篦子三尖杉牻牛兒基牻牛兒基焦磷酸合成酶基因的克隆與序列分析
2016-06-14 01:34:47齊小瓊胡晨熙李大衛(wèi)
江蘇農(nóng)業(yè)科學(xué) 2016年4期
關(guān)鍵詞:植物
齊小瓊+胡晨熙+李大衛(wèi)

摘要:三尖杉是我國(guó)特有植物,主要含有生物堿、黃酮及萜類等生物活性物質(zhì)。以篦子三尖杉為試驗(yàn)材料,根據(jù)同源克隆的方法擴(kuò)增三尖杉牻牛兒基牻牛兒基焦磷酸合酶(GGPPS)基因,并通過(guò)生物信息的方法對(duì)該基因進(jìn)行鑒定和分析。結(jié)果表明,篦子三尖杉GGPPS基因全長(zhǎng)1 217 bp,含有1個(gè)完整的1 182 bp開放閱讀框(ORF),編碼393個(gè)氨基酸的蛋白序列,GGPPS蛋白相對(duì)分子量為43.042 ku,理論等電點(diǎn)(pI)為6.57,其二級(jí)結(jié)構(gòu)主要由α-螺旋和無(wú)規(guī)則卷曲組成;蛋白質(zhì)同源分析表明,三尖杉GGPPS含有多聚異戊二烯基合成酶家族中保守的5個(gè)特征性結(jié)構(gòu)和2個(gè)富含天冬氨酸的區(qū)域;同源模建分析顯示,三尖杉GGPPS具有GGPPS蛋白典型的三維結(jié)構(gòu),特別與薄荷GGPPS蛋白的三維結(jié)構(gòu)及活性位點(diǎn)極其相似;系統(tǒng)進(jìn)化分析表明,三尖杉GGPPS蛋白歸屬植物進(jìn)化支,且與加拿大紅豆杉、曼地亞紅豆杉、海南粗榧的GGPPS蛋白歸為同一分支。
關(guān)鍵詞:篦子三尖杉;萜類化合物;牻牛兒基牻牛兒基焦磷酸合酶(GGPPS);克隆;序列分析;同源模建;系統(tǒng)進(jìn)化
中圖分類號(hào): S188;Q785
文獻(xiàn)標(biāo)志碼: A
文章編號(hào):1002-1302(2016)04-0079-04
萜類化合物是植物代謝中數(shù)量最多的一類代謝物,以異戊二烯為結(jié)構(gòu)單元,在植物的光合作用、呼吸作用、生長(zhǎng)、發(fā)育、繁殖、信號(hào)轉(zhuǎn)導(dǎo)和防御中發(fā)揮重要作用。植物中存在的某些萜類具有重要的經(jīng)濟(jì)價(jià)值和藥用價(jià)值,如紅豆杉(Taxus)中的二萜紫杉醇[1]、銀杏(Ginkgo biloba)中的二萜銀杏內(nèi)酯和倍半萜白果內(nèi)酯[2-3]、青蒿(Artemisia annua)中的倍半萜青蒿素[4]等。三尖杉屬(Cephalotaxus)植物主要含有生物堿、黃酮類化學(xué)成分,其中的三尖杉酯類生物堿由于具有明顯的抗癌活性得到了深入研究,關(guān)于黃酮類成分的研究較少。除此之外還含有一些萜類化合物,如二萜類的海松酸、柳杉酚等,初步研究表明,該類化合物對(duì)微生物有抑制作用[5]。牻牛兒基牻牛兒基焦磷酸(geranylgeranyl diphosphate,GGPP)是包括紫杉醇、銀杏內(nèi)酯、海松酸、柳杉酚在內(nèi)的所有二萜類化合物的共同前體,由牻牛兒基牻牛兒基焦磷酸合成酶(geranylgeranyl pyrophosphate synthase,GGPPS)催化,通過(guò)類異戊二烯合成途徑生成。在該反應(yīng)途徑中,GGPPS催化15 C的法呢基焦磷酸(farnesylpyrophosphate,F(xiàn)PP)與5 C的異戊烯焦磷酸(isopentenylallyl diphosphate,IPP)縮合生成GGPP[6-7]。因此,GGPPS是紅豆杉植物合成紫杉醇、銀杏合成銀杏內(nèi)酯、三尖杉屬植物合成海松酸和柳杉酚的關(guān)鍵酶之一。在三尖杉中克隆該基因并分析其序列的結(jié)構(gòu)特征和進(jìn)化模式,將為后續(xù)的功能研究和基因工程、代謝工程等技術(shù)手段大量獲得有用產(chǎn)物奠定基礎(chǔ)。
1 材料與方法
1.1 試驗(yàn)材料
1.1.1 植物材料 篦子三尖杉(Cephalotaxus oliveri)摘采于中國(guó)科學(xué)院武漢植物園,取其嫩葉立即置于液氮中保存?zhèn)溆谩?/p>
1.1.2 主要分子生物學(xué)試劑 普通Taq DNA聚合酶、高保真 LA Taq 酶、dNTP等試劑,購(gòu)自TaKaRa;DNA凝膠純化回收試劑盒,購(gòu)于Axygen生物技術(shù)有限公司。
1.2 試驗(yàn)方法
1.2.1 篦子三尖杉GGPPS基因的克隆
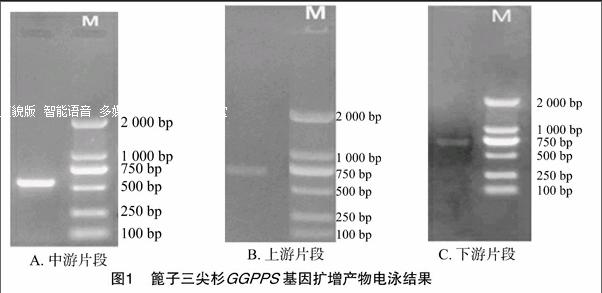
1.2.1.1 引物的設(shè)計(jì) 用加拿大紅豆杉 (Taxus canadensis) GGPPS蛋白(登錄號(hào):AADl6018)作為查詢序列,進(jìn)行Blastp相似性搜索,在三尖杉屬植物海南粗榧(C. hainanensis)中得到1條同目標(biāo)序列相似性較高的序列 (登錄號(hào):GBHQ10039.1)。根據(jù)這條序列設(shè)計(jì)3對(duì)特異引物(分別位于中部和左右端部),詳見表1。
1.2.1.2 篦子三尖杉GGPPS基因的分段擴(kuò)增 以提取的篦子三尖杉總DNA為模板,分別以F1-1、R1-2,F(xiàn)2-1、R2-2,F(xiàn)3-1、R3-2為引物進(jìn)行PCR擴(kuò)增。擴(kuò)增體系為25 μL,包含2.5 μL 10×PCR buffer(Mg2+ plus),2 μL dNTP混合物(每種2.5 mmol/L),各1 μL引物(10 μmol/L),0.5 μL Ex-Taq(2 U/μL),1 μL DNA模板(25 ng/μL),補(bǔ)加17 μL的水。反應(yīng)條件為:94 ℃ 5 min;94 ℃ 1 min,53 ℃ 1 min,72 ℃ 1 min,36個(gè)循環(huán);72 ℃延伸8 min。
PCR結(jié)束后,進(jìn)行DNA瓊脂糖凝膠電泳,對(duì)擴(kuò)增的DNA進(jìn)行檢測(cè)。
1.2.2 產(chǎn)物回收、測(cè)序 擴(kuò)增片段的凝膠回收按照試劑盒說(shuō)明書進(jìn)行操作,將回收的目的片段送華大基因測(cè)序。
1.3 數(shù)據(jù)分析方法
采用BioEdit(North Carolina State University) 軟件對(duì)克隆測(cè)序得到的片段進(jìn)行拼接。采用ORF finder進(jìn)行開放閱讀框確定。運(yùn)用DNAstar軟件推測(cè)氨基酸序列,運(yùn)用分析蛋白質(zhì)物理化學(xué)的Expasy Protparam軟件(http://web.expasy.org/protparam)分析蛋白質(zhì)的分子量、理論等電點(diǎn)、氨基酸組成。查詢Prosite數(shù)據(jù)庫(kù)查找蛋白質(zhì)含有的序列模式和功能域,使用在線軟件SOPMA程序(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)預(yù)測(cè)蛋白質(zhì)二級(jí)結(jié)構(gòu)并計(jì)算各種二級(jí)結(jié)構(gòu)所占百分比。使用SWISS-MODEL服務(wù)器[8-10] (http://swissmodel.expasy.org/),基于同源建模的原理,進(jìn)行篦子三尖杉GGPPS三級(jí)結(jié)構(gòu)的預(yù)測(cè),預(yù)測(cè)結(jié)果用PyMOL軟件展示和編輯。
以篦子三尖杉GGPPS蛋白作為查詢序列,運(yùn)用Blast工具(http//blast.ncbi.nlm.nih.gov/Blast.cgi)進(jìn)行序列相似性搜索,并選取相似性較高的序列用MEGA 6軟件[11]中的鄰接算法進(jìn)行進(jìn)化樹的構(gòu)建,其中多重序列比對(duì)使用Clustalx軟件進(jìn)行。
2 結(jié)果與分析
2.1 篦子三尖杉GGPPS序列的獲得與分析
采用改良的CTAB 法獲取篦子三尖杉總DNA,并通過(guò)電泳進(jìn)行純度檢測(cè),結(jié)果見圖1。用引物F2-1、R2-2進(jìn)行PCR擴(kuò)增,最終獲得560 bp左右的三尖杉GGPPS的中間片段(圖1-A)。分別以引物F1-1、R1-2,F(xiàn)3-1、R3-2進(jìn)行PCR擴(kuò)增,最終分別獲得 700、750 bp左右的上、下游片段(圖1-B、1-C)。產(chǎn)物經(jīng)回收純化和測(cè)序,用BioEditor軟件進(jìn)行拼接,獲得1條總長(zhǎng)為1 217 bp的GGPPS DNA序列。用ORF Finder軟件預(yù)測(cè)開放閱讀框(ORF),結(jié)果顯示:該DNA序列包含1段1 182 bp的完整編碼區(qū),編碼393個(gè)氨基酸。將獲得的基因組DNA序列與已報(bào)道的海南粗榧mRNA進(jìn)行比對(duì)發(fā)現(xiàn),篦子三尖杉GGPPS基因不含有內(nèi)含子,與大多數(shù)植物一樣,在外顯子-內(nèi)含子的進(jìn)化上是保守的。
2.2 篦子三尖杉GGPPS蛋白質(zhì)序列結(jié)構(gòu)特征分析
篦子三尖杉GGPPS基因完整的開放閱讀框(ORF)長(zhǎng)為1 182 bp,編碼393個(gè)氨基酸(圖2),相對(duì)分子質(zhì)量約為 43.042 ku,理論等電點(diǎn)為6.57。在氨基酸序列中,含量最高的為丙氨酸(Ala,39個(gè))、亮氨酸(Leu,39個(gè)),均占9.92%;其次是賴氨酸(Lys,33個(gè),占8.40%)、天門冬氨酸(Asp,29個(gè),占7.28%)、蘇氨酸(Thr,27個(gè),占6.87%)、纈氨酸(Val,27個(gè),6.87%)。酸性氨基酸[天門冬氨酸(Asp)、谷氨酸(Glu)]總數(shù)52個(gè),堿性氨基酸[精氨酸(Arg)、賴氨酸(Lys)]50個(gè);總親水性平均系數(shù)為-0.147。Prosite數(shù)據(jù)庫(kù)比對(duì)發(fā)現(xiàn),三尖杉GGPPS蛋白序列的177~193氨基酸位(氨基酸序列:LIhDDlpcmDnddfRRG)和311~323氨基酸位(氨基酸序列:VGllFQVvDDIlD)分別是2個(gè)多聚異戊二烯基合成酶的特異序列(圖3)。
SOPMA分析表明,α-螺旋(alpha helix,Hh)、無(wú)規(guī)則卷曲(random coil,Cc)是三尖杉GGPPS蛋白質(zhì)二級(jí)結(jié)構(gòu)的主要成分,組成α-螺旋、無(wú)規(guī)則卷曲、延伸鏈氨基酸比例分別為45.80%、41.48%、12.72%,不含其他結(jié)構(gòu)。蛋白質(zhì)三級(jí)結(jié)構(gòu)是在二級(jí)結(jié)構(gòu)的基礎(chǔ)上進(jìn)一步盤繞、折疊形成的,決定了蛋白質(zhì)的功能。在SWISS-MODEL服務(wù)器上預(yù)測(cè)篦子三尖杉GGPPS蛋白的三維結(jié)構(gòu),結(jié)果顯示,與薄荷(Menthax piperita)GGPPS蛋白同源性最高(PDB ID:3krp.2.A),為68.37%。以此結(jié)構(gòu)為模型構(gòu)建篦子三尖杉的三級(jí)結(jié)構(gòu),發(fā)現(xiàn)該蛋白具有植物GGPPS蛋白典型的三維結(jié)構(gòu),說(shuō)明功能也非常類似,該蛋白在進(jìn)化上很保守 (圖4)。
2.3 GGPPS蛋白質(zhì)同源性分析與系統(tǒng)進(jìn)化樹構(gòu)建
Blast比對(duì)結(jié)果顯示,三尖杉GGPPS蛋白質(zhì)序列與已報(bào)道的海南粗榧(C. hainanensis)、加拿大紅豆杉(T. Canadensis)、曼地亞紅豆杉(Taxus xmedia)、挪威云杉(Picea abies)、銀杏(Ginkgo biloba)、煙草(Nicotiana tabacum)、北美冷杉(Abies grandis)、馬尾松(Pinus massoniana)、橡膠樹(Hevea brasiliensis)、巴豆(Croton sublyratus)、大豆(Glycine max)、美花煙草(Nicotiana sylvestris)等12種高等植物GGPPS蛋白質(zhì)的一致性分別為98.9%、89.5%、88.8%、70.4%、70.1%、54.3%、69.4%、67.6%、54.0%、54.5%、54.4%、54.5%。多序列同源比對(duì)分析結(jié)果表明,不同植物來(lái)源的GGPPS蛋白質(zhì)的N末端差異很大,C末端保守性強(qiáng)。GGPPS蛋白在結(jié)構(gòu)和功能上屬于多聚異戊二烯合成酶家族,該家族具有5個(gè)保守功能域(Ⅰ~Ⅴ),5個(gè)保守區(qū)域中包含2個(gè)富含天冬氨酸區(qū)域:DDXXXXD、DDXXD。在篦子三尖杉GGPPS氨基酸序列中存在這樣的5個(gè)保守結(jié)構(gòu)域,結(jié)構(gòu)域Ⅱ、結(jié)構(gòu)域Ⅴ中也分別發(fā)現(xiàn)富含天冬氨酸的基序DDLPCMD、DDILD(圖5)。
采用Neighbor-joining算法,在MEGA 6軟件平臺(tái)上構(gòu)建植物GGPPS的分子系統(tǒng)進(jìn)化樹(圖6)。對(duì)比植物GGPPS分子系統(tǒng)進(jìn)化樹,三尖杉GGPPS蛋白歸屬于裸子植物進(jìn)化支,且與海南粗榧、加拿大紅豆杉、挪威云杉、北美冷杉、馬尾松的GGPPS蛋白歸為同一小分支(圖6)。被子植物、裸子植物GGPPS各形成獨(dú)立的分支,兩者形成姊妹關(guān)系,說(shuō)明這兩大分支從同一祖先平行分化而來(lái);該基因樹與物種樹基本吻合,說(shuō)明該基因的進(jìn)化與物種的分化基本一致。
3 結(jié)論
通過(guò)PCR技術(shù)克隆獲得的篦子三尖杉GGPPS DNA序列長(zhǎng)度為1 217 bp,含有1 182 bp的完整開放閱讀框。三維結(jié)構(gòu)模擬表明:該蛋白具有GGPPS蛋白典型的三維結(jié)構(gòu);多重序列比對(duì)顯示,三尖杉GGPPS 蛋白與其他物種來(lái)源的GGPPS蛋白相似性較高,尤其是與海南粗榧、加拿大紅豆杉、曼地亞紅豆杉的GGPPS相似性高達(dá)88%以上,它們都包含多聚異戊二烯合成酶家族的5個(gè)保守性結(jié)構(gòu)域(圖5)。在三尖杉GGPPS蛋白的氨基酸序列的177~193、311~323位分別是2個(gè)多聚異戊二烯基合成酶的特異序列LIhDDlpcmDnddfRRG、VGllFQVvDDIlD(圖3),這2個(gè)保守的富含天冬氨酸的區(qū)域位于酶的催化活性位點(diǎn),與IPP、烯丙基底物的結(jié)合有關(guān)。從結(jié)構(gòu)到功能預(yù)測(cè)都顯示,克隆的基因具有典型的GGPPS特征,因此克隆的篦子三尖杉GGPPS編碼蛋白很可能參與或調(diào)節(jié)了三尖杉二萜類等物質(zhì)的生物合成,但完全闡明三尖杉萜類化合物的生物合成和調(diào)控機(jī)制還需要進(jìn)行進(jìn)一步試驗(yàn)驗(yàn)證。
參考文獻(xiàn):
[1]占愛瑤,由香玲,詹亞光. 植物萜類化合物的生物合成及應(yīng)用[J]. 生物技術(shù)通訊,2010,21(1):131-135.
[2]朱 俊,許 鋒,廖詠玲. 銀杏萜內(nèi)酯調(diào)控研究進(jìn)展[J]. 中國(guó)農(nóng)學(xué)通報(bào),2007,23(7):301-305.
[3]張洪娟,譚碧玥,曹福亮. 銀杏GbGGPS基因的克隆及序列分析[J]. 南京林業(yè)大學(xué)學(xué)報(bào):自然科學(xué)版,2013,37(4):8-12.
[4]陳大華,葉和春,李國(guó)鳳,等. 植物類異戊二烯代謝途徑的分子生物學(xué)研究進(jìn)展[J]. 植物學(xué)報(bào),2000,42(6):551-558.
[5]Politi M,Braca A,de Tommasi N,et al. Antimicrobial diterpenes from the seeds of Cephalotaxus harringtonia var. drupacea[J]. Planta Medica,2003,69(5):468-470.
[6]Lange B M,Rujan T,Martin W,et al. Isoprenoid biosynthesis:the evolution of two ancient and distinct pathways across genomes[J]. Proceedings of the National Academy of Sciences of the United States of America,2000,97(24):13172-13177.
[7]Rodríguez-Concepción M,Boronat A. Elucidation of the methylerythritol phosphate pathway for isoprenoid biosynthesis in bacteria and plastids. A metabolic milestone achieved through genomics[J]. Plant Physiology,2002,130(3):1079-1089.
[8]Biasini M,Bienert S,Waterhouse A,et al. SWISS-MODEL:modelling protein tertiary and quaternary structure using evolutionary information[J]. Nucleic Acids Research,2014,42(W1):W252-W258.
[9]Arnold K,Bordoli L,Kopp J,et al. The SWISS-MODEL workspace:a web-based environment for protein structure homology modelling[J]. Bioinformatics,2006,22(2):195-201.
[10]Benkert P,Biasini M,Schwede T. Toward the estimation of the absolute quality of individual protein structure models[J]. Bioinformatics,2011,27(3):343-350.
[11]Tamura K,Stecher G,Peterson D,et al. MEGA6:molecular evolutionary genetics analysis version 6.0[J]. Molecular Biology and Evolution,2013,30(12):2725-2729.趙金艷,盧 宏,張振武,等. 快速PCR介導(dǎo)的NeuroD-3′UTR的定點(diǎn)突變研究[J]. 江蘇農(nóng)業(yè)科學(xué),2016,44(4):83-86.
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級(jí)版(2021年3期)2021-03-19 06:12:40
小天使·二年級(jí)語(yǔ)數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
