外源ABA處理的玉米葉片酵母雙雜交cDNA文庫(kù)的構(gòu)建及評(píng)價(jià)
2016-06-14 01:16:54馬芳芳蔣明義
江蘇農(nóng)業(yè)科學(xué) 2016年4期
馬芳芳+蔣明義

摘要:為深入研究ABA信號(hào)轉(zhuǎn)導(dǎo)機(jī)制以及篩選該信號(hào)途徑中相關(guān)重要蛋白的相互作用,利用Gateway技術(shù),以外源ABA處理的玉米葉片為材料首先構(gòu)建了cDNA初級(jí)文庫(kù),初級(jí)文庫(kù)再通過(guò)LR重組的方式最終構(gòu)建成以pDEST22為目的載體的酵母雙雜交cDNA文庫(kù)。經(jīng)質(zhì)量鑒定,該文庫(kù)的滴度為3.9×106 CFU/mL,文庫(kù)總?cè)萘窟_(dá)到1.17×107 CFU,平均插入cDNA片段長(zhǎng)度大于800 bp,重組率大于95%。結(jié)果表明所獲酵母雙雜交文庫(kù)質(zhì)量較高,符合文庫(kù)構(gòu)建的標(biāo)準(zhǔn),保證了文庫(kù)的覆蓋度,為進(jìn)一步開(kāi)展互作蛋白篩選的后續(xù)工作奠定了基礎(chǔ)。
關(guān)鍵詞:ABA;玉米;酵母雙雜交;cDNA文庫(kù);滴度;重組率;互作蛋白
中圖分類號(hào): S513.01;Q78
文獻(xiàn)標(biāo)志碼: A
文章編號(hào):1002-1302(2016)04-0058-04
脫落酸(ABA)是一種重要的植物激素,它在植物生長(zhǎng)發(fā)育的各個(gè)方面,包括種子發(fā)育、萌發(fā)和休眠、營(yíng)養(yǎng)生長(zhǎng)及對(duì)各種生物、非生物脅迫的響應(yīng)等方面起作用[1-2]。水分脅迫下(包括干旱、鹽漬),植物體通過(guò)迅速積累ABA以促進(jìn)氣孔關(guān)閉以及調(diào)節(jié)眾多基因的表達(dá),從而使植株對(duì)逆境環(huán)境做出適應(yīng)反應(yīng)[3-6]。大量研究表明,ABA信號(hào)轉(zhuǎn)導(dǎo)是一個(gè)非常復(fù)雜的網(wǎng)絡(luò),許多重要的中間組分如胞內(nèi)鈣離子、鈣調(diào)素、磷酸肌醇、cADPR、H+、活性氧、一氧化氮以及蛋白可逆磷酸化等都在ABA信號(hào)轉(zhuǎn)導(dǎo)途徑中發(fā)揮著重要的作用[7]。然而ABA參與的許多信號(hào)途徑的詳細(xì)過(guò)程特別是具體的分子作用機(jī)理仍有待闡明,鑒定參與ABA信號(hào)途徑的細(xì)胞組分,并對(duì)其功能進(jìn)行闡明分析,將有助于我們提高對(duì)ABA信號(hào)轉(zhuǎn)導(dǎo)途徑的認(rèn)識(shí)。
酵母雙雜交系統(tǒng)是一種用來(lái)研究蛋白質(zhì)間相互作用的簡(jiǎn)便、快速而有效的方法[8],該方法由Fields和Song于1989年首先建立,至今被廣泛應(yīng)用于生物研究的各個(gè)領(lǐng)域[9-10]。它的一個(gè)很重要的功能就是發(fā)現(xiàn)新的蛋白質(zhì)以及發(fā)現(xiàn)蛋白質(zhì)的新功能,即從cDNA文庫(kù)中篩選出與已知蛋白相互作用的未知蛋白,進(jìn)而為探索該蛋白的相關(guān)特性及功能奠定基礎(chǔ)[11-15]。所以,高質(zhì)量酵母雙雜交 cDNA 文庫(kù)的構(gòu)建是利用酵母雙雜交技術(shù)進(jìn)行大規(guī)模互作蛋白篩選的前提和保障[16]。近年來(lái),針對(duì)許多重要農(nóng)作物如水稻[17]、小麥[18]、大麥[19]、玉米[20]等,眾多學(xué)者構(gòu)建了以不同組織、器官、細(xì)胞類型或分化時(shí)期為材料的酵母雙雜交cDNA文庫(kù),為相關(guān)作物的蛋白質(zhì)組學(xué)研究提供了基礎(chǔ)。然而國(guó)內(nèi)外關(guān)于外源ABA處理的玉米cDNA文庫(kù)的相關(guān)研究還未見(jiàn)報(bào)道,本研究以施用外源ABA的玉米為材料,利用Gateway技術(shù)構(gòu)建了ABA處理的玉米葉片酵母雙雜交cDNA文庫(kù),旨在有效地分離與已知蛋白相互作用的靶蛋白,以期為深入研究ABA信號(hào)網(wǎng)絡(luò)提供一定的理論依據(jù)。
1 材料與方法
1.1 供試材料
1.1.1 植物材料 以玉米(Zea mays L.)雜交種農(nóng)大108為材料,將玉米種子浸泡12 h后,25 ℃催芽,挑取發(fā)芽一致的種子播于含有營(yíng)養(yǎng)液的沙土中培養(yǎng)[晝/夜溫度28 ℃/22 ℃,光照度200 μmol/(m2·s),相對(duì)濕度75%,晝/夜光周期14 h/10 h]。當(dāng)幼苗的第2片真葉完全展開(kāi)時(shí),用鋒利的刀片將玉米幼苗從莖基部快速切割下來(lái),放在純水中2 h以消除傷害脅迫[21],然后用100 μmol/L外源ABA進(jìn)行處理[溫度25 ℃、光強(qiáng)200 μmol/(m2·s)],在不同的處理時(shí)間點(diǎn)(0.25、0.5、1、2、4、8、12、24 h),分別剪取第2片葉,迅速置于液氮中冷凍,將最終所得材料混合均勻,-80 ℃ 保存?zhèn)溆谩?/p>
1.1.2 菌株和載體 大腸桿菌DH10B、文庫(kù)載體pDONR222(用于構(gòu)建初級(jí)文庫(kù))、pDEST22(用于構(gòu)建酵母雙雜交文庫(kù))均購(gòu)自Invitrogen公司。
1.1.3 主要試劑 Trizol Reagent、CloneMiner cDNA Library Construction Kit、FastTrack 2.0 mRNA Isolation Kit、1 kb Plus DNA Ladder、UltraPureTMPhenol-Chloroform-Isoamyl Alcohol (體積比25 ∶24 ∶1) 購(gòu)自Invitrogen公司;氨芐青霉素、卡那霉素為Sigma公司產(chǎn)品;Taq DNA Polymerase、DNA Marker Ⅲ、瓊脂糖凝膠DNA回收試劑盒、質(zhì)粒小提試劑盒購(gòu)自天根生化科技有限公司。
1.1.4 引物 M13 Forward,5′-GTAAAACGACGGCCAG-3′; M13 Reverse,5′-CAGGAAACAGCTATGAC-3′; pDEST22 Forward,5′-TATAACGCGTTTGGAATCACT-3′; pDEST22 Reverse,5′-AGCCGACAACCTTGATTGGAGAC-3′。所用引物由上海生工生物技術(shù)有限公司合成。
1.2 玉米葉片酵母雙雜交cDNA文庫(kù)的構(gòu)建
1.2.1 玉米葉片總RNA的提取和mRNA的純化 取玉米葉片,在預(yù)冷的研缽中將其用液氮研磨成細(xì)粉,用Trizol提取其總RNA,具體過(guò)程參照說(shuō)明書進(jìn)行。得到的總RNA經(jīng)過(guò)質(zhì)量及純度檢測(cè)之后,用于下一步的mRNA純化操作,具體的純化過(guò)程參照FastTrack2.0 Kit說(shuō)明書進(jìn)行。得到的mRNA經(jīng)質(zhì)量及純度檢測(cè)之后,用于下一步的文庫(kù)構(gòu)建操作。
1.2.2 cDNA初級(jí)文庫(kù)的構(gòu)建 參照CloneMiner cDNA Library Construction Kit說(shuō)明書,按以下步驟進(jìn)行:首先將上步純化得到的mRNA進(jìn)行cDNA第1鏈的合成;然后進(jìn)行cDNA第2鏈的合成及純化;cDNA與三框重組接頭attB1 Adapter連接后進(jìn)行cDNA的分級(jí)分離及收集;利用Gateway技術(shù)將收集得到的cDNA與入門文庫(kù)載體質(zhì)粒pDONR222混合進(jìn)行BP重組反應(yīng);重組反應(yīng)產(chǎn)物經(jīng)Proteinase K處理以及純化之后,利用電轉(zhuǎn)化法轉(zhuǎn)入大腸桿菌DH10B;加入1 mL SOC培養(yǎng)基,37 ℃培養(yǎng)1 h,即為cDNA初級(jí)文庫(kù)菌液;加甘油至終濃度20%,-80 ℃保存?zhèn)溆谩?
1.2.3 酵母雙雜交cDNA文庫(kù)的構(gòu)建 參照CloneMiner cDNA Library Construction Kit說(shuō)明書,將上述得到的cDNA初級(jí)文庫(kù)菌液接種于含有卡那霉素抗性的肉湯培養(yǎng)基中,30 ℃過(guò)夜培養(yǎng);第2天中抽質(zhì)粒并檢測(cè)吸光度;將中抽得到的質(zhì)粒稀釋到終濃度300 ng/μL,取出其中1 μL,利用Gateway技術(shù)將其與酵母雙雜交文庫(kù)載體質(zhì)粒pDEST22(含GAL4 Transcriptional-Activation Domain)混合進(jìn)行LR重組反應(yīng);重組反應(yīng)產(chǎn)物經(jīng)Proteinase K處理以及純化之后,利用電轉(zhuǎn)化法轉(zhuǎn)入大腸桿菌DH10B;加入3 mL SOC培養(yǎng)基,37 ℃培養(yǎng)1 h,即為酵母雙雜交cDNA文庫(kù)菌液;加甘油至終濃度20%,-80 ℃保存?zhèn)溆谩?/p>
1.2.4 文庫(kù)質(zhì)量的鑒定
1.2.4.1 庫(kù)容量的鑒定 分別從“1.2.2”節(jié)和“1.2.3”節(jié)得到的轉(zhuǎn)化后文庫(kù)細(xì)菌原液中取出10 μL,稀釋1 000倍后,從中吸出50 μL涂布于含有相應(yīng)抗性的LB平板上(初級(jí)文庫(kù)鑒定平板使用卡那霉素抗性、酵母雙雜交文庫(kù)鑒定平板使用氨芐青霉素抗性),37 ℃培養(yǎng)12~16 h后進(jìn)行計(jì)數(shù),根據(jù)公式:文庫(kù)滴度(CFU/mL)=平板克隆數(shù)×稀釋倍數(shù)/涂板體積;文庫(kù)總?cè)萘浚–FU)=文庫(kù)滴度×文庫(kù)總體積,計(jì)算出文庫(kù)的滴度以及總?cè)萘俊?/p>
1.2.4.2 重組率和插入片段長(zhǎng)度鑒定 從平板上隨機(jī)挑取24個(gè)單克隆進(jìn)行菌落PCR,初級(jí)文庫(kù)菌落PCR使用的引物Primer 1和引物Primer 2分別是M13 Forward和M13 Reverse,酵母雙雜交文庫(kù)菌落PCR使用的引物Primer 1和引物Primer 2分別是pDEST22 Forward和pDEST22 Reverse,PCR反應(yīng)條件:94 ℃預(yù)變性5 min;94 ℃變性30 s,56 ℃復(fù)性30 s,72 ℃延伸3 min,共30個(gè)循環(huán);72 ℃延伸10 min;4 ℃保溫。PCR產(chǎn)物用1% Agarose 凝膠電泳檢測(cè),鑒定插入片段大小和陽(yáng)性克隆重組率。
2 結(jié)果與分析
2.1 玉米葉片總RNA和純化的mRNA質(zhì)量分析
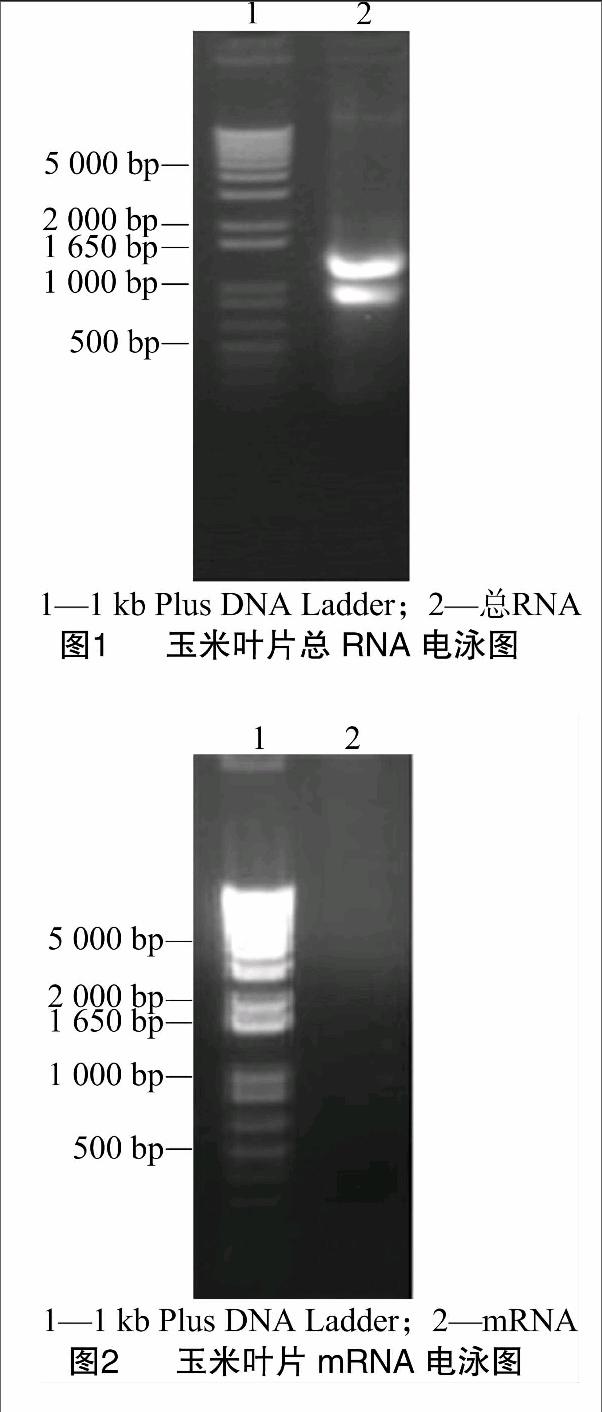
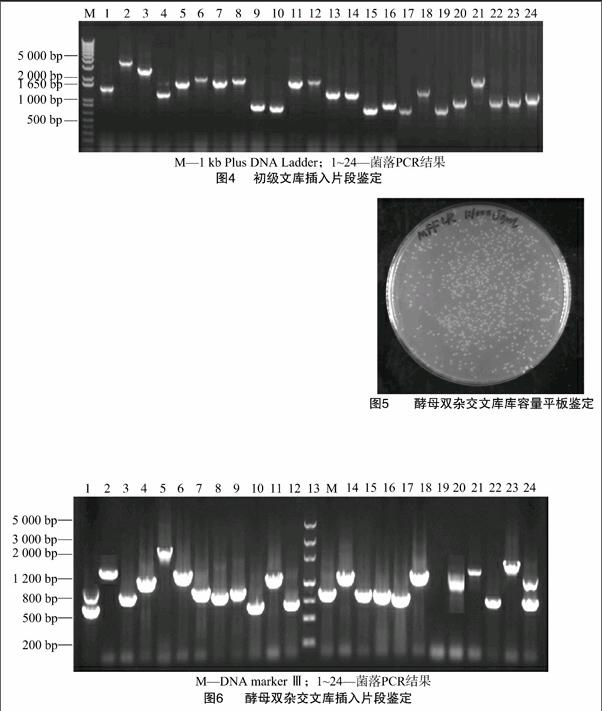
利用Trizol Reagent提取了外源ABA處理的玉米葉片總RNA,用1%瓊脂糖凝膠電泳檢測(cè),可見(jiàn)28S和18S條帶明亮清晰,亮度比約2 ∶1,表明該總RNA完整性好且基本上未發(fā)生降解(圖1)。經(jīng)總RNA紫外分析測(cè)定,結(jié)果顯示D260 nm/D280 nm為1.95,表明總RNA純度好,符合實(shí)驗(yàn)要求,可用于后續(xù)實(shí)驗(yàn)操作。總RNA樣品經(jīng)過(guò)分離純化后得到了mRNA,電泳圖結(jié)果可見(jiàn)其在較大范圍內(nèi)呈涂布狀分布(圖2),與預(yù)期結(jié)果相符,說(shuō)明純度較高,完整性較好。
2.2 cDNA初級(jí)文庫(kù)的質(zhì)量鑒定
2.2.1 庫(kù)容量測(cè)定 從cDNA初級(jí)文庫(kù)原始菌液中取出10 μL,稀釋1 000倍后,吸出50 μL涂布于含卡那霉素抗性的LB平板上,37 ℃培養(yǎng)12~16 h后計(jì)菌落數(shù),結(jié)果為643個(gè)(圖3),菌液量為1 mL。根據(jù)公式:文庫(kù)滴度(CFU/mL)=平板克隆數(shù)×稀釋倍數(shù)/涂板體積;文庫(kù)總?cè)萘浚–FU)=文庫(kù)滴度×菌液總體積,計(jì)算得出:cDNA初級(jí)文庫(kù)的滴度=643×1 000/0.05=1.28×107 CFU/mL;cDNA初級(jí)文庫(kù)的總?cè)萘?1.28×107 CFU/mL×1 mL=1.28×107 CFU。
2.2.2 重組率和插入片段大小測(cè)定 從平板上隨機(jī)挑取24個(gè)單克隆進(jìn)行菌落PCR,PCR產(chǎn)物用1% Agarose 凝膠電泳檢測(cè),根據(jù)電泳圖,可以看出所有克隆的PCR結(jié)果都呈陽(yáng)性,重組率達(dá)100%,插入cDNA片段長(zhǎng)度大小不等,主要分布在500~3 000 bp范圍內(nèi),多態(tài)性較好,且平均插入片段長(zhǎng)度大于1 000 bp(圖4)。
2.3 酵母雙雜交文庫(kù)cDNA文庫(kù)的質(zhì)量鑒定
2.3.1 庫(kù)容量測(cè)定 從酵母雙雜交cDNA文庫(kù)原始菌液中取出10 μL,稀釋1 000倍后,吸出50 μL涂布于含氨芐青霉素抗性的LB平板上,37 ℃培養(yǎng)12~16 h后計(jì)菌落數(shù),結(jié)果為195個(gè)(圖5),菌液量為3 mL。根據(jù)公式:文庫(kù)滴度(CFU/mL)=平板克隆數(shù)×稀釋倍數(shù)/涂板體積;文庫(kù)總?cè)萘浚–FU)=文庫(kù)滴度×菌液總體積,計(jì)算得出:酵母雙雜交文庫(kù)的滴度=195×1 000/0.05=3.9×106 CFU/mL;酵母雙雜交文庫(kù)的總?cè)萘?3.9×106 CFU/mL×3 mL=1.17×107 CFU。
2.3.2 重組率和插入片段大小測(cè)定 從平板上隨機(jī)挑取24個(gè)單克隆進(jìn)行菌落PCR,PCR產(chǎn)物用1% Agarose 凝膠電泳檢測(cè),從電泳圖中可以看出24個(gè)克隆中有23個(gè)為陽(yáng)性結(jié)果,重組率大于95%,cDNA插入片段長(zhǎng)度大小不等,主要分布在500~3 000 bp范圍內(nèi),多態(tài)性較好,且平均插入片段長(zhǎng)度大于800 bp(圖6)。
3 討論
一個(gè)高質(zhì)量酵母雙雜交 cDNA 文庫(kù)的獲得,能夠?yàn)槲覀冞M(jìn)行大規(guī)模互作蛋白的篩選提供前提和保障,對(duì)于我們研究相關(guān)生物的功能蛋白質(zhì)組學(xué)具有重要意義。目前很多生物種類的cDNA文庫(kù)已經(jīng)獲得,并且在此基礎(chǔ)上獲得了大量的信息[17-20],然而關(guān)于外源ABA處理的玉米cDNA文庫(kù)的相關(guān)研究還未見(jiàn)報(bào)道。已有的研究發(fā)現(xiàn),一些蛋白激酶如MAPK[22]、CCaMK[23-25]、CDPK[26]等在ABA誘導(dǎo)的抗氧化防護(hù)過(guò)程中發(fā)揮著重要的作用,我們擬利用酵母雙雜交技術(shù)篩選與這些蛋白相互作用的靶蛋白,進(jìn)而探究該蛋白在ABA信號(hào)途徑中的具體作用機(jī)制,因此,外源ABA處理的玉米葉片酵母雙雜交cDNA文庫(kù)的建立就成為了不可或缺的重要基礎(chǔ)工具。
RNA質(zhì)量的好壞直接影響文庫(kù)質(zhì)量的高低,本研究中提取的玉米葉片總RNA純度高、完整性好且基本上未發(fā)生降解,經(jīng)分離純化后得到的mRNA在較大范圍內(nèi)呈涂布狀分布,具有較高的純度和完整性,可用于后續(xù)的文庫(kù)構(gòu)建。本研究在構(gòu)建酵母雙雜交cDNA文庫(kù)的過(guò)程中使用了Gateway技術(shù),一種基于已研究的非常清楚的λ噬菌體位點(diǎn)特異性重組系統(tǒng),該系統(tǒng)能夠高效而快速地將DNA定向重組進(jìn)入不同的載體系統(tǒng)中,進(jìn)而應(yīng)用于蛋白的表達(dá)與功能分析[27-29]。由于在載體構(gòu)建時(shí)無(wú)需經(jīng)歷限制性內(nèi)切酶的酶切和酶連等過(guò)程,并且通過(guò)引入3種不同的接頭使得插入片段能夠包含3種不同的讀碼方式,這樣既降低了嵌合克隆出現(xiàn)的概率、保證了插入片段的序列完整性,同時(shí)又克服了由于插入片段中非編碼區(qū)帶有終止密碼子而造成的蛋白提前終止翻譯,有效地提高了文庫(kù)的完整性和覆蓋性[30]。
通常一個(gè)高質(zhì)量的cDNA文庫(kù),其庫(kù)容量應(yīng)至少大于 1×106 CFU,這樣才能保證cDNA種類的完整性以及文庫(kù)的代表性。本試驗(yàn)對(duì)構(gòu)建的玉米葉片酵母雙雜交cDNA文庫(kù)的分析結(jié)果表明,庫(kù)容量為1.17×107 CFU,保證了文庫(kù)的覆蓋度。而平均插入cDNA片段長(zhǎng)度則體現(xiàn)了文庫(kù)遺傳信息的完整性,如果大部分克隆插入了全長(zhǎng)cDNA序列,則說(shuō)明文庫(kù)的完整性好。本試驗(yàn)構(gòu)建的文庫(kù)重組率大于95%,平均插入cDNA 片段長(zhǎng)度大于800 bp,說(shuō)明該文庫(kù)達(dá)到了高質(zhì)量文庫(kù)的標(biāo)準(zhǔn),可以用來(lái)進(jìn)行后續(xù)的大規(guī)模互作蛋白的篩選試驗(yàn)。
參考文獻(xiàn):
[1]Zhu J K. Salt and drought stress signal transduction in plants [J]. Annu Rev Plant Biol,2002,53:247-273.
[2]Xu Z Y,Kim D H,Hwang I. ABA homeostasis and signaling involving multiple subcellular compartments and multiple receptors [J]. Plant Cell Rep,2013,32:807-813.
[3]Cutler S R,Rodriguez P L,F(xiàn)inkelstein R R,et al. Abscisic acid:emergence of a core signaling network [J]. Annu Rev Plant Biol,2010,61:651-679.
[4]Fujita Y,F(xiàn)ujita M,Shinozaki K,et al. ABA-mediated transcriptional regulation in response to osmotic stress in plants [J]. J Plant Res,2011,124:509-525.
[5]Hubbard K E,Nishimura N,Hitomi K,et al. Early abscisic acid signal transduction mechanisms:newly discovered components and newly emerging questions [J]. Genes Dev,2010,24:1695-1708.
[6]Umezawa T,Nakashima K,Miyakawa T,et al. Molecular basis of the core regulatory network in ABA responses:sensing,signaling and transport [J]. Plant Cell Physiol,2010,51:1821-1839.
[7]Wasilewska A,Vlad F,Sirichandra C,et al. An update on abscisic acid signaling in plants and more [J]. Mol Plant,2008,1:198-217.
[8]張雨良,張樹(shù)珍,王健華,等. 感染高粱花葉病毒甘蔗葉片cDNA文庫(kù)構(gòu)建及評(píng)價(jià) [J]. 熱帶作物學(xué)報(bào),2012,33(6):1096-1100.
[9]Fields S,Song O. A novel genetic system to detect protein-protein interactions [J]. Nature,1989,340:245-246.
[10]Song O,Dolan J W,Yuan Y L,et al. Pheromone-dependent phosphorylation of the yeast STE12 protein correlates with transcriptional activation [J]. Genes Dev,1991,5:741-750.
[11]Walhout A J,Boulton S J,Vidal M. Yeast two-hybrid systems and protein interaction mapping projects for yeast and worm [J]. Yeast,2000,17:88-94.
[12]Ito T,Ota K,Kubota H,et al. Roles for the two-hybrid system in exploration of the yeast protein interactome [J]. Mol Cell Proteomics,2002,1:561-566.
[13]Auerbach D,Thaminy S,Hottiger M O,et al. The post-genomic era of interactive proteomics:facts and perspectives [J]. Proteomics,2002,2:611-623.
[14]Chen Y,Xu D. Computational analyses of high-throughput protein-protein interaction data [J]. Curr Protein Pept Sci,2003,4:159-181.
[15]Cho S,Park S G,Lee D H,et al. Protein-protein interaction networks:from interactions to networks [J]. J Biochem Mol Biol,2004,37:45-52.
