm;4級為傷點有侵染,病斑直徑10~15 mm;5級為傷點有侵染,病斑直徑大于15 mm。
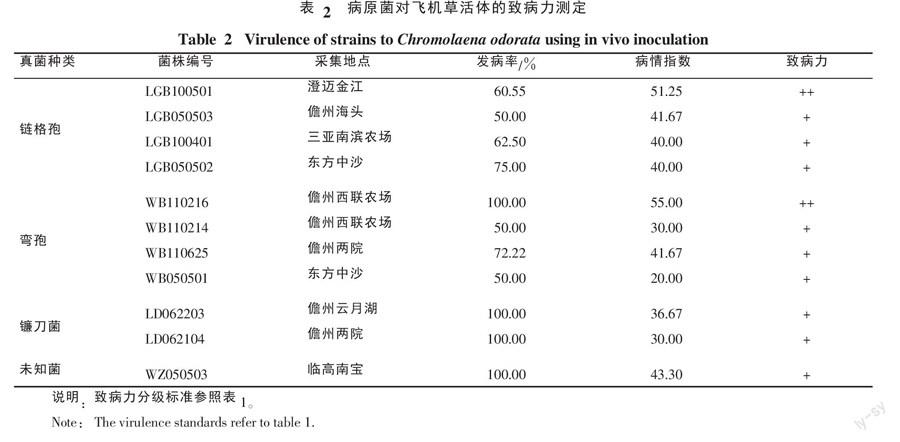
(2)菌絲對飛機草活體的致病力測定。選取11株致病力較高的菌株,采用水培法進行測定[10]。從田間采集健康、大小均一的飛機草植株,每株含4~6健康新鮮葉片,于三角瓶中扦插水培,待恢復生長24 h后接種,接種處以薄脫脂棉保濕,7 d后統計發病率和發病情況。其它方法見1.2.4(1)。
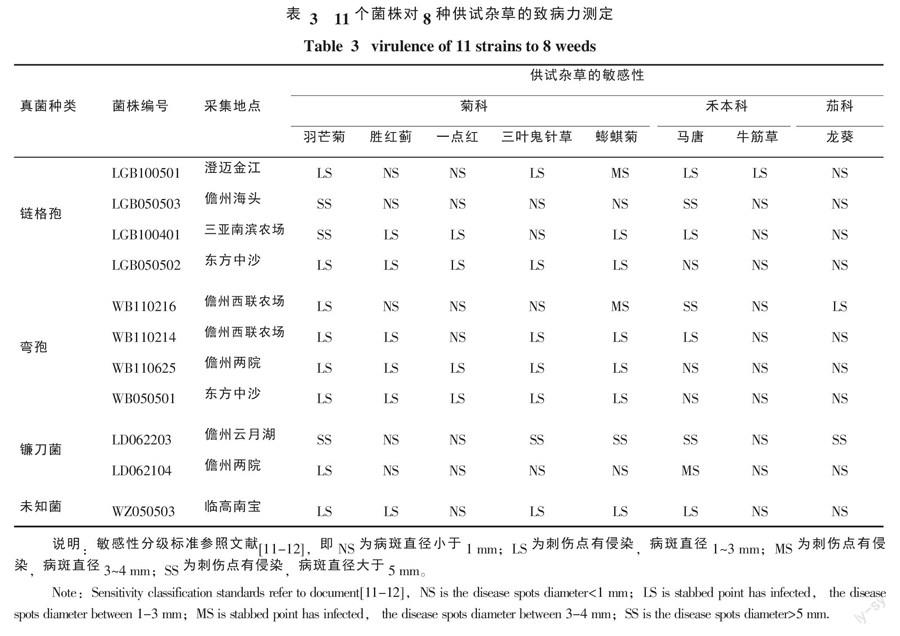
(3)寄主范圍測試。采用離體葉片針刺法測定11個菌株對常見菊科、禾本科及科雜草的致病力,以了解其寄主范圍,7 d后統計發病情況。其它方法見1.2.4(1)。
2 結果與分析
2.1 飛機草自然發病狀況及病原情況
海南的飛機草真菌性病害主要有5種,病原分別由鏈格孢屬、彎孢屬、炭疽屬、鐮刀菌屬、尾孢屬等引起,其中最為常見的是由尾孢屬引起的角斑病,在陰暗潮濕、生長密集的地方發病較為普遍,以老葉發病較為嚴重;由鐮刀菌屬引起的病害一般發生在葉緣處,病建交界比較明顯,病斑初期呈黑褐色,后期病葉逐漸變黃直至枯死,此病害只在儋州和海口兩地發現;由鏈格孢屬引起的病害,病斑呈明顯的輪紋狀,病斑初期為小黃點,后期多為不規則型深黑褐色病斑,葉緣處病斑較為明顯,此病害在東方、澄邁、三亞等地大量發生,其余地區零星分布;由彎孢屬引起的病害其病斑在葉片上隨機分布,與由鏈格孢侵染引起的病斑輪紋類似,但輪紋不明顯,后期病斑稍小,此病害只在儋州和澄邁兩地發現,發生較為嚴重;由炭疽菌屬引起的病害病斑為黃褐色的圓形及多角形,初期多從葉尖或葉緣發病,后期病斑連合成片,病部壞死變黑,此病在澄邁和儋州發生。
2.2 重要菌株的鑒定
2.2.1 鏈格孢(參考菌株LGB100401) 分生孢子梗一般比菌絲粗,顏色比菌絲略深;頂端產生手雷形或橢圓形的分生孢子(圖1-a),呈現暗灰褐色,多為單生,大多數具喙,橫隔1~5個,縱隔1~2個;孢子大小(31.88~56.79)μm×(10.92~13.61)μm。對其rDNA的ITS序列進行BLAST比對,從發病飛機草上分離的菌株LGB100401(登錄號KT209589)與GenBank中Alternaria alternata的相似性為100%。結合形態學與分子鑒定,確定菌株LGB100401為鏈格孢Alternaria alternata。
2.2.2 新月彎孢(參考菌株WB110216) 分生孢子梗單生或叢生,產于菌絲末端或中間細胞上,直立或彎曲,不分枝,有分隔,淡褐色至暗褐色,頂部產孢區顏色較淡。分生孢子(圖1-b)多為單生,深褐色,具隔膜3個,從基部起第3個細胞膨大且彎曲,顏色較深,孢子大小為(10.36~30.85)μm×(4.31~14.33)μm。對其rDNA ITS序列進行BLAST比對發現,從發病飛機草上分離的菌株WB110216 (登錄號KT209588)與GenBank中Curvularia lunata的相似性為99%。結合形態學與分子鑒定,確定菌株WB110216為新月彎孢Curvularia lunata。
2.2.3 尖孢鐮刀菌(參考菌株LD062203) 分生孢子梗(圖1-c),顏色較淺;產孢細胞單瓶梗(圖1-d),具分枝;大型分生孢子(圖1-e)多為單生,無色,3~5隔膜,足胞明顯,頂細胞逐漸變窄,形狀有鐮刀形、橢圓形等,大小(26.57~36.18)μm×(4.04~4.48)μm;小型分生孢子大小(12.67~15.52)μm×(3.67~4.40)μm,單生,無色,一般為0~1隔膜,形狀為腎形等;厚垣孢子大小為(7.80~12.41)μm×(7.82~13.48)μm, 多為簇生或頂生,球型,無橫隔。對其rDNA ITS序列進行BLAST比對發現,從發病飛機草上分離的菌株LD061903(登錄號KT339743)與GenBank中Fusarium oxysporum的同源性為99%。結合形態學與分子鑒定,確定菌株LD061903為尖孢鐮刀菌Fusarium oxysporum。
2.3 分離菌株致病力測定
2.3.1 菌絲對離體葉片的致病力初步測定 挑選分離率較高的5種病菌共46個菌株進行致病力測定。結果表明,強致病力菌株共有12株(占總26.09%),中等致病力菌株有13株(占28.26%), 弱致病力菌株有19株(占41.30%), 2個菌株無致病力(占4.3%)(表1)。
接種7 d后,供試的病原菌中,鏈格孢菌株LGB100501病情指數最高為84.00,其次是LGB100401菌株病情指數為79.44,發病癥狀見圖2;彎孢菌株WB110216病情指數最高為76.30;而鐮刀菌菌株LD062203(發病癥狀見圖2)和LD061903病情指數最高為56.00;炭疽菌株TJ110205病情指數最高為73.33;未知菌WZ050502、WZ050503菌株病情指數則高達96.75和96.89。
2.3.2 菌絲對飛機草活體的致病力測定 活體致病力測定結果表明,有4個菌株的發病率均達到100%,且病斑擴展較慢,接種7 d的病情指數較低,為30.00~55.00。其它7個菌株的發病率在50%~75%,病情指數在20.00~51.25。致病力達2級的僅有菌株LGB100501 和WB110216(表2)。
2.4 寄主范圍測試
11個菌株針對8種供試材料共88個測定中(表3),在不同植物中表現高度感病的菌株共有9個,分別為菌株LGB050503、LGB100401、LD062203(對羽芒菊);菌株LGB050503、WB110216、LD062203(對馬唐);菌株LD062203(對三葉鬼針草、蟛蜞菊、龍葵)。表現中度感病的菌株僅有3個,分別為菌株LGB100501、WB110216(對蟛蜞菊)和LD062104(對馬唐)。所有測定中,高度感病和中度感病的雜草很少,僅占13.6%;此外,少數為輕微致病(病斑不擴散),多數對供試雜草無致病性。
在供試病原菌中,鐮刀菌LD062203的侵染力最強,供試的8種雜草中有5種對其極為敏感,表現高度感病;其次是鏈格孢菌LGB050503、彎孢菌WB110216,2種植物對其極敏感或較敏感,表現中度感病;其余供試病原菌侵染力較弱,供試植物對其不敏感或稍敏感。對供試雜草而言,9個雜草表現為高度感病,其中菊科雜草羽芒菊和禾本科的馬唐各占3個,表現較為突出。
3 討論與結論
早期研究指出,飛機草尾孢C. eupatorii侵染速度遠滯后于飛機草的生長速率,其生防作用較小[13],因此,本研究主要針對飛機草鏈格孢、鐮刀菌、彎孢菌3種病原展開研究;通過離體和活體測定綜合分析,供試的46個菌株中以鏈格孢屬菌株LGB100501、LGB100401和彎孢屬菌株WB110216對飛機草致病力最強。寄主范圍測試發現,供試雜草對絕大部分菌株的敏感性較低,具有較高的選擇性;菊科雜草羽芒菊和禾本科雜草馬唐對部分菌株極敏感。總的來說,上述菌株對其原寄主飛機草的致病力強于對其他寄主植物的致病力。
活體測定與離體測定相比,大部分菌株的致病力都較弱,與自然條件下致病情況存在差異。可能的原因有:一是活體接種的保濕條件不夠;二是可能自然條件下大部分飛機草病原菌為復合侵染,而實驗室單一病原菌致病力相對較弱。Zachariades等[14]曾指出,從美洲分離的飛機草病原菌難以控制非洲南部飛機草,與該病原菌的致病力及野外的生存競爭能力有很大關系。田間雜草的生物防治是極其復雜的過程,受諸多因素的影響,因此,上述病菌對田間飛機草的生防價值還有待進一步確定。
國內,戴新賓等[15]在惡性入侵雜草紫莖澤蘭Eupatorium adenophora病株上分離獲得A. alternata,并將其菌絲體成功開發為真菌除草劑。另外,研究表明,鏈格孢A. alternata和彎孢C. lunata除了具有很強的致病能力外,它們產生的毒素也具有很好的除草活性,是開發新型微生物除草劑的重要材料來源,如AAC-毒素可防治紫莖澤蘭,其作為天然除草劑已經申請專利[16-17]。本研究篩選出的鏈格孢和新月彎孢菌株,表現出良好的生防潛力,其真菌毒素的除草活性研究及活性物質的分離工作正在進行。本研究結果豐富了飛機草的病菌資源和信息,為飛機草的生物防治奠定了基礎。
參考文獻
[1] Te Beets D C. The status of biological control of weeds with fungal pathogens[J]. Ann Rev Phytopathol, 1992(30): 637-658.
[2] Charudattan R. Biological control of weeds by means of plant pathogens: Significance for integrated weed management in modern agro-ecology[J]. BioControl, 2001, 46(2): 229-260.
[3] Trujillo E E. History and success of plant pathogens for biological control of introduced weeds in Hawaii[J]. Biological Control, 2004, 33(1): 113-122.
[4] Elango D E, Holden, Prior C. The potential of plant pathogens collected in Trinidad for biological control of Chromolaena odorata [J]. International Journal for Pest Management, 1993, 39(4): 393-396.
[5] Barreto R W, Evans, H C. The mycobiota of the weed Chromolaena odorata in southern Brazil, with particular reference to fungal pathogens for biological control[J]. Mycological Research, 1994, 98(10): 1 107-1 116.
[6] Alana den Breeyen, Johannes Z, Groenewald, et al. Morphological and molecular characterisation of Mycosphaerellaceae associated with the invasive weed[J]. Fungal Diversity, 2006, 23(2006): 89-110.
[7] Prashanthi S K, Kulkarni S. Aureobasidium pullulans, a potential mycoherbicide for biocontrol of eupatorium[J]. Current Science, 2005, 8(1): 18-21.
[8] 方中達. 植病研究方法(第三版)[M]. 北京: 中國農業出版社, 1998.
[9] 魏景超. 真菌鑒定手冊[M]. 北京: 科學出版社, 1982: 124-126.
[10] 全國明, 毛丹鵑, 章家恩, 等. 飛機草的繁殖能力與種子的萌發特性[J]. 生態環境學報, 2011, 20(1): 72-78.
[11] 萬佐璽, 強 勝, 徐尚成, 等. 鏈格孢菌的產毒培養條件及其毒素的致病范圍[J]. 中國生物防治, 2001(1): 10-15.
[12] 何 偉, 鄧 暉, 牛永春, 等. 東北地區四種常見禾本科雜草病原真菌資源調查及其致病性測定[J]. 植物保護, 2011(1): 99-104.
[13] Sangakkara U R, Attanayake K B, Dissanayake U, et al. Allelopathic impact of Chromolaena odorata[J]. Journal of Plant Diseases and Protection, 2008, 21: 323-326.
[14] Zachariades C, Strathie L W, Retief E, et al. Progress towards the biological control of Chromolaena odorata[J]. African Entomology, 2011, 19(2): 282-302.
[15] 戴新賓, 陳世國, 強 勝, 等. 鏈格孢菌毒素對紫莖澤蘭葉片光合作用的影響[J]. 植物病理學報, 2004, 34(1): 55-60.
[16] Qiang S, Zhu Y, Summerell B, et al. Mycelium of Alternaria alternata as a potential biological control agent for Eupatorium adenophorum[J]. Biocontrol Science and Technology, 2006, 16(7): 653-668.
[17] 姜述君, 強 勝, 朱云枝. 畫眉草彎孢霉菌除草活性化合物的分離鑒定及其生物活性測定[J]. 植物保護學報, 2006, 33(3): 313-318.
責任編輯:林海妹
