多功能融合酶基因Linker的優(yōu)化及表達(dá)
2016-06-14 02:00:25王銳麗劉敏杰薛業(yè)敏
江蘇農(nóng)業(yè)科學(xué) 2016年4期
王銳麗+劉敏杰+薛業(yè)敏

摘要:在融合酶阿拉伯/木糖苷酶-木聚糖酶(Xar-xynA)之間插入不同組成和長度的多肽Linker,避免催化結(jié)構(gòu)域相互之間的干擾,改善融合酶的催化效率。5個(gè)含不同長度及Ala/Gly比例連接肽Linker的融合突變體在大腸桿菌重組表達(dá)、純化,并對融合酶的酶學(xué)性質(zhì)、動(dòng)力學(xué)參數(shù)和酶解作用進(jìn)行研究。結(jié)果表明,融合酶Xar-L1/L2/L3-xynA和融合酶Xar-xynA有著相似的最適溫度和最適pH值。其中融合酶Xar-L1-xynA在65~80 ℃和pH值6.6~8.2有更強(qiáng)的阿拉伯/木糖苷酶熱穩(wěn)定性和pH值穩(wěn)定性,將融合酶Xar-L1-xynA在65 ℃保溫60 min其阿拉伯/木糖苷酶活性有著明顯的激活現(xiàn)象。并獲得連接Xar和XynA的最優(yōu)Linker:L1和L4,麥麩和玉米芯木聚糖產(chǎn)物酶解試驗(yàn)的結(jié)果表明,融合酶Xar-L1-xynA和Xar-L4-xynA比其他的融合酶有著更高的催化效率。
關(guān)鍵詞:多功能性酶;基因融合;木糖苷酶/阿拉伯糖苷酶;木聚糖酶;連接肽
中圖分類號: Q786
文獻(xiàn)標(biāo)志碼: A
文章編號:1002-1302(2016)04-0031-05
木聚糖是存在于農(nóng)業(yè)廢棄物中最豐富的半纖維素,由于木聚糖結(jié)構(gòu)及降解方式的復(fù)雜性,徹底降解需要多種水解酶的協(xié)同作用。同時(shí)使用多種水解酶來降解木聚糖,雖然清潔高效,但工序復(fù)雜,成本高。近來Fan Z等[1-3]將有關(guān)水解酶融合形成多功能融合酶來作用于水溶性阿拉伯木聚糖,玉米芯等農(nóng)業(yè)廢棄物,這方面研究不但提高了利用酶的綜合效率,而且簡化了工序和降低了成本。
構(gòu)建的融合酶常因前后結(jié)構(gòu)域的相互干擾而影響各自活性的發(fā)揮,于是在2結(jié)構(gòu)域中插入多肽Linker(連接序列)來減弱或消除這種干擾。已發(fā)現(xiàn)有不少多肽可作為連接融合酶的2個(gè)結(jié)構(gòu)域的Linker,賦予多功能融合酶的空間柔性,增強(qiáng)酶位點(diǎn)識別,保持相臨結(jié)構(gòu)域的功能獨(dú)立(不受干擾)[4]。 Carlsson等在融合酶的2個(gè)催化單元間插入多肽Linker(Linker1:GDPLERSDP;Linker2:GDPLEANGIRSDP)使融合酶中的乳糖苷酶和半乳糖脫氫酶的活性提高[5];Lu等在融合酶中插入Linker (GGGGSGGGGS),使融合酶中的葡聚糖酶活性增加326%,木聚糖酶活性增加43%[6]等。
由于熱穩(wěn)定的酶在工業(yè)中有著廣泛的應(yīng)用前景,本研究選用疏棉狀嗜熱絲孢菌Thermomyces lanuginosus DSM 5826木聚糖酶A(XynA)作為重組融合酶的一個(gè)酶源。該木聚糖酶優(yōu)先降解高聚合度的木聚糖鏈,產(chǎn)物是不同聚合度的低聚木糖,不產(chǎn)生木單糖,這對于生產(chǎn)低聚木糖具有很好的應(yīng)用價(jià)值[7]。另外選擇1個(gè)具有雙重活性的酶即嗜熱厭氧乙醇菌Thermoanaerobacter ethanolicus產(chǎn)生的阿拉伯呋喃糖苷酶(Xar)作為融合酶的酶源[8],可以減少融合酶源的個(gè)數(shù)。
本研究針對半纖維素的結(jié)構(gòu)及其降解復(fù)雜的特點(diǎn),采用不同組分和長度的多肽Linker連接雙活性阿拉伯/木糖苷酶(Xar)和木聚糖酶A(XynA)構(gòu)建多功能半纖維素酶,并對融合酶的酶學(xué)性質(zhì)、動(dòng)力學(xué)參數(shù)和酶解作用進(jìn)行研究,以獲得三功能融合酶的催化效率增強(qiáng)的最優(yōu)linker。使得通過蛋白質(zhì)工程構(gòu)建多功能半纖維素酶來促進(jìn)生物轉(zhuǎn)化成為可能,為酶法降解半纖維素的工業(yè)化生產(chǎn)提供有效的技術(shù)路線和解決方案。
1 材料與方法
1.1 材料
質(zhì)粒和菌株:含有嗜熱乙醇菌JW200(Thermoanaerobacter ethanolicus JW200)α-阿拉伯糖苷酶/木糖苷酶(Xar)基因和疏棉狀嗜熱絲孢菌DSM 5826(Thermomyces lanuginosus DSM 5826)木聚糖酶A(XynA)基因的質(zhì)粒pET-20b-xar和pET-20b-xar-xynA由前期研究構(gòu)建保存。大腸桿菌(Escherichia coli)DH10B(Promega)用于質(zhì)粒擴(kuò)增和分子克隆等過程,E. coli JM109(DE3)(Promega)作為目的基因的表達(dá)宿主。
主要試劑:限制性內(nèi)切酶EcoRⅤ、XbaⅠ、XhoⅠ,Pyrobest聚合酶,T4 DNA連接酶購自TaKaRa公司;燕麥木聚糖(OSX),對硝基苯酚-α-阿拉伯呋喃糖苷(pNPAF)和對硝基苯酚-β-木糖苷(pNPX)購自Sigma公司。
1.2 方法
1.2.1 基因操作 DNA片斷的分離、DNA的內(nèi)切酶水解和連接、基因的電轉(zhuǎn)化等方法均參照文獻(xiàn)[9]。瓊脂糖電泳膠回收DNA片斷和質(zhì)粒的制備參考QIAGEN公司QIAquick Gel Extraction Kit和QIAprep Spin Plasmid Miniprep Kit試劑盒使用指南進(jìn)行。
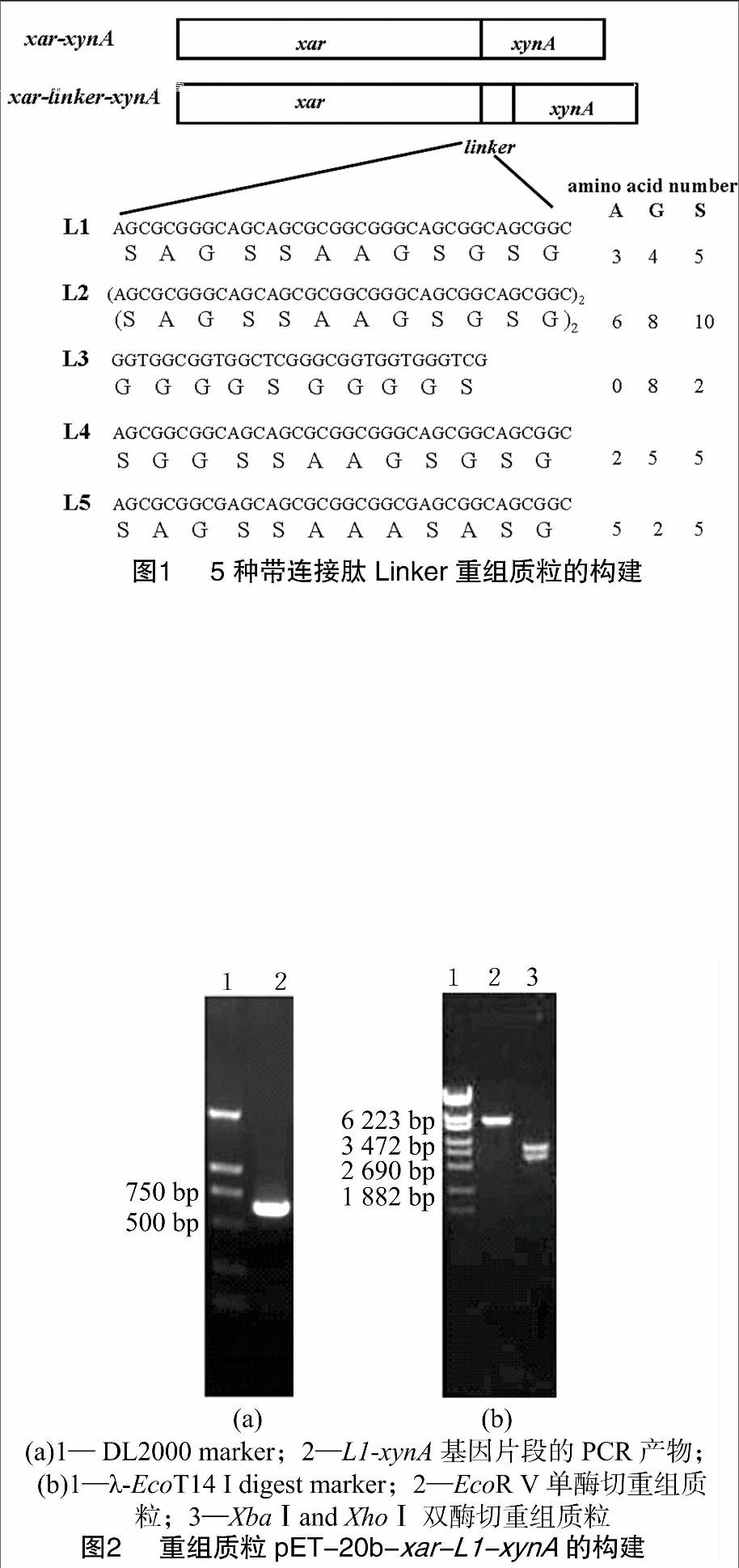
1.2.2 重組質(zhì)粒pET-20b-xar-linker-xynA的構(gòu)建 設(shè)計(jì)長引物將連接肽融入基因間,融合基因中插入的連接肽氨基酸序列及其名稱及本研究融合基因涉及的引物序列見表1。將12個(gè)氨基酸長度的連接肽L1(SAGSSAAGSGSG)相應(yīng)的堿基序列設(shè)計(jì)在長引物S1上,通過PCR將連接肽融入2個(gè)基因之間。以質(zhì)粒pET-20b-xar-xynA為模板,利用相應(yīng)的上下游引物S1和P4(表1)擴(kuò)增含連接肽L1的基因xynA。反應(yīng)體系(50 μL):10 × buffer 5 μL;dNTP(各2.5 mmol/L)4 μL;S1(50 μmol/L)1 μL;P4(50 μmol/L)1 μL;模板(10 μg/mL)1 μL;Probest DNA polymerase(1.25 U/μL) 1 μL;H2O 37 μL。PCR 擴(kuò)增參數(shù)均設(shè)為:先在95 ℃ 變性 5 min,加Pyrobest DNA聚合酶1 μL;然后94 ℃ 變性30 s,50 ℃ 退火40 s,72 ℃延伸1 min,循環(huán)30次后,72 ℃保溫 10 min。將所得PCR片段和載體用T4 DNA連接酶于16 ℃連接過夜,將連接液電轉(zhuǎn)化感受態(tài)細(xì)胞E. coli DH10B。挑取陽性克隆,提取質(zhì)粒,用單、雙酶切驗(yàn)證。送至上海美吉生物技術(shù)公司測序,正確的轉(zhuǎn)化子命名為pET-20b-xar-L1-xynA。
連接肽L2含有24個(gè)氨基酸,即(SAGSSAAGSGSG)2。其相應(yīng)的上下游引物為P1和S2(表1)。構(gòu)建成功的重組質(zhì)粒命名為 pET-20b-xar-L2-xynA。
連接肽L3的氨基酸序列為GGGGSGGGGS,同連接肽L1的插入方法一樣,參考大腸桿菌氨基酸偏好密碼子,將連接肽L3的堿基序列設(shè)計(jì)在長引物S3上。重組質(zhì)粒命名為pET-20b-xar-L3-xynA。
在連接肽L1的基礎(chǔ)上設(shè)計(jì)了連接肽L4:SGGSSAAGSGSG和L5:SAGSSAAASASG,在已構(gòu)建的重組質(zhì)粒的基礎(chǔ)上,設(shè)計(jì)了引物S4和S5。構(gòu)建的重組質(zhì)粒命名為pET-20b-xar-L4/L5-xynA。
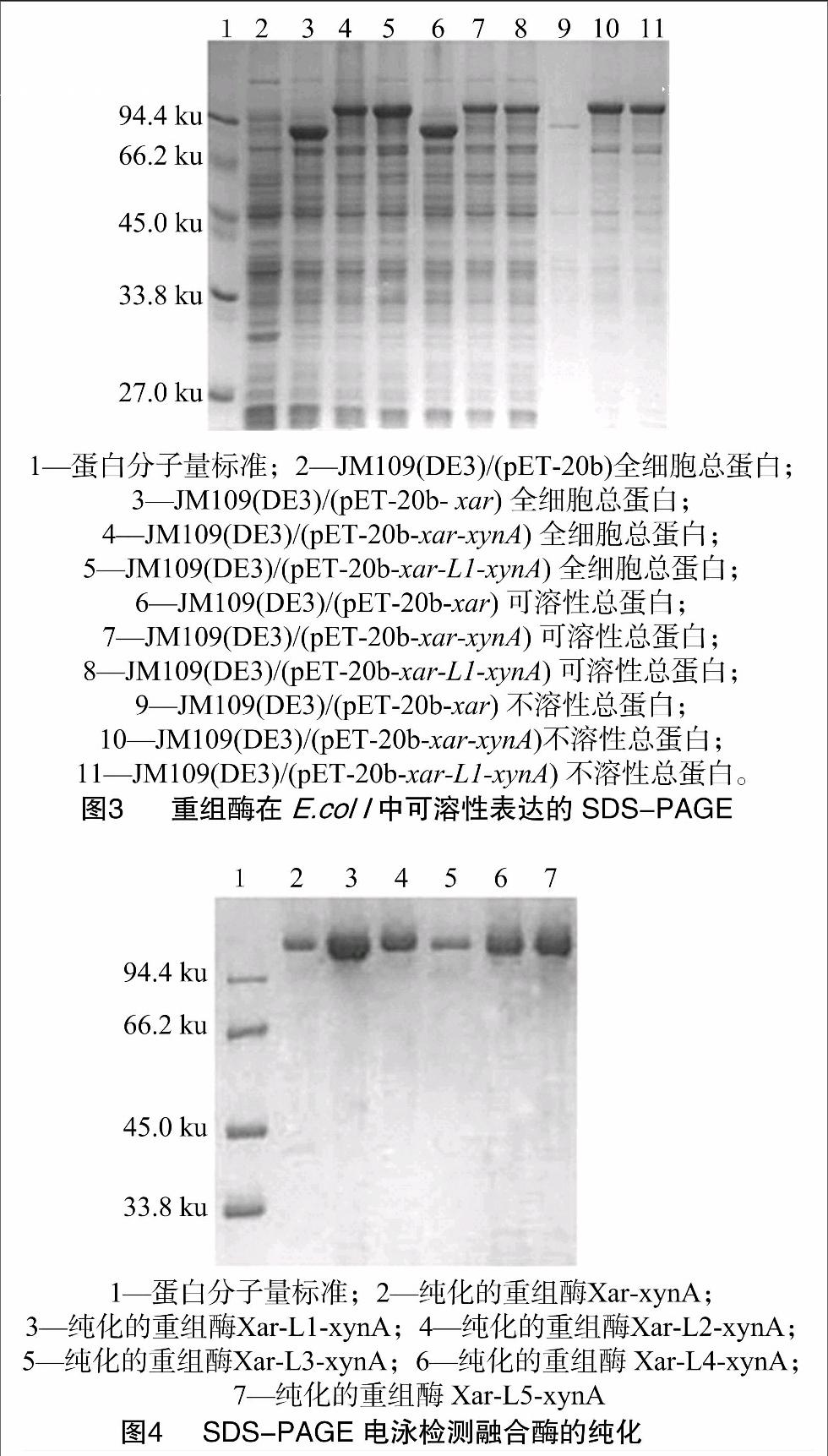
1.2.3 重組酶的可溶性表達(dá)和活性檢測 將重組質(zhì)粒pET-20b-xar,pET-20b-xar-xynA和pET-20b-xar-L1-xynA分別電轉(zhuǎn)化于E. coli JM109(DE3),挑取單菌落接入含Amp抗性的LB培養(yǎng)液,進(jìn)行IPTG誘導(dǎo)培養(yǎng)。待培養(yǎng)結(jié)束,檢測各樣品細(xì)胞密度,取菌液1 mL,離心收集菌體。用 100 mmol/L pH值6.2的鄰苯二甲酸氫鉀-咪唑緩沖液 (PIB) 重懸,調(diào)整細(xì)胞密度一致。經(jīng)超聲破碎,離心30 min,上清即為粗酶液。分別進(jìn)行木聚糖酶、阿拉伯糖苷酶和木糖苷酶活性的測定[8,10]。同時(shí)進(jìn)行SDS-PAGE,以檢測各種重組質(zhì)粒在宿主E. coli JM109(DE3)的可溶性表達(dá)情況。
1.2.4 重組酶的純化 將質(zhì)粒pET-20b-xar-xynA和pET-20b-xar-L1/L2/L3/L4/L5-xynA分別電轉(zhuǎn)化于JM109(DE3),挑取單菌落接種于LB培養(yǎng)基中。將培養(yǎng)好的菌液離心收集細(xì)胞,用高壓破碎法破碎細(xì)胞,離心,接著65 ℃下熱處理20 min,最后用Ni2+親和層析法純化帶連接肽的重組融合酶。
1.2.5 重組酶Xar-L1/L2/L3-xynA的酶學(xué)性質(zhì)
1.2.5.1 最適反應(yīng)溫度的測定 木聚糖酶、木糖苷酶、阿拉伯糖苷酶活性的最適反應(yīng)溫度的測定:酶促反應(yīng)在50 mmol/L pH值 6.2的PIB緩沖液中進(jìn)行。在45~95 ℃范圍,每隔5 ℃ 分別測定3種酶的活力。以最高酶活力為100%,計(jì)算相對活性。
1.2.5.2 最適反應(yīng)pH值 的測定 木聚糖酶、木糖苷酶和阿拉伯糖苷酶活性分別在65 ℃、90 ℃和80 ℃下測定不同的pH值(4.2~8.2)對酶活力的影響,以最高酶活為100%,計(jì)算相對活性。
1.2.5.3 熱穩(wěn)定性的測定 木聚糖酶活性熱穩(wěn)定性測定,是將融合酶分別置于50~80 ℃范圍內(nèi),每隔5 ℃下保溫1 h,然后在65 ℃、pH值 6.2測定殘留活力,以保存于冰浴中的酶活力為100%,計(jì)算相對活性,找到酶活力半衰期為1 h的溫度。木糖苷酶活性熱穩(wěn)定性的測定,是將酶置于65~90 ℃下保溫1 h,然后在90 ℃、pH值6.2測定殘留活力。阿拉伯糖苷酶活性熱穩(wěn)定性的測定,是將酶置于65~90 ℃下保溫1 h,然后在80 ℃、pH值 6.6 測定殘留活力。
1.2.5.4 pH值穩(wěn)定性的測定 將純酶置于不同pH值(4.2~8.2)的100 mmol/L PIB緩沖液中,在37 ℃下保溫 1 h,冷卻,然后補(bǔ)加相應(yīng)的底物和緩沖液達(dá)到200 μL。再于最適溫度和最適pH值下分別測定殘留活力性,以保存于冰浴中的酶活力為100%,作pH值穩(wěn)定性曲線。
1.2.5.5 動(dòng)力學(xué)參數(shù)的測定 用pH值6.2 PIB緩沖液分別配制0.25~2.5 mg/mL的燕麥木聚糖(OSX)和0.02~0.2 mmol/L 的pNPX作為底物,用pH值5.8的緩沖液配制0.1~1.0 mmol/L的pNPAF作為底物,然后分別在最適溫度65 ℃、90 ℃、70 ℃測重組融合酶的木聚糖酶、木糖苷酶和阿拉伯糖苷酶的活性。采用Lineweaver-Burk作圖法,計(jì)算重組融合酶3種活性的Km、Vmax及Kcat/Km值,其中Kcat=最大反應(yīng)速度/蛋白摩爾數(shù)。
1.2.6 融合酶的酶解試驗(yàn) 分別稱取玉米芯木聚糖(CX)[11]和去淀粉麥麩(DSWB)[12]200 mg溶于pH值 7.0、250 mmol/L的磷酸緩沖液中,總體系為5 mL。以不加入任何融合酶的酶解體系為試驗(yàn)對照,其他的試驗(yàn)項(xiàng)(1~6)分別加入純化融合酶Xar-xynA和Xar-L1/L2/L3/L4/L5-xynA。以加入的融合酶Xar-L1-xynA的木聚糖酶酶單位(20 U)為標(biāo)準(zhǔn),其他試驗(yàn)項(xiàng)加入等摩爾數(shù)相對應(yīng)的融合酶。于65 ℃條件下反應(yīng)1、4、10、24 h后取樣利用DNS法測還原糖濃度。
2 結(jié)果與分析
2.1 重組質(zhì)粒pET-20b-xar-linker-xynA的構(gòu)建
重組質(zhì)粒pET-20b-xar-xynA和pET-20b-xar-L1/L2/L3/L4/L5-xynA構(gòu)建的示意圖見圖1。
將連接肽L1的堿基序列設(shè)計(jì)在長引物S1上,通過PCR將連接肽融入2個(gè)基因之間。擴(kuò)增的DNA片段與實(shí)際大小一致為600 bp,沒有明顯的非特異性條帶,說明所選PCR擴(kuò)增條件能有效擴(kuò)增L1-xynA基因片段(圖2-a)。
對于已構(gòu)建的重組質(zhì)粒pET-20b-xar-L1-xynA采用XbaⅠ和XhoⅠ雙酶切,得到線性的pET-20b質(zhì)粒和插入的xar-L1-xynA基因片段,大小分別為3.7 kb和2.9 kb,與預(yù)計(jì)結(jié)果的相同(圖2-b),測序結(jié)果表明該質(zhì)粒構(gòu)建成功。
用同樣的方法構(gòu)建和驗(yàn)證其他4種帶連接肽的重組質(zhì)粒pET-20b-xar-L2/L3/L4/L5-xynA。
2.2 融合酶Xar-L1-xynA的可溶性表達(dá)
