基于拉曼光譜分析寒地水稻葉片的有機結構
2016-06-14 17:12:12譚峰才巧玲馬志欣孫雪成侯召龍
江蘇農業科學 2016年4期
譚峰+才巧玲+馬志欣+孫雪成+侯召龍

摘要:為了從分子水平探討水稻病菌的傳播和發病機制,特別是與分子病理學和分子藥物學有關的研究項目的開展,需要研究細胞內生物大分子的結構及結構與功能的關系。利用拉曼光譜儀對正常水稻葉片進行拉曼光譜采集,指認出水稻葉片細胞中重要組成成分的拉曼特征頻率,如羰基、亞甲基、巰基、氨基等多種官能團,并對拉曼光譜曲線中出現的特殊結構進行理論分析和解釋。然后針對細胞組成的兩大成分,即蛋白質和遺傳物質的結構構像進行相應的拉曼光譜分析。通過分析水稻葉片中各部分的二級結構構象給出全面的分析和解釋,實現了水稻葉片半定量分析的目的。為今后寒地水稻病害早期檢測的對比分析提供了理論基礎。
關鍵詞:拉曼光譜;蛋白質;遺傳物質;官能團;二級結構
中圖分類號: S511.01
文獻標志碼: A
文章編號:1002-1302(2016)04-0358-03
寒地水稻是指東北地區所產的單季晚稻品種。水稻病害是影響寒地水稻產量的重要因素之一,每年均有病害發生,嚴重時可以減產40%~50%[1-2]。研究水稻病害內部發病機制以及對水稻葉片結構破壞的機制具有重要意義。對水稻葉片檢測通常采用透射電鏡、多光譜成像技術、高光譜成像技術等方法,這些方法已經能夠較準確地識別水稻病害情況,但仍存在識別方法普適性差、無損檢測方法缺失的問題[3]。拉曼光譜是一門基于拉曼散射效應而發展起來的光譜分析技術,體現的是分子的振動或轉動信息,與常規化學分析方法相比,拉曼光譜技術具有無損、快速、環保、靈敏度高、穩定性好且無需制樣等特點[4-5]。崔向榮等利用拉曼光譜技術檢測骨髓間充質干細胞在電磁場照射后蛋白質、脂類、核酸等結構和數量的變化的可行性,得出3 000 Hz 脈沖電磁場對骨髓間充質干細胞產生了分子層面的影響[6]。Waleed等利用拉曼光譜技術檢測PC12細胞裂解液在細胞內DA水平的生化成分的變化[7],同時確定拉曼光譜在檢測細胞外DA水平處理劑的變化與運用高效液相色譜法結果一致。董蓉等應用單細胞拉曼分析技術結合PCA方法,以枯草芽孢桿菌芽孢為對象,探究芽孢在強堿脅迫下的致死機制[8]。由此可見,利用拉曼光譜技術在生物、農業等方面的應用越來越廣泛,已經成為一種有效的檢測手段。但目前利用拉曼光譜對水稻植株檢測的研究較少。因此,本研究利用拉曼光譜和水稻葉片細胞組織的知識理論進行研究,以期為水稻病害早期檢測的對比分析提供一定的理論基礎。
1 材料與方法
1.1 樣品的采集
水稻葉片樣本2011—2014年采集于黑龍江省農墾總局建三江分局下屬勝利農場科技示范園區的26.67 hm2試驗田,品種為墾稻19號。水稻是蒸騰作用非常強的農作物,正常天氣采集下來的水稻葉片10 min之內便會因為葉片水分降低出現明顯的卷葉現象,所以采集樣本必須現場采摘葉片,并直接在附近的室內進行拉曼光譜采集。
1.2 試驗方法
選用由DeltaNu公司生產的Advantage 532型拉曼光譜儀,采用背向接收的方式采集拉曼光譜信號,在波段200~3 400 cm-1 范圍內對現場樣本進行拉曼光譜采集。采集到的原始數據,一是利用編寫的Matlab程序進行數據處理,得到樣本光譜的峰值,并顯示處理以后的光譜圖像;二是通過結合Berjot方法針對二級碳、三級碳、四級碳以及譜線展寬與水稻葉片內結構進行關聯分析,對照拉曼光譜特征頻率表,得到寒地水稻葉片中大部分關鍵化學鍵的拉曼光譜分析結果[9]。
2 結果與分析
2.1 細胞中重要成分在拉曼光譜中的分析
利用拉曼光譜分析儀在中等激光強度、532 nm波長、曝光時間1 s、無偏振檢測的條件下采集正常水稻葉片的拉曼光譜,采集波數范圍為200~3 400 cm-1。幾丁質結合蛋白是在植物中主要存在的物質,其中半胱氨酸(cysteine)是幾丁質結合域的主要特征。半胱氨酸分子式為HSCH2CH(NH2)COOH,它含有羰基—CO、亞甲基—CH2—、巰基—SH、氨基—NH2等多種官能團。446、535 cm-1分別對應半胱氨酸羧基(—COOH)的面內搖擺振動和面外搖擺振動,是區別于胱氨酸、乙酰半胱氨酸的標志特征峰。從峰值相對強度可以看出在水稻葉片中半胱氨酸含量相對較少,導致標志特征峰強度低。695 cm-1為半胱氨酸的C—S鍵,其實此處譜峰是693、 695 cm-1 峰的疊加譜峰,但由于儀器分辨率不高,故只能看到1個明顯的拉曼峰。990、1 043 cm-1為半胱氨酸中的甲基(—CH3),它們同屬于甲基的面內搖擺振動。同時可以看到 990 cm-1峰的強度偏弱,再次說明半胱氨酸的含量較少。1 043 cm-1附近峰是個椅形結構,在1 043 cm-1附近的幾個波數內是一種水平結構,這是由于受附近1 050 cm-1譜峰交疊影響而不能表現為突出的峰(圖1-a)。1 349、1 365、1 385、1 392、1 412 cm-1等為半胱氨酸分子中—CO的伸縮振動與—O—H鍵偶合的拉曼譜峰,在這些拉曼譜峰中以波數 1 392 cm-1峰相對強度最大。1 348、2 966 cm-1為半胱氨酸中的亞甲基(—CH2)的伸縮振動和變形振動(圖1-b)。
1 680~1 710 cm-1為巰基(—SH)在周圍不飽和鍵化學環境影響下的吸收峰;1 700~1 640 cm-1為飽和鍵化學環境影響下的吸收峰。2 498 cm-1為巰基的伸縮振動,說明在植物葉片中具有巰基的存在;同時,處在不飽和巰基頻率范圍段內的峰值有1 687、1 699、1 710 cm-1等3個;在飽和巰基頻率范圍段內的峰值有1 641、1 653、1 670、1 687、1 699 cm-1 等5個(圖1-c、圖1-d)。
2.2 水稻蛋白質的主鏈構象與二級結構分析
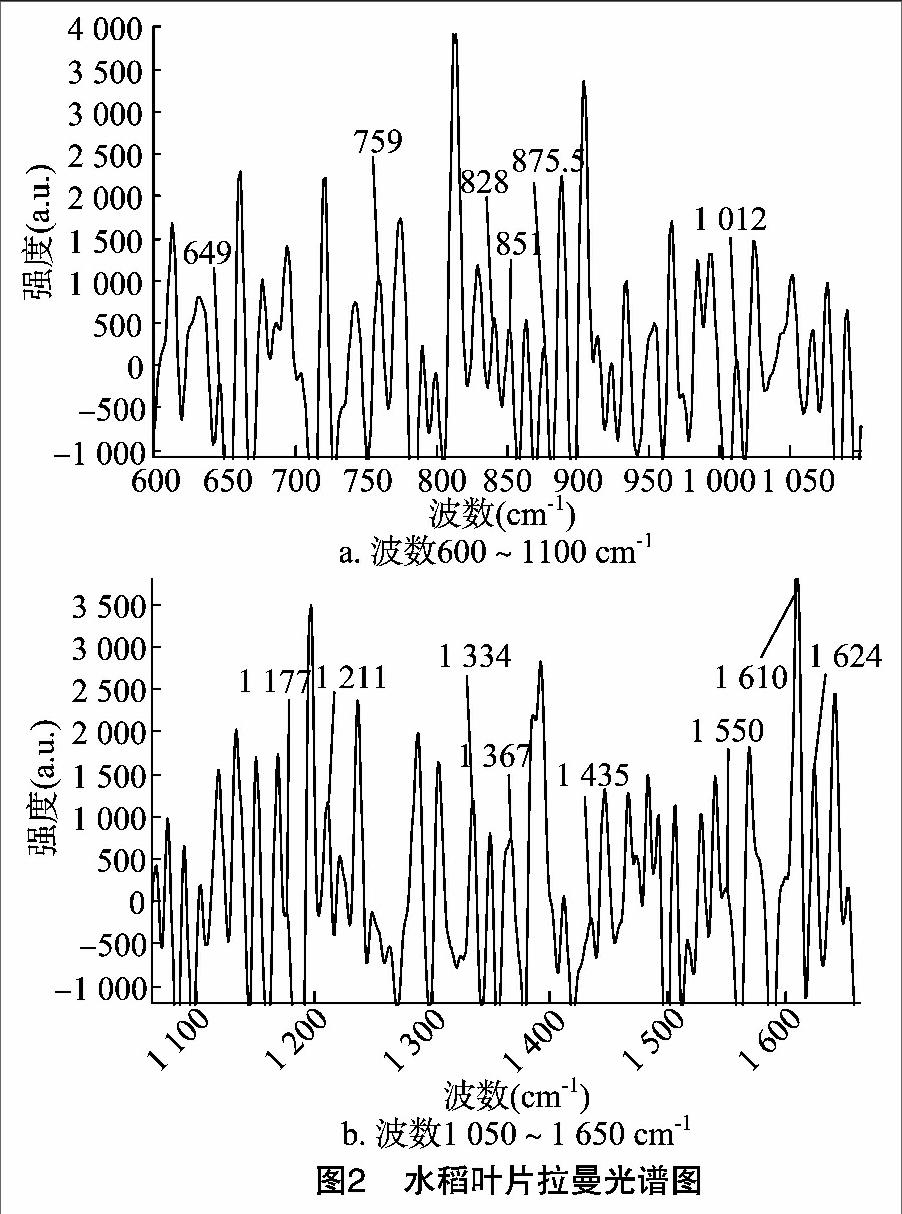
從水稻葉片的拉曼光譜中可以得到許多可貴的信息,包括葉片蛋白質中類似于芳香族組成的氨基酸的信息,進一步得到的二級結構的信息。蛋白質的主鏈構象的拉曼光譜復雜多樣,但它們主要由骨架C—C、C—N和肽鍵(—CONH—)引起。C—C伸縮振動的譜線,α-螺旋區域在890~945 cm-1,無規卷曲區域在945~960 cm-1,且構象靈敏;而C—N伸縮振動在1 070~1 130 cm-1,且構象靈敏。蛋白質中的肽鍵可以引起多種類型的振動模式,分別叫做酰胺Ⅰ、酰胺Ⅱ、酰胺Ⅲ、酰胺Ⅳ。其中酰胺Ⅰ是肽鍵中的羰基(—CO)伸縮振動,在自由結構中的譜峰在1 670~ 1 700 cm-1,在結合結構中在1 630~1 680 cm-1;酰胺Ⅲ是C—N鍵的伸縮振動,在拉曼光譜中為強譜線,譜峰所在位置在1 290 cm-1附近。它們對蛋白質的構象變化十分敏感,而酰胺Ⅰ峰在拉曼譜圖上明顯,但是拉曼譜圖上的酰胺Ⅱ峰及NH峰非常弱以致不好辨識。在研究蛋白質的及多肽結構時,由于譜峰過多,酰胺Ⅱ和酰胺Ⅳ就起不到作用。蛋白質結構中某些氨基酸含有C—S鍵,它有2種構象,其中632、646.5、661 cm-1為含有扭曲構象的C—S鍵;704、721、743 cm-1為含有反式構象的C—S鍵。1 410 cm-1附近為羧基(—COOH)對稱伸縮振動;2 550~2 590 cm-1 之間存在巰基(—SH) (圖2)。
851、828 cm-1的雙峰為酪氨酸(C9H11NO3)對羥苯基環的呼吸振動和環平面外彎曲振動的倍頻之間的費米共振引起的,且雙峰隨側鏈微環境而改變。現在的強度比值I851/828=30 968/31 854=0.972,該值小于1,說明該酪氨酸的部分是埋藏在蛋白質分子內部。對酪氨酸殘基進行計算,其中酪氨酸的殘基滿足如下表達式:
N埋+N露=1;
0.5N埋+1.25N露=I851/828。(1)
得到N露≈0.63,N埋≈0.37。但需要注意,因為水稻葉片光譜采集在有熒光處采集而非在暗室中采得,計算所得酪氨酸殘基誤差相對較大,但是在有誤差的情況下說明了一定的蛋白質構型問題。與酪氨酸對應的譜線還有647、1 177、1 211、1 610、1 624 cm-1等5條譜線[9]。
色氨酸中具有吲哚環。吲哚是吡咯與苯并聯的化合物,別稱苯并吡咯,分子式為C8H7N。2種并合方式,分別稱為吲哚和異吲哚。吲哚是一種亞胺,具有弱堿性;雜環的雙鍵一般不發生加成反應;在強酸的作用下可以生發二聚合和三聚合作用;在特殊的條件下,能進行芳香親電取代反應。色氨酸的吲哚環引起的譜線分別在759、875.5、1 012、1 334、1 367、1 550、1 624 cm-1附近。其中吲哚環的N—H彎曲振動譜線在 1 435 cm-1 處,而1 367 cm-1的譜線對于周圍化學結構環境的改變是非常敏感的,可以看出的譜線是鈍形峰,說明色氨酸中的吲哚還是暴露在蛋白質結構以外,如果色氨酸殘基是埋藏的,該譜線會呈尖銳峰形(圖2)。
1 004 cm-1為苯丙氨酸的單基取代苯環是引人注意的強峰,但它的構象不靈敏,如果蛋白質沒有共價鍵斷裂,可以作為內標,屬于苯丙氨酸殘基的譜線還有624、1 609、1 624 cm-1等3條譜線。如果在純蛋白質水溶液中,還有1 032、 1 203 cm-1 等2條譜線,但是由于殘基受到其他基團的影響,在水稻葉片的蛋白質拉曼光譜中沒有得到體現。S—S鍵有3種構象,在(510±5) cm-1出現的譜線屬于扭曲-扭曲-扭曲構象(gauche-gauche-gauche),在(525±5) cm-1的譜線屬于扭曲-扭曲-反式構象(gauche-gauche-trans),在(540±5) cm-1的譜線屬于反式-扭曲-反式構象(trans-gauche-trans)。
2.3 水稻核蛋白及遺傳物質的二級結構分析
雖然組蛋白是蛋白質成分的一部分,根據細胞組成來看,也不宜作為純蛋白質進行分析,所以將其歸入遺傳物質進行拉曼光譜分析。核小體是染色體的基本結構單位,由DNA和組蛋白構成。其中,DNA是細胞的主要遺傳物質,是一種長鏈聚合物,由4種脫氧核苷酸,即腺嘌呤、鳥嘌呤、胞嘧啶和胸腺嘧啶組成,一般由C、H、O、N、P等5種元素組成。1個脫氧核苷酸分子由3個分子組成,即含氮堿基、脫氧核糖和磷酸。而糖類和磷酸分子借由酯鍵相連,組成長鏈骨架。由于細胞核內相對濕度比較大,所以DNA的構象是一種含水量大的構象形式,其特征譜線大約在840 cm-1波數上。脫氧核酸的譜線有4根,分別為1 446、1 467、924、984 cm-1。脫氧核糖的譜線有934、1 012、1 051、1 066、1 446 cm-1等5條,其中 1 012、1 051 cm-1 的譜線屬于脫氧核糖的C—O伸縮振動,721、1 223、1 306、1 335、1 505 cm-1屬于腺嘌呤一些基團的譜線。1 492 cm-1 屬于鳥嘌呤的一些基團的譜線。1 528、1 385、1 262 cm-1 的譜線分別屬于腺嘌呤和胞嘧啶,胸腺嘧啶、胞嘧啶和鳥嘌呤,以及鳥嘌呤和胞嘧啶的一些基團。屬于包括堿基之間氫鍵在內的胸腺嘧啶羰基的伸縮振動的譜線在 1 670 cm-1 處,當堿基之間的氫鍵斷裂時該譜線會向低波數位移。1 197 cm-1的譜線屬于堿基外C—N伸縮振動。DNA骨架磷酸基團的譜線有2根,它們是在774 cm-1的磷酸二酯PO2對稱伸縮振動的譜線和1 089 cm-1磷酸離子對稱伸縮振動。
屬于堿基腺嘌呤、鳥嘌呤、胞嘧啶和胸腺嘧啶的譜線如下:(1)屬于腺嘌呤(A)的有720、1 250、1 305、1 334、1 426、1 492、1 517 cm-1;(2)屬于鳥嘌呤(G)的有686、1 325、1 426、1 492 cm-1;(3)屬于胞嘧啶(C)的有1 250 cm-1;(4)屬于胸腺嘧啶(T)的有661、1 211、1 237、1 653 cm-1。
RNA骨架磷酸基團有2根譜線。其中磷酸離子對稱伸縮振動在1 103.5 cm-1處譜線是構象不靈敏的,構象靈敏的磷酸二酯PO2對稱伸縮振動的譜線在813 cm-1處。若磷酸骨架處于有序的狀態時,會在813 cm-1處產生強的拉曼光譜。而若RNA的構象變為無序時它將移位到808~795 cm-1 之間的某個譜線處。對于RNA的4種堿基特征拉曼譜線與DNA中相一致。
碳水化合物是組成細胞內糖蛋白和糖脂的重要成分,它們在細胞通信和供應細胞活動所需的能源方面扮演了重要的角色。對于能以高度支鏈化形式及α、β-糖苷異構形式存在的寡糖鏈,其種類和連接方式非常多,它能以最小的結構單位承載最大的生物信息量而成為最理想的信息載體。糖蛋白的糖類部分與細胞的抗原結構、受體、細胞免疫、細胞識別等有密切關系。根據譜圖分析,獲得該細胞內的部分碳水化合物的信息:(1)屬于葡萄糖的有550、743、1 025、1 051、1 076、1 119、1 349 cm-1;(2)屬于D-甘露糖的有743、924、955、1 076、1 103 cm-1;(3)屬于葡萄糖醛酸的有564、743、1 025、1 051、1 076、1 119、1 349 cm-1;(4)屬于N-乙酰基葡萄糖的有743、984、1 025 cm-1。
3 結論與討論
本研究從細胞的化學成分入手,分析了包括羰基—CO、亞甲基—CH2—、巰基—SH、氨基—NH2等多種官能團的拉曼特征振動頻率,并對光譜曲線中出現的特殊結構(如椅形結構)進行理論分析和解釋。然后針對細胞組成的兩大成分蛋白質和遺傳物質的結構構像進行相應的拉曼光譜分析。在蛋白質分析過程中,分析酪胺酸中埋藏的和暴露的殘基的比例關系,分析C—C單鍵伸縮振動的α-螺旋、β-折疊、β-回折和無規卷曲方面的信息,同時著重對于氨基酸縮合肽鍵的酰胺Ⅰ、Ⅱ、Ⅲ、Ⅳ譜線進行較為細致的分析。遺傳物質是每個生物體所具有重要結構,本研究針對核小體以及組成DNA的脫氧核苷酸進行構型分析,同時也分析了RNA的骨架磷酸基團的特征拉曼譜線;并總結得出水稻葉片中的腺嘌呤、鳥嘌呤、胞嘧啶和胸腺嘧啶的多個對應譜峰。針
對糖蛋白和糖脂的重要組成成分,分析了碳水化合物中葡萄糖、D-甘露糖、葡萄糖醛酸、N-乙酰基葡萄糖等4種構象的拉曼特征譜線。由于植物細胞的結構復雜程度不可能將立體結構完全進行構象分析,所以通過水稻葉片中各種部分的二級結構構象分析,實現了水稻葉片半定量分析的目的,為今后拉曼光譜檢測寒地水稻病害對比分析提供了理論基礎。
參考文獻:
[1]歐全宏,趙興祥,周湘萍,等. 稻瘟病、玉米銹病和蠶豆銹病葉的傅里葉變換紅外光譜研究[J]. 光譜學與光譜分析,2012,32(9):2389-2392.
[2]譚 峰,汪 春,尚廷義. 基于近紅外光譜的寒地水稻稻瘟病檢測數據分析[J]. 農機化研究,2011,33(11):44-47.
[3]楊 燕,何 勇. 基于高光譜圖像的稻瘟病抗氧化酶值早期預測[J]. 農業工程學報,2013,29(20):135-141.
[4]周麗娜,于海業,張 蕾,等. 基于葉綠素熒光光譜分析的稻瘟病害預測模型[J]. 光譜學與光譜分析,2014,34(4):1003-1006.
[5]鄒 喬,杜顯元,張 琛,等. 飽和一元醇類分子拉曼光譜振動峰的歸屬研究[J]. 光譜學與光譜分析,2013,33(1):106-110.
[6]崔向榮,蘇 偉,吳智輝,等. 中頻脈沖電磁場照射骨髓間充質干細胞的拉曼光譜[J]. 中國組織工程研究,2014,18(37):5929-5934.
[7]El-Said W A,Choi J W. In-situ detection of neurotransmitter release from PC12 cells using Surface Enhanced Raman Spectroscopy[J]. Biotechnology & Bioprocess Engineering,2014,19 (6):1069-1076.
[8]董 蓉,盧明倩,李 鋒,等. 強堿脅迫枯草芽孢桿菌芽孢致死的拉曼光譜研究[J]. 光譜學與光譜分析,2013,33(9):2416-2420.
[9]迪 安. 分析化學手冊[M]. 北京:科學出版社,2003.田 上,沙之敏,岳玉波,等. 不同類型溝渠對農田氮磷流失的攔截效果[J]. 江蘇農業科學,2016,44(4):361-365.
