南草蜥和南滑蜥斷尾及再生的研究
2016-06-05 14:15:31葉志強劉文華葉靈娟倪洪斌徐大德
生態科學 2016年2期
葉志強, 劉文華, 葉靈娟, 倪洪斌, 徐大德
肇慶學院生命科學學院, 肇慶526062
南草蜥和南滑蜥斷尾及再生的研究
葉志強, 劉文華, 葉靈娟, 倪洪斌, 徐大德*
肇慶學院生命科學學院, 肇慶526062
葉志強, 劉文華, 葉靈娟, 等. 南草蜥和南滑蜥斷尾及再生的研究[J]. 生態科學, 2016, 35(2): 52-56.
YE Zhiqiang, LIU Wenhua, YE Lingjuan, et al. A case study on the tail loss and regeneration ofTakydromus sexlineatusandScincella reevesii[J]. Ecological Science, 2016, 35(2): 52-56.
研究肇慶地區南草蜥和南滑蜥斷尾和再生形態特征, 結果顯示, 斷尾位置、再生時間以及兩者的交互作用對南滑蜥和南草蜥的尾長均具有顯著影響。在自然斷尾下, 南滑蜥和南草蜥較多的斷尾位置位于近基部(斷尾1/2), 斷尾個體分別約占29.3%(87/297)和49.2%(29/59)。實驗表明, 二者的尾自切處在2天后開始愈合并長出再生尾, 南滑蜥在遠基部(全斷尾)的再生速度最高, 而南草蜥則在近基部(斷尾3/4), 其尾再生率都明顯高于南滑蜥。
南草蜥; 南滑蜥; 斷尾和再生; 尾自切; 再生率
1 前言
尾的自殘與再生是蜥蜴類動物的一種反捕行為,也是一種常見的現象; 是長期進化形成的保護性適應, 斷尾的蜥蜴在經過一段時間后, 斷尾處仍可再生出新尾[1~2], 蜥蜴自切所斷下的尾巴, 會自主地跳動和扭動, 從而轉移捕食者的注意力, 為自身逃跑爭取時間[3~6], 一種逃避敵害的自衛保護性行為[7]。迄今為止, 國內有關蜥蜴斷尾與再生的研究物種有3 科(石龍子科、蜥蜴科和壁虎科)15 種[8]: 即北草蜥(Takydromus septentrionalis)[1]、多疣壁虎(Gekko japonicus)[1]、快步麻蜥(Eremias velox)[2]、崇安地蜥(Platyplacopus sylvaticus)[9]、大壁虎(Gekko gecko)[10]等。
南滑蜥(Scincella reevesii)隸屬于爬行綱, 有鱗目, 蜥蜴亞目, 石龍子科, 滑蜥屬, 系卵胎生蜥蜴,主要分布于我國廣西、廣東等省, 一般棲息于平原和山區交界處的草叢中[11]。南草蜥(Takydomus sexlineatusDaudiin)隸屬于爬行綱, 蜥蜴目, 蜥蜴科, 草蜥屬,在我國主要分布于福建、湖南、貴州、云南、廣東、海南、廣西等省, 屬卵生蜥蜴, 柔性卵[11]。目前國內南滑蜥和南草蜥的斷尾與再生方面的研究尚未進行過相關的報道。
本文通過對南滑蜥和南草蜥的尾長、斷尾時間、斷尾長度以及再生時間等相關研究與數據分析, 主要探討兩種蜥蜴的斷尾程度和再生時間。分析二者不同的再生關系以及再生過程中的特點, 對兩種蜥蜴種內和種間斷尾情況、斷尾率形態學特點進行比較, 可為其研究兩種蜥蜴行為生態學和形態生物學特點以及為進一步探索爬行類生物行為生態學的研究提供參考資料。
2 材料與方法
2.1 實驗材料
本實驗所用的南滑蜥和南草蜥標本于2012年3月和 5月捕自肇慶市郊北嶺山, 并將其帶回生物園實驗室, 經測量、稱重、鑒定性別和剪趾標記后, 統計雌雄斷尾情況。南滑蜥和南草蜥按 1:1性比(20♀/20♂)關養在塑料箱(長×寬×高=530 mm× 370 mm×300 mm)內, 箱內底有沙礫、草皮等以模擬自然生境。南滑蜥和南草蜥能在箱內自由取食足量的面包蟲和飲水, 接受自然光照。定期在飲水中添加復合維生素、鈣片和礦物質, 確保動物獲得較全面的營養。室內溫度恒定在(25±1) ℃左右。待實驗完畢后,將所有的南草蜥和南滑蜥親本放回原捕地。
體溫測定用WMZ23型電子點溫計(上海醫療儀器廠), 體重稱取用日本生產(MOD-EL)型 DJ-200S電子天平(精度0.001mg), 形態測量用上海量具刃具廠0-200游標卡尺。
2.2 實驗設計
根據南滑蜥和南草蜥雌雄自然種群斷尾程度分為四種, 即尾基部(斷尾1/4)、近基部(斷尾1/2)、尾中部(斷尾 3/4)和遠基部(全斷尾), 定期觀察兩種蜥蜴的斷尾傷口愈合和新尾的再生情況。尾再生率的計算公式為: 尾再生率(d-1)=尾再生長度(cm)/再生時間(d)/平均全尾長(cm)。
2.3 數據處理
實驗數據用STATISITCA 6.0統計軟件包分析。數據進一步分析用Kolmogorov-Simirnov和Bartlett檢驗數據正態性和方差同質性。檢測后所有數據均滿足參數統計條件, 用單因素方差分析(one-way ANOVA)和重復檢驗方差分析(repeated-measure ANOVA)等統計方法對相關數據進行統計分析。描述性統計值用平均值 ± 標準誤表示, 顯著性水平設為α = 0.05。
3 結果與分析
3.1 野外斷尾情況及種間比較
本實驗所捕的426條(♂♂:♀♀ =151:275)南滑蜥中, 有297條(約70.0%, 105♂♂, 192♀♀)至少發生過一次尾自切(圖1), 野外自然種群斷尾率無明顯的性別差異(♂♂:♀♀ = 70.0% :70.0%;G= 0, df= 1,P>0.90)。所捕獲的動物中, 接近尾基部(斷尾 1/4)斷尾個體占24.6%(73/297), 近基部(斷尾1/2)斷尾個體約占 29.3%(87/297), 尾中部(斷尾 3/4)斷尾個體約占23.9%(71/297), 遠基部(全斷尾)斷尾個體約占22.2%(66/297)。不同部位斷尾發生頻率無兩性差異(G= 6.29, df= 3,P> 0.05)(圖2a)。合并兩性數據后發現, 尾基部、近基部和尾中部的斷尾頻率無顯著差異(G= 3.19, df= 3,P< 0.1)(圖2a)。
本實驗所捕的126條(♂♂:♀♀ =57:69)南草蜥中, 有59條(約46.8%, 32♂♂, 27♀♀)至少發生過一次尾自切(圖1), 野外自然種群斷尾率無明顯的性別差異(♂♂:♀♀ = 39.1%:56.1%;G= 3.64, df= 1,P> 0.05)。所捕獲的動物中, 接近尾基部(斷尾 1/4)斷尾個體越占 18.6%(11/59), 近基部(斷尾 1/2)斷尾個體約占49.2%(29/59), 尾中部(斷尾3/4)斷尾個體約占25.4% (15/59), 遠基部(全斷尾)斷尾個體約占6.8%(4/59)。不同部位斷尾發生頻率無性別差異(G= 1.83, df= 3,P> 0.1)(圖2b)。合并兩性數據后發現, 尾基部、近基部和尾中部的斷尾頻率存在顯著差異(G= 22.82, df= 3,P< 0.005), 大部分個體尾自切發生在接近尾基部處(圖2b)。
兩種蜥蜴野外斷尾發生頻率存在顯著的種間差異(SR:TS= 70.0%:46.8%;G= 21.55, df= 1,P<0.005)(圖 1), 不同部位斷尾發生頻率也存在顯著的種間差異(G= 13.78, df= 3,P< 0.005)(圖2)。
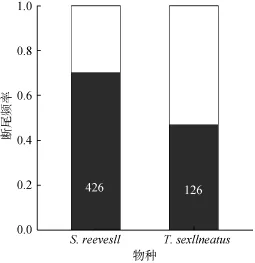
圖1 野外南滑蜥和南草蜥斷尾頻率(數據基于426條南滑蜥成體和126條南草蜥成體)Fig. 1 Tail loss rate of two lizards collected from the field. Data are based on 426 S.reevesii adults and 126T. sexlineatusadults.
3.2 尾再生及種間比較
表1顯示了斷尾再生實驗的南滑蜥和南草蜥體長及尾長的描述性統計值。
以斷尾位置和再生時間為因子對南滑蜥和南草蜥體長及尾長分別進行重復檢驗方差分析(repeatedmeasures ANOVA), 結果顯示斷尾位置、再生時間以及兩者的交互作用對南滑蜥和南草蜥的體長均無顯著影響(P> 0.09), 而三者對南滑蜥和南草蜥的尾長均具有顯著影響(P< 0.0001), 兩種蜥蜴尾長均隨斷尾程度的加深而變短, 尾再生14天后的尾長均明顯大于斷尾時的尾長。

表1 斷尾再生實驗的兩種蜥蜴體長和尾長的描述性統計值Tab. 1 Descriptive statistics for snout-vent length and tail length of two lizards in tail regeneration experiment.
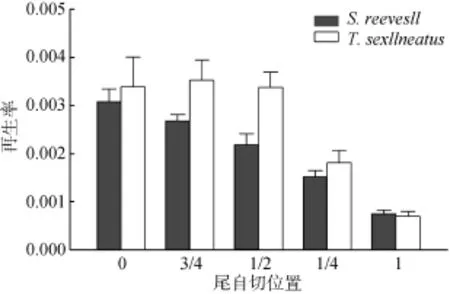
圖3 不同斷尾位置兩種蜥蜴尾再生率比較Fig. 3 Comparision of tail regeneration rate in two lizards at different position of tail loss
以物種和斷尾位置為因子對兩種蜥蜴的斷尾再生率進行雙因子方差分析(tow-way ANOVA), 結果表明斷尾位置對斷尾再生率具有顯著影響(F4, 58= 33.23,P< 0.0001), 嚴重斷尾的再生率明顯高于稍微斷尾和不斷尾; 而物種、物種和斷尾位置間交互作用不是斷尾再生率的變異來源(Species:F1, 58= 2.60,P= 0.112; interaction:F4, 58= 0.98,P= 0.424)(圖3)。
4 討論
從本實驗結果得知, 蜥蜴斷尾是有一定條件的,當蜥蜴尾部突然受到外部刺激過強時多數都會發生斷尾[8,12-13]。蜥蜴斷尾后, 在一段時間內都能長出再生尾, 只是再生尾的長度和生長時間因種類而出現差異[8]。然而蜥蜴的斷尾與再生過程基本上是一致的, 如多疣壁虎[1]、快步麻蜥[2]、崇安地蜥[8]、蛤蚧[14]和麗斑麻蜥[15]。
從圖1可知: 426條南滑蜥中, 至少有70.0%的蜥蜴發生過一次尾自切, 而 126條南草蜥中, 則至少有 46.8%, 這比麗斑麻蜥[15]和蝘蜓[16]的自然斷尾率都要高, 與壁虎屬(如無蹼壁虎[17])發生的自然斷尾率相近。所捕獲的南滑蜥中(圖2a), 斷尾位置較多位于近基部(斷尾 1/2), 其斷尾個體約占 29.3%, 而南草蜥中(圖 2b), 斷尾位置也是較多位于近基部(斷尾 1/2), 其斷尾個體約占 49.2%, 這與壁虎屬(如原尾蜥虎[18])的斷尾程度(常斷至尾基部)相一致。合并圖1和圖2數據顯示, 當蜥蜴斷去的尾長分別為完整尾的1 /4、1 /2和3 /4時, 其運動速度比具有完整尾部的個體要低[18]。在自然種群中南滑蜥和南草蜥尾遠基部斷尾的概率較低, 分別是 22.2%和 6.8%;這與藍尾石龍子[19]尾遠端斷尾的概率較低相符, 這可能與少量斷尾并不足以吸引天敵的注意力從軀干部分轉移到斷落尾段有關。此外, 較大程度的尾自切可能會顯著影響斷尾個體的適合度, 使其付出難以承受的生存代價。一般認為特定蜥蜴種類的斷尾程度應取決于尾自切的利益和代價之間的權衡[20]。蜥蜴尾部在運動過程中對平衡身體具有重要作用,尾自切導致尾部擺動頻率和后肢運動的變化最終影響運動速度[12,13,18], 所以當蜥蜴遇到敵害或受到外部強刺激時, 它們不惜斷去尾部來獲取更多的逃生機會。
實驗結果顯示斷尾位置、再生時間以及兩者的交互作用對南滑蜥和南草蜥的體長均無顯著影響,但三者對它們的尾長均具有顯著影響(表1)。兩種蜥蜴在春季斷尾時, 斷面上有少量血液滲出, 一般兩天后結出黑色的痂, 結痂的第三天就開始長出了黑色圓錐形的尾, 經過兩個月的生長(其間有蛻皮現象), 新老尾在粗細上已連續, 但顏色區別明顯。從表 1可知, 不管是哪種種類的哪種斷尾程度, 它們的體長在經過14天后無明顯變化, 但它們的再生尾在經過14天后明顯發生變化, 其中南滑蜥在遠基部(全斷尾)的再生速度最高(表1、圖3), 而南草蜥則是在近基部(斷尾3/4)(表1、圖3)。兩種蜥蜴的不同尾自切的再生速率表現大大不同, 南草蜥的尾部再生速率均大于南滑蜥(圖3)。斷尾再生速度可能受蜥蜴本身發育的影響, 可能是再生芽基中攜帶著尾的全部遺傳信息, 而生長受時間限制是一種有限度的生長。其次因為尾部是許多蜥蜴種類的重要能量貯存器官, 斷尾引起的尾部貯能丟失可減緩動物的生長速率[19]。而南草蜥的形態個體明顯大于南滑蜥, 因此南草蜥尾部貯存的能量多于南滑蜥, 導致南草蜥各尾部再生的速度都明顯高于南滑蜥(圖3)。統計結果表明: 斷尾位置對斷尾再生率具有顯著影響, 嚴重斷尾的再生率明顯高于稍微斷尾和不斷尾。
綜上所述, 本實驗在野外捕捉的兩種蜥蜴, 都出現不同程度尾自切現象, 并且南滑蜥出現尾自切的概率比南草蜥高, 其二者之間差異尚需更多的實驗數據進行進一步檢驗。從四種斷尾程度可知: 同種蜥蜴的斷尾程度越高, 蜥蜴的再生能力會相應提高(如3/4 > 1 /2 >1/4); 而種間蜥蜴: 南草蜥的3/4斷尾(1/2 和1/4斷尾)的再生速度均大于南滑蜥。本實驗結果, 南滑蜥和南草蜥斷尾和再生的比較是在斷尾位置、再生時間及其兩者的交互作用的基礎上發生的, 其再生率隨著不同的斷尾程度呈不同的生長速率。而對兩種蜥蜴斷尾及再生的調控機制、生理生化機理和影響因素以及代價等還有待于進一步研究。
[1] 趙肯堂, 張祥, 陳麗華. 北草蜥和多疣壁虎尾的自殘及再生研究[C]//趙爾宓. 從水生到陸生—劉承釗教授誕辰九十周年紀念文集:蛇蛙研究叢書(1). 北京:中國林業出版社, 1990: 138–144.
[2] 吾瑪爾. 快步麻蜥斷尾及其再生的實驗研究[J]. 干旱區研究, 2000, 17(2): 61–64.
[3] EDMUNDS M. Defence in animals: a survey of antipredator defences[M]. Longman Publishing Group, 1974.
[4] EVANS S E. Caudal autotomy in a lower Jurassic eosuchian[J].Copeia, 1981(4): 883–884.
[5] Arnold E N. Caudal autotomy as a defense[J]. Biology of Reptilia, 1985: 237–273.
[6] PAFILIS P, E D VALAKOS, J FOUFOPOULOS. Comparative postautotomy tail activity in six Mediterranean lacertid lizard species[J]. Physiological and Biochemical Zoology, 2005, 78(5): 828–838.
[7] 陳紹軍. 麗斑麻蜥的種群結構與自截斷尾再生的初步觀察[J]. 四川動物, 1994, 13(1): 173–174.
[8] 唐鑫生, 王潔平. 崇安地蜥斷尾再生與產卵行為初報[J].四川動物, 2006, 24(3): 366–369.
[9] 胡英超, 謝箭秧, 朱奇平, 等. 蜥蜴尾的自切及再生[J].生態學雜志, 2015, 34(5): 1441–1447.
[10] 陳振昆, 丁光. 大壁虎尾部及斷尾的解剖學研究[J]. 云南農業大學學報, 1996, 11(2): 68–71.
[11] 趙爾宓, 趙肯堂, 周開亞. 中國動物志: 爬行綱. 第二卷[M].北京: 中國科學院中國動物志編輯委員會, 科學出版社, 1999.
[12] 方俊九, 湯秀榮. 胎生蜥蜴在我國的分布, 形態及生活習性的觀察[J]. 兩棲爬行動物學報, 1983, 2(3): 51–55.
[13] 董丙君, 趙文閣, 劉志濤, 等. 胎生蜥蜴斷尾及再生的研究[J]. 四川動物, 2007, 26(2): 428–429.
[14] 林呂何. 蛤蚧的斷尾及其再生研究[J]. 動物學雜志, 1978, 2: 24–27.
[15] 陳紹軍. 麗斑麻蜥自截斷尾與蜥尾結構的關系[J]. 生物學通報, 1991, 26(2): 20–21.
[16] 李曉晨. 蝘蜓日活動節律、食性及繁殖特征的初步研究[J]. 動物學雜志, 1997, 32(6): 11–14.
[17] 陳紹軍. 無蹼壁虎的種群結構與自截斷尾的初步調查[J].中藥材, 1993, 3: 12–14.
[18] 徐大德, 計翔, 陸洪良, 等. 體溫, 攝食, 斷尾和雌體繁殖狀態對原尾蜥虎運動表現的影響[J]. 生態學報, 2009, 29(4): 1745–1755.
[19] 林植華, 樊曉麗, 陸洪良, 等. 斷尾對藍尾石龍子能量儲存和運動表現的影響[J]. 生態學報, 2010, 30(10): 2541–2548.
[20] MAGINNIS T L. The costs of autotomy and regeneration in animals: a review and framework for future research[J]. Behavioral Ecology, 2006, 17(5): 857–872.
A case study on the tail loss and regeneration of Takydromus sexlineatus and Scincella reevesii
YE Zhiqiang, LIU Wenhua, YE Lingjuan, NI Hongbin, XU Dade
Department of Biology,Zhaoqing University,Zhaoqing Guangdong526062,China
This experiment studys the tail loss and regeneration features ofTakydromus sexlineatusandScincella reevesiiin Zhaoqing. Result shows that the position of tail loss, regeneration time and their interactions have notable effects on tail length of two species. Most of the lossing tail position ofTakydromus sexlineatusand Scincella reevesiiis close to the near base(1/2 tail loss), and tail loss individuals accounts for 29.3% (87/297) and 49.2% (29/59) respectively in natural tail loss condition. The research shows that the tail loss part will get recovery and regenerate after 2 days. The highest regeneration rate is in the far base (all tail loss) for Takydromus sexlineatuswhile in the near base (3/4 tail loss) for Scincella reevesii. In short, the regeneration rate ofTakydromus sexlineatusis significantly higher thanScincella reevesii.
Takydromus sexlineatus, Scincella reevesii,tail loss and regeneration, tail loss, regeneration rate
10.14108/j.cnki.1008-8873.2016.02.008
Q 958.12
A
1008-8873(2016)02-052-05
2013-00-00;
2014-00-00
國家自然科學基金資助項目(31271124); 廣東省自然基金資助項目(s2013010016062); 大學生創新創業訓練計劃項目(DC201455)
葉志強(1957—), 實驗師, 研究方向為動物學, E-mail: ye57109@zqu.edu.cn
*通信作者:徐大德, E-mail: zqxdd@163.com
