擬南芥缺失突變體at14a的比較轉錄組分析
2016-06-14 01:29:15王琳孫慶玲劉輝梁建生
江蘇農業科學 2016年4期
王琳+孫慶玲+劉輝+梁建生


摘要:以at14a相關基因型(野生、缺失)擬南芥植株為試驗材料,利用轉錄組學和生物信息學等方法,分析野生型(col)、缺失突變體(at14a)擬南芥差異基因表達狀況,初步探討擬南芥AT14A的功能。結果表明:at14a在擬南芥植株根、莖、葉和花中均有表達,尤其在葉片中的表達量較高;相較于野生型,缺失突變體at14a中有194個差異基因,其中上調基因122個,下調基因72個;通過GO富集分析發現,2種基因型植株差異基因主要參與了應答脅迫過程,這表明at14a可能通過調控AT5G39610、AT1G53240、AT4G32770、AT1G02920、AT1G10760這些逆境脅迫相關基因的表達來參與應答脅迫。差異分子主要定位于細胞核,具有綁定的分子功能。通過pathway分析發現,2個基因型植株差異基因主要參與了核糖體代謝、碳水化合物代謝、光合等代謝途徑。研究結果為深入研究at14a作用的分子機理奠定了基礎。
關鍵詞:AT14A;擬南芥;缺失突變體;基因芯片;轉錄組學
中圖分類號:S188;Q786
文獻標志碼: A
文章編號:1002-1302(2016)04-0070-04
植物細胞壁可能與膜蛋白相互作用,其中一類膜蛋白就是與動物細胞整合素蛋白相似的類整合素蛋白。整合素是動物細胞黏附分子的重要成員,屬于一類跨膜蛋白的超家族,是由2個跨膜糖蛋白亞基(α、β)通過非共價鍵連接而成的異二聚體,它們黏附于細胞外基質(ECM)并結合細胞骨架。整合素分子的結構特點使其成為動物細胞內外信號雙向轉導過程中的關鍵分子之一,它能介導細胞膜雙向信號轉導,即通過調控細胞外的信號從而調控其與細胞外配體的結合活性;同時,細胞外基質與整合素的結合誘導細胞內的信號傳遞過程。整合素在多種生理活動中具有重要的功能,它們調節細胞的遷移、極性、生長、分化,并與基因表達的信號轉導元件相互作用[1-3]。在植物中,整合素尚未被證實,有可能是因為植物已經將這些蛋白功能以不同于哺乳動物細胞的方式進行組合,以適應植物質膜的獨特結構,并與細胞壁相互作用[4]。自從Schindler等首次發現與動物整合素具有相似功能的類整合素蛋白以來,在植物中已經確定了許多類整合素蛋白[5-6]。
Nagpal等在1999年報道了在擬南芥cDNA庫中用抗脊椎動物整合素β1亞基胞內高度保守結構域抗體作探針分離和鑒定出1個與整合素具有部分相似序列的cDNA克隆——AT14A,這為植物類整合素的存在提供了直接證據。AT14A(At3G28300)編碼基因有1個1 154 bp的開放閱讀框,只存在1個外顯子,由1 459個核苷酸組成,其中10至1 164位核苷酸組成編碼385個氨基酸的蛋白,分子量預測為43 ku。序列分析發現,AT14A蛋白有1個小的結構域與真菌、昆蟲和人類的整合素存在序列相似性,并有1個編碼11個氨基酸的高度保守區,它與人類D1整合素胞內結構域同源性較高[7]。筆者前期研究表明,擬南芥AT14A蛋白功能類似于動物中的整合素,介導了懸浮細胞細胞壁-質膜-細胞骨架連續體(WMC)的連接[8]。然而,關于AT14A發揮生理功能的機制尚不清楚。
轉錄組學是新興的研究細胞表型和功能的重要手段,在研究基因結構、表達和功能上開拓了新型研究方向。轉錄組(transcriptome)概念最先是由Velculescu等在1997年提出的,是指某一特定生物體在特定狀態下所有基因轉錄產物的總和,主要包括mRNA、非編碼RNA[9]。相對于基因組而言,轉錄組更具有時間性、空間性,轉錄組反映的是特定條件下活躍表達的基因[10],基因芯片技術是轉錄組學分析的常用方法。它起源于20世紀90年代,是指高密度固定在硅片、玻片、陶瓷等固相支持介質上的生物分子微陣列,具有快速、高效、微型化、高通量、大規模、高度并行性等特點[11]。因此,本研究通過基因芯片技術分析AT14A對擬南芥基因表達的影響,從而探討AT14A的功能,以期為類整合素的研究提供理論依據。
1 材料與方法
1.1 試驗材料
本試驗所用的植物材料是擬南芥(Arabidopsis thaliana)。at14a基因位于3號染色體上,編號為AT3G28300。以野生型(Col)、純合突變體(at14a)(SALK_101761)2種基因型擬南芥為材料。野生型擬南芥(Col)由加拿大大不列顛哥倫比亞大學陳金桂博士提供,后經筆者所在實驗室自行繁種保存。at14a是AT14A的T-DNA插入功能缺失突變體。以不同組織的擬南芥為材料,提取總RNA,用于at14a基因的表達分析。以生長6周的擬南芥葉片為材料,提取總RNA,用于基因芯片分析。
1.2 試劑
總RNA提取試劑盒:RNAprep Pure Plant Kit[天根生化科技(北京)有限公司];cDNA第1鏈合成試劑盒(Roche);RT-qPCR Mix(Bio-Rad);RNA提取和反轉錄過程中所用EP管及槍頭均為無菌級(美國Axygen公司);熒光定量PCR試劑盒:SYBR Green Supermix(美國BIO-RAD公司);Taq DNA 聚合酶:Thermo Scientific DreamTaq Green DNA聚合酶。引物由上海捷瑞生物工程公司合成。
1.3 熒光定量PCR方法
根據植物總RNA提取試劑盒的說明,提取擬南芥葉片總RNA,并參照反轉錄試劑盒的說明,以總RNA為模板,反轉錄為cDNA,用于at14a的表達分析。以擬南芥數據庫中的at14a基因編碼序列為參照,用Primer 5軟件設計at14a基因定量PCR引物。以根、莖、葉、花等組織的cDNA 為模板,擴增擬南芥at14a基因,進行表達量分析。反應體系20 μL,其中上下游引物各0.5 μL,定量PCR混合液10 μL,模板cDNA 1 μL,補雙蒸水至20 μL。反應條件:95 ℃ 1 min;95 ℃15 s,60 ℃1 min,40個循環;95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s,60 ℃ 30 s,導出數據使用ABI 7300自帶軟件進行定量分析。
1.4 基因芯片分析
1.4.1 RNA樣品制備及芯片雜交 采用Trizol法提取擬南芥葉片中的總RNA,通過異丙醇沉淀法濃縮RNA,并進一步采用RNeasy Mini Spin Column試劑盒對總RNA進行柱純化,用分光光度計定量、甲醛變性膠電泳質檢。采用RNA擴增標記方法合成帶生物素標記的cRNA,將純化后的cRNA與Agilent 公司擬南芥全基因組芯片雜交過夜,然后在芯片工作站進行芯片洗脫,采用芯片掃描儀進行掃描。這些工作均由北京博奧生物有限公司完成,每個樣品重復3次。
1.4.2 芯片圖像的采集與數據分析 采用Feature Extraction圖像分析軟件對芯片圖像進行分析,將圖像信號轉化為數字信號。然后對信號值進行歸一化處理,用SAM(Significance Analysis of Microarrays)軟件進行差異基因的分析。FDR控制在5%以內,再以倍數變化大于2倍或小于50%的標準篩選差異基因。
1.5 生物信息學分析
本試驗通過TAIR Gene Ontology (GO) Annotations tool (http://www.arabidopsis.org/tools/bulk/go/index.jsp )和MAS(Molecular Annotation System)系統對差異基因進行分析。
1.6 統計分析
研究中每個值的表達方式為“平均值±標準誤差”,不同處理之間進行了多重比較,所有數據進行了單因素方差分析。
2 結果與分析
2.1 at14a的組織表達特性
圖1表明,在擬南芥根、莖、葉和花各組織中,at14a都有表達,尤其是葉片中的at14a基因表達量最高,根、莖中的at14a表達量最低,花中at14a表達量居于兩者之間。
2.2 總RNA質檢和芯片雜交質量檢測
采用Trizol法提取6個擬南芥樣本葉片的總RNA,純化后,紫外分光光度計測定D230 nm、D260 nm、D280 nm,計算出各RNA樣品的濃度(表1)。由圖2可知,RNA樣品電泳條帶清晰。總體看出,RNA樣品的純度、總量及完整性符合Agilent表達譜芯片試驗要求,可以繼續后續芯片試驗。
6個樣本純化后cRNA產物分別與芯片進行雜交和掃描,芯片雜交掃描結果顯示擬南芥基因組中所有基因表達的熒光原始信號強度。由圖3可知,芯片四周邊界點線均勻一致,表明芯片質量可靠。信號檢測報告表明:樣品各項檢測參數達到質控要求,雜交控制探針等陽性對照信號、看家基因信號值正常;平均背景值與噪音值較低;陽性率數值正常。這些結果說明,本組基因芯片的質量、雜交和檢測體系均無問題,芯片檢測結果可靠。
2.3 不同基因型擬南芥轉錄組分析
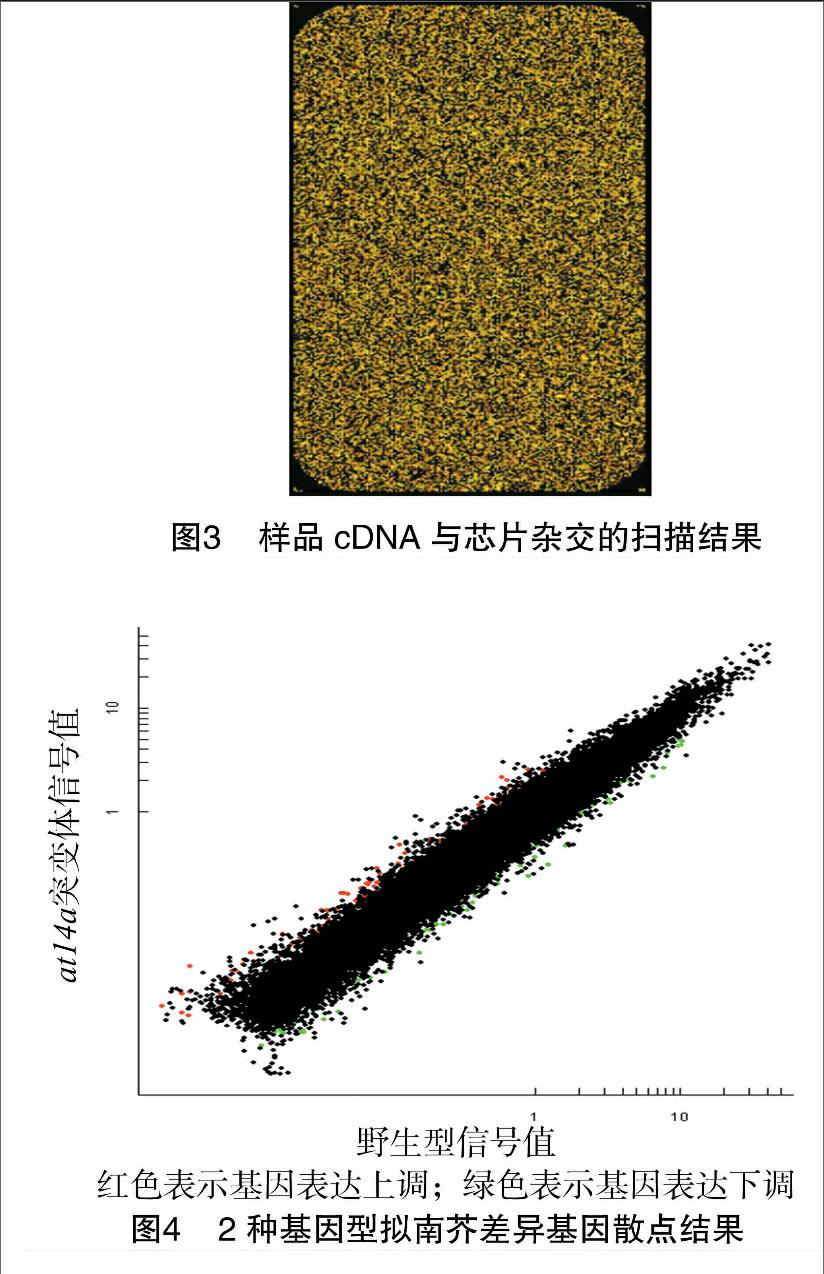
通過軟件對野生型(col)和缺失突變體(at14a)植株葉片的差異基因進行了分析。由圖4可知,野生型與缺失突變體植株的差異基因共194個,其中122個上調,72個下調。
2.3.1 不同基因型擬南芥差異基因的基因本體(Gene Ontology)分析 Gene Ontology數據庫旨在建立注釋基因和蛋白質知識的標準詞匯體系。GO功能注釋包括細胞定位(cellularcomponent)、分子功能(molecular function)、生物學過程(biological processes),3者緊密聯系,共同描述基因的特征。細胞定位是指基因產物在細胞中的位置;分子功能描述基因或基因產物的分子生物學活性和功能;生物學過程通常由多種分子功能有序組成。通過Gene Ontology分析,有助于更好地了解擬南芥at14a缺失后,植株基因表達的變化規律。由圖5-A可知,2個基因型植株葉片差異基因參與的生物過程多種多樣,如細胞代謝、應答脅迫、運輸、信號轉導、轉錄等,應答脅迫是其中比較重要的過程。這從轉錄水平上說明,定位于質膜的at14a作為細胞壁-質膜-細胞骨架連續體的中間分子,可能在擬南芥響應脅迫過程中起關鍵作用。由圖5-B可知,2個基因型植株葉片差異基因主要定位于細胞核,分布比例高達20.35%,其次是細胞質、細胞內、葉綠體等部位。這為at14a定位于質膜,并參與細胞壁的組成提供了組織基礎。由圖5-C可知,2個基因型植株葉片差異基因主要具有綁定功能,還有轉移酶活性、激酶活性、水解酶活性等功能。這表明at14a具有和其他基因相互作用的分子基礎。
2.3.2 不同基因型擬南芥差異基因的Pathway分析 日本京都基因和基因組百科全書(KEGG)是生物系統學的數據庫,它整合了細胞功能、生物體特性知識,描述了分子間相互作用。由圖6可知,2種基因型植株葉片差異基因參與的代謝途徑多種多樣,如核糖體代謝、淀粉與蔗糖代謝、氨基糖代謝、與細胞色素P450相關的異源性代謝、葉綠素卟啉環代謝、乙醛酸代謝、色氨酸代謝、光合代謝等。這些結果表明,at14a可能參與這些代謝途徑的調控。
3 討論與結論
葉片在植物光合作用、蒸騰作用、水和營養物質的吸收等過程中發揮重要作用,從而影響整個植物的生長和發育。因此,葉片組織的轉錄組學研究在植物轉錄組學的研究中具有舉足輕重的作用。at14a在葉片中表達較高,這為它在生物進程中發揮功能提供了組織基礎。
近期的研究表明,AT14A是WMC連續體中必不可少的核心分子,可以作為植物細胞壁和細胞骨架之間的連接載體,與動物整合素一樣,在控制極性和形態中起重要作用。它參與了細胞質膜與細胞壁間的黏附,并參與了滲透脅迫誘導的質-壁分離過程[8]。越來越多的證據表明,在植物細胞中WMC連續體對脅迫起重要的調控作用,但是它通過膜蛋白介導的分子機制還不清楚。相關研究表明,植物類整合素參與了霉菌毒素的滲透作用、機械刺激應答、細胞壁黏附等[12]。筆者的研究也發現,2個基因型植株差異基因主要參與了應答脅迫過程。
AT5G39610(ATNAC2)是編碼NAC結構域的轉錄因子。
在鹽脅迫下,野生型、NTHK1轉基因株系中該基因表達上調;此外,它還參與氧化脅迫[13]。AT4G32770(VTE1)編碼生育酚環化酶,與生育酚(維生素E)的合成有關,它參與應答高光、氧化、低溫等逆境脅迫過程[14]。AT1G53240(MMDH1)編碼線粒體蘋果酸脫氫酶,它參與冷脅迫和鹽脅迫應答過程,并參與細菌防御反應;它還參與蘋果酸、三羧酸循環等碳水化合物的代謝過程[15]。AT1G02920(GST11)編碼谷胱甘肽轉移酶,具有和鈷離子、銅離子、谷胱甘肽結合的功能,它定位于細胞質、細胞核和液泡等部位,參與應答鹽脅迫、細菌和真菌防御,還參與谷胱甘肽代謝和毒素分解代謝等途徑[16]。AT1G10760(SEX1)編碼淀粉降解所需的α-葡聚糖水合二激酶,它具有α-葡聚糖水合二激酶活性,能與ATP、金屬離子結合,它定位于葉綠體、線粒體等部位,參與了應答冷脅迫過程,與淀粉分解途徑有關[17]。AT14A可能通過調控這些逆境相關脅迫基因的表達,從而參與了擬南芥響應逆境脅迫過程。
本試驗通過轉錄組學和生物信息學的方法初步分析了擬南芥缺失突變體at14a的基因表達變化及其參與的生物學過程和代謝途徑等,分析結果較直觀,為進一步研究at14a作用的分子機制奠定了基礎。
參考文獻:
[1]Hynes R O. Integrins:bidirectional,allosteric signaling machines[J]. Cell,2002,110(6):673-687.
[2]Qin J,Vinogradova O,Plow E F. Integrin bidirectional signaling:a molecular view[J]. PLoS Biology,2004,2(6):726-729.
[3]Wegener K L,Partridge A W,Han J,et al. Structural basis of integrin activation by talin[J]. Cell,2007,128(1):171-182.
[4]Kumar M N,Hsieh Y F,Verslues P E. At14a-Like1 participates in membrane-associated mechanisms promoting growth during drought in Arabidopsis thaliana[J]. Proceedings of the National Academy of Sciences of the United States of America,2015,112(33):10545-10550.
[5]Nick P. Microtubules,signalling and abiotic stress[J]. Plant Journal for Cell & Molecular Biology,2013,75(2):309-323.
[6]Schindler M,Meiners S,Cheresh D A. RGD-dependent linkage between plant cell wall and plasma membrane:consequences for growth[J]. Journal of Cell Biology,1989,108(5):1955-1965.
[7]Nagpal P,Quatrano R S. Isolation and characterization of a cDNA clone from Arabidopsis thaliana with partial sequence similarity to integrins[J]. Gene,1999,230(1):33-40.
[8]Lü B,Liang J S,Zhang J H,et al. AT14A mediates the cell wall-plasma membrane-cytoskeleton continuum in Arabidopsis thaliana cells[J]. Journal of Experimental Botany,2012,63(11):4061-4069.
[9]Velculescu V E,Zhang L,Zhou W,et al. Characterization of the yeast transcriptome[J]. Cell,1997,88(2):243-251.
[10]章 瓊,蔣高中,李 冰. 水產動物對氨氮脅迫響應的轉錄組分析研究進展[J]. 江蘇農業科學,2015,43(3):227-230.
[11]魏松紅,劉志恒,紀明山,等. 基因芯片技術在植物病害中的應用[J]. 河南農業科學,2008(3):20-22.
[12]Knepper C,Savory E A,Day B. Arabidopsis NDR1 is an integrin-like protein with a role in fluid loss and plasma membrane-cell wall adhesion[J]. Plant Physiology,2011,156(1):286-300.
[13]He X J,Mu R L,Cao W H,et al. AtNAC2,a transcription factor downstream of ethylene and auxin signaling pathways,is involved in salt stress response and lateral root development[J]. The Plant Journal,2005,44(6):903-916.
[14]Havaux M,Eymery F,Porfirova S,et al. Vitamin E protects against photoinhibition and photooxidative stress in Arabidopsis thaliana[J].The Plant Cell,2005,17(12):3451-3469.
[15]Nakaminami K,Matsui A,Nakagami H A,et al. Analysis of differential expression patterns of mRNA and protein during cold-acclimation and de-acclimation in Arabidopsis[J]. Molecular & Cellular Proteomics,2014,13(12):3602-3611.
[16]Kouno T,Ezaki B. Multiple regulation of Arabidopsis AtGST11 gene expression by four transcription factors under abiotic stresses[J]. Physiologia Plantarum,2013,148(1):97-104.
[17]Yano R,Nakamura M,Yoneyama T,et al. Starch-related α-glucan/water dikinase is involved in the cold-induced development of freezing tolerance in Arabidopsis[J]. Plant Physiology,2005,138(2):837-846.張彥妮,李兆婷,張艷波,等. 毛百合試管鱗莖形成和膨大的培養優化[J]. 江蘇農業科學,2016,44(4):74-78.
