不同鉛水平下紫莖澤蘭細胞內鉛的分布和化學形態的分析
2016-06-23 10:37:44劉小文齊成媚顏冬冬王秋霞郭美霞曹坳程
廣西植物 2016年3期
劉小文, 齊成媚, 李 園, 顏冬冬, 王秋霞, 郭美霞, 曹坳程*
( 1. 湖南科技學院 化學與生物工程學院, 湖南 永州 425199; 2. 植物病蟲害生物學國家重點實驗室,北京 100193; 3. 中國農業科學院植物保護研究所, 北京 100193 )
不同鉛水平下紫莖澤蘭細胞內鉛的分布和化學形態的分析
劉小文1,2,3, 齊成媚1, 李園2,3, 顏冬冬2,3, 王秋霞2,3, 郭美霞2,3, 曹坳程2,3*
( 1. 湖南科技學院 化學與生物工程學院, 湖南 永州 425199; 2. 植物病蟲害生物學國家重點實驗室,北京 100193; 3. 中國農業科學院植物保護研究所, 北京 100193 )
摘要:該研究運用差速離心法和化學試劑逐步提取法,分析了重金屬鉛在紫莖澤蘭亞細胞內的分布和主要化學形態。結果表明:隨著Pb濃度的升高,紫莖澤蘭的葉、根、莖中各亞細胞組分Pb含量逐漸增加;紫莖澤蘭中的Pb在葉片分布于可溶性部分和細胞壁中,兩者占總量的75.34%~84.63%;莖也主要分布于可溶性部分和細胞壁中,占總量的36.10%~57.14%和20.07%~36.52%;而在根中則富集于細胞壁和可溶性部分,分別占39.2%~49.78%和28.27%~37.62%,其他細胞器中的Pb含量均較少。紫莖澤蘭葉中的Pb以鹽酸提取態和水提取態為主,兩者占總量的58.74%~73.04%;莖中的Pb以醋酸提取態和氯化鈉提取態為主;而根中的Pb則以醋酸提取態和鹽酸提取態占優勢,兩者占總量的39.15%~52.91%。
關鍵詞:鉛, 紫莖澤蘭, 亞細胞分布, 化學形態
重金屬污染是最為廣泛的土壤污染源之一,并逐步發展成污染生態學的研究熱點(劉秀梅和王慶仁, 2001)。植物修復是近年來發展迅速的一門環境污染修復技術,是以植物對某種或某些化學元素的耐性或超累積效應為基礎而作用于環境中的污染物,并能良好地利用植物與環境微生物之間的共存效應而成為一種新興的環境污染修復技術,對治理土壤重金屬污染產生了重要作用(Garbisu & Alkorta, 2001; Salt et al, 1995)。植物主要通過避性和耐性兩種方式來實現適應環境中的重金屬污染,避性是植物以自我調節的方式遠離不良的生存環境,從而免受或減輕重金屬污染對其危害性,如根系向重金屬含量低的部分生長,或是植物通過自身代謝將吸收的重金屬轉運至外環境中;耐性是植物體內通過一些生理變化,與重金屬結合呈現出解毒形態,如與植物細胞壁結合,或與植物細胞內合成的蛋白、多肽、有機酸反應而形成絡合物,通過改變重金屬原有的活性,使其形成無毒或毒性較弱的形態,再轉運到液泡內貯存等。Pb是植物生長的非必需元素,可通過影響植物的光合作用,破壞葉綠素的合成和抗氧化酶系統,濃度過高時能嚴重影響植物正常生理代謝功能,進而抑制植物生長發育,嚴重時可使植物死亡(Wu et al, 2003)。
紫莖澤蘭(Eupatoriumadenophorum)是原產于中美洲的菊科澤蘭屬多年生草本植物,現已擴散至亞洲、歐洲、非洲、大洋洲、北美洲等區域且已被諸多國家及權威研究機構公認為世界級惡性入侵雜草(牛燕芬等,2011)。該植物于20世紀40年代前后從中緬邊境以自然擴散的方式傳入我國云南,經半個多世紀發展后已蔓延至我國貴州、廣西、重慶、四川、湖北、西藏等地并以其對環境極強的適應性迅速建立起了種群優勢,嚴重危害了當地的自然環境和生物鏈平衡(賈海江等,2009)。鑒于紫莖澤蘭的危害性大,研究紫莖澤蘭的防控措施已迫在眉睫。目前,研究重金屬污染土壤的熱點是植物修復技術,且已證實紫莖澤蘭生長能力強、產量高,具備環境污染修復先鋒植物的特征(魯平等,2005;李雪瑤和應浩,2009)。紫莖澤蘭可有效吸收富集土壤中的Pb,且植株的不同部位對Pb的耐受能力呈現出很大差異(李冰等,2008;汪文云等,2008;Zu et al,2005)。筆者在調查中也發現,紫莖澤蘭可在重金屬Pb污染嚴重的環境中保持正常生長,且隨著土壤Pb含量不同,其生長狀況也明顯不同,紫莖澤蘭體內可能存在某些耐重金屬的機制。前期研究表明,紫莖澤蘭的生長狀態隨Pb處理水平表現出低促高抑的效應,且在高濃度Pb脅迫下,紫莖澤蘭各組織中Pb的累積量遠超過一般植物的累積水平。具備超累積效應的植物對重金屬的吸附能力跟植株在細胞和亞細胞水平上區隔重金屬的作用強弱有關(Kupper et al,1999)。研究重金屬在植物亞細胞分布將有助于揭示植株內重金屬的解毒形態和富集區的具體分布(Hall,2002)。因此,本研究以差速離心技術分離紫莖澤蘭各組織的亞細胞組分,并運用化學試劑逐步提取法對Pb在紫莖澤蘭細胞內的亞細胞分布特征和結合形態富集量進行了研究,擬揭示其對Pb的耐性和解毒機理,以期為研究紫莖澤蘭對Pb的生理響應和富集Pb的分子機理奠定基礎。
1材料與方法
1.1 植物培養
試驗在室內大棚中進行,場地環境為相對濕度(55±8)%、白天溫度(25±5)℃、夜間溫度(15±5)℃。場地土壤采自四川省西昌學院內實驗基地土壤表層,相關參數為pH值6.30(水)/7.71(CaCl2)、總N 853 mg·kg-1、總P 441 mg·kg-1、總K 2 313 mg·kg-1,CEC 11.23 meq、有機質含量25.1 mg·kg-1、重金屬Pb含量20 mg·kg-1。Pb脅迫處理水平為0(CK)、200、500、1 000 mg·kg-1,Pb以Pb(CH3COO)2的形式加入。經上述處理后將土壤充分拌勻,穩定2周后移植紫莖澤蘭幼苗。選取株高8~10 cm且質量接近的植株,隨機分配至每個濃度處理的土壤中,每個處理栽種16盆,每盆1株。移植完成后,用稱重法補充水分至田間持水量的60%左右并保持至收獲,重金屬脅迫處理60 d后,收獲紫莖澤蘭植株。植株先用自來水沖洗表面雜質后,再用20 mmol·L-1Na2-EDTA交換20 min,去除根系表面吸附的Pb2+,最后用去離子水洗凈,并用吸水紙吸干植株表面水分,于-20 ℃環境的冰箱中儲存。處理過的樣品組分為根、莖和葉片3個組分,依次測定重金屬全量、亞細胞分布和亞細胞內化學形態。
1.2 植物亞細胞組分的分離
將紫莖澤蘭各組織研磨勻漿后用差速離心法分離不同組織中的各亞細胞組分。參照Weige & Jager(1980)和Gabbrielli et al(1980)的方法,并針對處理方法的局限性加以完善:取樣0.200 0 g,加入8 mL提取液[0.25 mol·L-1蔗糖+50 mmol·L-1Tris-HCl緩沖液(pH7.5)+1 mmol·L-1二硫赤鮮糖醇],勻漿處理后,收集勻漿液后在300 r·min-1下冷凍離心30 s,至沉淀完全析出,過濾分離上清液,即為細胞壁組分(F1);收集離心樣品的上清液并在2 000 r·min-1下離心15 min,待沉淀穩定后分離上清液,沉淀部分即為細胞核和葉綠體組分(F2);收集細胞核和葉綠體沉淀后上清液在10 000 r·min-1下離心20 min,重復上述分離操作,所謂不溶物為線粒體組分(F3);收集經三次差速離心分離的上清液,為可溶組分(F4),各步驟均在4 ℃下進行。
1.3 紫莖澤蘭體內重金屬化學形態分析
采用不同試劑分步提取法逐步分離紫莖澤蘭植株內內重金屬各化學形態,方案如下:取樣0.200 0 g,加入8 mL提取劑研磨勻漿后轉入10 mL塑料離心管中,在25 ℃恒溫振蕩22 h后,5 000 r·min-1下離心10 min,倒出上清液,加入8 mL的提取劑,25 ℃下恒溫振蕩1 h,10 000 r·min-1離心10 min,取上清液混合后充分搖勻,采用下列5種提取劑依次逐步提取:80%乙醇(FE),去離子水(FW),1 mol·L-1NaCl溶液(FNaCl),2% HAc(FHAc),0.6 mol·L-1HCl(FHCl),最后為殘留態(FR)。
1.4 Pb含量的測定
以提取劑作為空白對照,所得樣品用去離子水洗凈后轉移至100 mL三角瓶中,將其蒸干后采用濃HNO3∶HF∶HClO4(2∶1∶2)法測定Pb的含量,先消煮至澄清,過濾后用去離子水定容,北京瑞利WFX-120A /石墨爐原子吸收分光光度計測定,所有數據用SAS 8.2軟件進行處理。
2結果與分析
2.1 紫莖澤蘭各組織中Pb的亞細胞分布
重金屬為避免損傷植物核心的細胞、組織和細胞器,從而體現對應的選擇性分配,形成了在亞細胞各組分中的分配差異,其主要累積區位于植物的細胞壁和液泡中,但由于植物種類、Pb處理水平等條件的不一致,會導致Pb在兩者之間分配比例存在差異。不同濃度的Pb處理下,Pb在紫莖澤蘭各組織的亞細胞分布及各組分所占比例如表1所示,在紫莖澤蘭的亞細胞組分分離樣品中,Pb的回收率為82.17%~97.65%,除對照中葉片外,紫莖澤蘭葉片Pb的亞細胞組分的含量均呈現F4>F1>F3>F2的分布格局,即均以含可溶性部分F4分布最高,占28.72%~54.23%,其次是細胞壁F1,占23.51%~44.44%,兩者占全量的75.34%~84.63%,少量分布在F2和F3中,與對照相比,Pb在F4組分中明顯上升,而在F1組分中呈現逐步降低的趨勢;紫莖澤蘭莖中Pb主要富集于F4和F1,占36.10%~57.14%和20.07%~36.52%,少量富集于F2和F3中,隨Pb濃度增加,Pb在細胞壁和可溶性部分分配比例均呈先增后降的趨勢,而細胞核和葉綠體的分配比例則先減少后增加。根系中細胞亞細胞組分Pb含量依次為F1>F4>F3>F2,大部分Pb位于細胞壁和可溶性部分,分別占39.2%~49.78%和28.27%~37.62%。隨Pb濃度增加,Pb在細胞壁分配比例逐漸下降,而可溶性部分則逐漸增加。以上結果證實了Pb在紫莖澤蘭根莖葉中的主要分布位點集中在可溶性部分和細胞壁,少量分布在紫莖澤蘭各組織的細胞核和葉綠體、線粒體等細胞器中,細胞壁對Pb有較強的束縛能力,對其根來說這種束縛作用更為明顯,隨Pb濃度的增加各組織細胞壁對Pb的束縛能力有所降低,并能降低紫莖澤蘭各組織的細胞核、葉綠體和線粒體等細胞器中Pb的分布,有效地保護了紫莖澤蘭正常的生理功能。
2.2 紫莖澤蘭各組織中Pb的化學形態
一般來說,重金屬都以其特定的形態存在于植物的不同組織中,并保持動態平衡,各組織中重金屬特征形態的性質和活性,對重金屬在植物體內的毒性強弱、轉運及累積有明顯影響。各種化學結合形態的Pb在紫莖澤蘭各組織的含量,所占比例如表2所示。不同濃度Pb處理下,葉中不同形態的Pb占的比例均有所不同,除對照外,均以鹽酸提取態FHCl和水提取態Fw為主,兩者占總量的58.74%~73.04%,其次為乙醇提取態FE,醋酸提取態FHAc和氯化鈉提取態FNaCl最少,隨Pb濃度的增加,FHCl有所減低,而FE則明顯升高。在莖中, 除對照外,各處理Pb的提取形態依次為FHAc>FNaCl>FW>FHCl>FE,
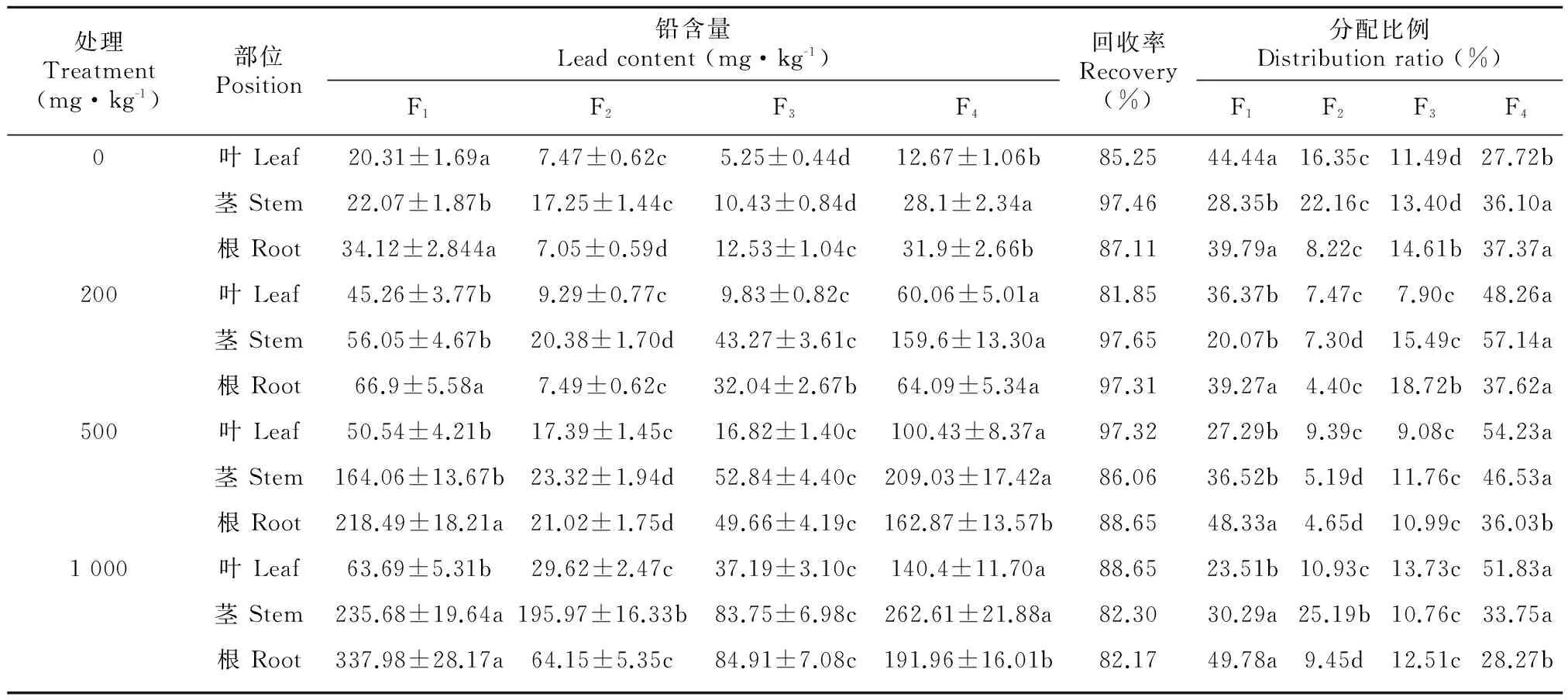
表 1 Pb脅迫下對紫莖澤蘭各組織中Pb亞細胞分布
注: F1. 細胞壁組分; F2. 細胞核和葉綠體組分; F3. 線粒體組分,F4. 可溶組分。表中數據為平均值±標準差(n=3),Pb含量和分配比例中同一行的不同字母表示不同組分有顯著差異(P<0.05)。下同。
Note: F1. Cell wall fraction; F2. Nuclear and chloroplast fraction; F3. Mitochondrial fraction; F4. Soluble fraction. Date are shown as Mean±SD. (n=3). Different letters within the same Pb subcellular fraction of the same lead level indicate significant differences between the four cultivars at 0.05 level. The same below.
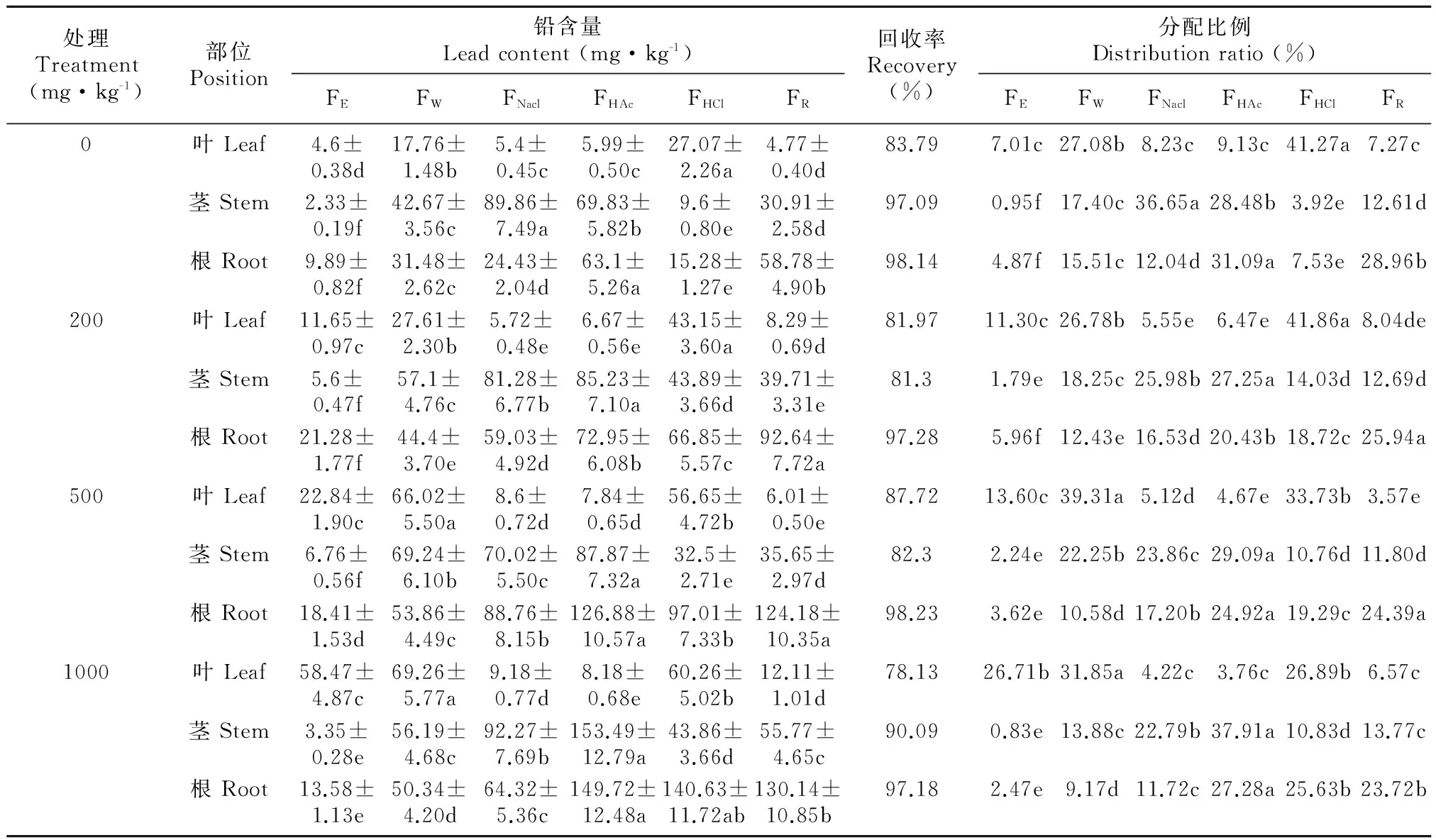
表 2 Pb脅迫下紫莖澤蘭中Pb的化學形態
注: FE. 乙醇提取態; FW. 水提取態; FNaCl. 氯化鈉提取態; FHAc. 醋酸提取態; FHCl. 鹽酸提取態; FR。
Note: FE. ethanol-extractable; Fw. water-extractable; FNacl. Nacl-extractable; FHAc. HAc-extractable; FHCl. HCl-extractable; FR. residual.
FHAc和FNaCl提取態所占比例優勢明顯,且FHAc隨Pb濃度的增加而增加,FNaCl則逐漸減少。Pb在根的主要形態依次為FHAc>FHCl>FNaCl>FW>FE,FHAc和FHCl占總量的39.15%~52.91%,且兩者均隨Pb的增加而分布比例增大。上述結果說明在紫莖澤蘭不同部位Pb存在著不同的化學形態,根莖葉中Pb均與多種配位體結合。
3討論
植物自身進化出了許多機制來適應被重金屬污染土壤的逆境脅迫環境,避免受到重金屬的毒害作用,與其它因素在植株對重金屬的忍耐與解毒過程中發揮了重要作用(楊衛東等, 2009),主要包括選擇性排斥,通過阻止重金屬跨膜運輸來限制重金屬吸收、富集及代謝,通過質子泵作用于重金屬將其泵出胞外;重金屬鰲合,通過鰲合素、金屬硫蛋白、有機酸等贅合重金屬,使其轉化為低毒形態,并通過液泡將其隔離;細胞壁固定,胞外碳水化合物能將重金屬結合并固定于細胞壁上(Zornoza et al, 2002; Ke et al, 2007; Wu et al, 2005; Ramos et al,2002)。
超累積植物自身一般都具備相應的生理解毒機制從而對重金屬產生耐性,使植株能適應高濃度重金屬污染的區域,并保持正常生長,這是因為重金屬在植物體內以贅合物等解毒形態存在,從而失去原有的毒性,形成解毒機制,如與植物細胞壁結合、植物的主動運輸作用將重金屬轉運至液泡、與有機酸或某些蛋白質結合形成絡合物等(夏小燕等, 2007),在植物對重金屬的耐性與富集特性方面,植物細胞壁的固持作用及液泡區隔化效應發揮了重要作用(Kupper et al, 2000);液泡中含有多種能與重金屬離子結合的有機酸、有機堿、蛋白質等,使其在細胞內區隔化;以細胞壁為主體的非原生質體部分以特定的成分及理化性質,構成植物中的“死組織”,其生理代謝活動水平低,這些親金屬離子的配位基團由植物細胞壁中多糖分子(木質素、纖維素、半纖維素、粘膠等)和蛋白質分子中的大量羥基、羧基、氨基、醛基和磷酸基等通過相互作用形成,這些配位集團進入植物體內與重金屬離子發生配位作用,有效防止植物體內重金屬離子發生跨膜轉運,大幅度減少了到達原生質體中重金屬離子的富集量,保護了植物細胞功能區正常生理活性;由此可見,細胞壁是植物體內吸收重金屬代謝解毒的重要場所(Hayens,1980; Allan & Jorrell, 1989)。當細胞壁吸附重金屬離子的量接近飽和時,重金屬離子進入細胞內部通過轉運進入液泡中儲存,液泡中含有多種有機酸、有機堿、蛋白質等,可作用于重金屬使其在植物細胞內被區隔化,降低了原生質中重金屬富集量,從而避免或減輕重金屬對植株的危害。
本研究結果表明紫莖澤蘭中的Pb在葉片分布于可溶性部分和細胞壁中,兩者占全量的75.34%~84.63%;莖類似與葉片也主要分布于可溶性部分和細胞壁中,占總量的36.10%~57.14%和20.07%~36.52%;而根中則富集于細胞壁和可溶性部分,分別占39.2%~49.78%和28.27%~37.62%,其他細胞器中的Pb含量均較少。He et al(2002)認為超累積植物東南景天Pb的亞細胞分布主要集中在細胞壁上,其次是可溶性部分;周小勇等(2008)研究表明長柔毛委陵菜中47%~77%的Pb分布于細胞壁上,其次分布于可溶性部分;江行玉和趙可夫(2002)證實蘆葦細胞中不同位置Pb的含量有明顯差異,Pb在細胞內的具體分布為細胞間隙>細胞壁>液泡>細胞質;本研究結果與上述結果類似,紫莖澤蘭吸收的Pb首先與細胞壁中眾多的化合物形成穩定的絡合物或不溶性有機物而固定下來,若細胞壁上可承載的Pb離子接近飽和狀態后,多余的Pb會通過細胞膜進入原生質中,與液泡中一些可溶性部分如有機酸、有機堿等物質結合而Pb在液泡被區隔化(周小勇等, 2008),使Pb對其它細胞器的危害大大減弱,使紫莖澤蘭對重金屬Pb的耐性相對于一般植物明顯增強。因此,大量的Pb在紫莖澤蘭植株內可溶性部分被區隔化和細胞壁沉淀可能是其耐受高濃度Pb的重要機制,這說明了紫莖澤蘭對于Pb污染土壤有良好的生態修復價值。
植物體內重金屬的遷移能力、活性、毒性和基質分離的難易與其在植物體內呈現的化學形態密切相關(Qiao & Ho, 1996)。重金屬區被植物體內室化的方式一般是通過與植物體內存在的配體結合,這些有機配體主要有3種類型:以檸檬酸、蘋果酸、草酸等為代表的含氧羧酸;以金屬硫蛋白與鰲合素等為代表的含硫基化合物;以組氨酸等為代表的含氮氨基酸。Wu et al(2003)和楊居榮等和黃翌認為由于在不同提取劑中,重金屬的溶解度有很大差異,故Pb、Cd等重金屬存在的化學形態可通過不同溶劑對應的提取量來反映。其中,以氯化物、硝酸鹽為主的重金屬結合態可用乙醇提取;去離子水對以水溶性有機酸鹽、重金屬的一代磷酸鹽M(H2PO4)等方式結合的重金屬有較好的提取效果;氯化鈉能提取與蛋白質結合態或呈吸著態的重金屬和在植物體內以果膠酸鹽的形式結合的重金屬等;而醋酸常用來提取包括M2HPO4和正磷酸鹽M3PO4在內的難溶于H2O的重金屬磷酸鹽結合態;鹽酸一般用來提取草酸鹽等。提取劑的極性強弱對應提取的重金屬的活性、毒性及遷移能力差異顯著,極性越強,提取得到的重金屬鹽在植物體內的活性、毒性及遷移能力越低,以此來分離得到與植物基質結合程度各異的重金屬離子(許嘉琳和宋文昌, 1991)。
長柔毛委陵菜中Pb以鹽酸提取態為主,占總量的28%~60%(周小勇等, 2008);藥用植物麥冬、北沙參和地黃中的Pb的形態是以鹽酸提取態和醋酸提取態形式存在,占藥用部分的65%(劉軍和李先恩, 2002);小麥和水稻葉中的Pb都以鹽酸提取態為主,根中都以醋酸提取態和鹽酸提取態為主(許嘉琳和宋文昌, 1991);蘆葦體內的Pb也以鹽酸提取態和醋酸提取態為主(江行玉和趙可夫, 2002);本研究結果顯示紫莖澤蘭葉中Pb以鹽酸提取態和水提取態為主,兩者占總量的58.74%~73.04%,在莖中,以醋酸提取態和氯化鈉提取態為主,在根中,醋酸提取態和鹽酸提取態占優勢,兩者占總量的39.15%~52.91%;說明紫莖澤蘭葉片中的Pb的存在方式主要以水溶性的有機酸鹽、磷酸鹽為主;紫莖澤蘭莖中的Pb主要以難溶于水的重磷酸鹽、果膠酸鹽方式被吸附或與蛋白質結合的形態存在;根中Pb的存在方式也主要是以難溶于水的重磷酸鹽醋酸、草酸鹽的形式等,表明紫莖澤蘭體內的Pb存在多種配位體,且螯合的鉛都不易被植物吸收,紫莖澤蘭體內遷移能力強的乙醇提取態的比例較低,Pb在其體內的移動性較差,這種Pb形態可明顯促進紫莖澤蘭體內的Pb在細胞壁的固定和液泡的區隔化,使得Pb在紫莖澤蘭植株體內轉運困難,從而避免了Pb對紫莖澤蘭造成損傷。
4結論
本研究證實了紫莖澤蘭中的Pb在莖、葉片中的主要分布區域位于可溶性部分和細胞壁中,而在根中則富集于紫莖澤蘭細胞壁和其它可溶性部分,其它細胞器中的Pb含量均較少;Pb對其它細胞器的毒性由于這種特殊亞細胞分布大大降低,使得紫莖澤蘭對Pb的耐性明顯提高。紫莖澤蘭體內存在多種Pb配位形式,且螯合的鉛大都不易被植物體吸收,紫莖澤蘭體內遷移能力強的乙醇提取態的比例較低,Pb在其體內的移動性較差,這種Pb形態十分有利于細胞壁對Pb的固定和液泡對Pb的區隔化,限制了Pb在紫莖澤蘭植物體內轉運,從而避免了Pb對紫莖澤蘭植株造成損傷。紫莖澤蘭能有效吸收土壤中的Pb,且能適應高濃度Pb處理的土壤,體現了一定的超富集植物特性。可見紫莖澤蘭作為環境污染修復植物,在Pb及其它重金屬污染的土地區的防治上有著良好的應用前景。
參考文獻:
ALLAN DL, JARRELL WM, 1989. Proton and copper adsorption to maize and soybean root cell walls [J]. Plant Physiol, 89(3): 823-832.
GARBISU C, ALKORTA I, 2001. Phytoextraction:a cost-effective plant-based technology for the removal of metals from the environment [J]. Bioreso Technol, 77(3): 229-236.
Gabbrielli R,Panddfini T,Vergnano O,et a1, 1990. Comparison of two ser-pentine species with different nickel tolerance strategies [J]. Plant Soil,22:271-277.
HALL JL, 2002. Cellular mechanisms for heavy metal detoxification and tolerance [J]. J Exper Bot, 53(366): 1-11.
HAYENS RJ, 1980. Ione xehange ProPerties of roots and ionie interactions within the apoplasm: Their role in ion accumulation by Plants [J]. Bot Rev, 46: 75-99.
HE B, YANG XE, NI WZ, et al, 2002. Sedum alfredii: a new lead-accumulating ecotype [J]. Acta Bot Sin Chin Ed, 44(11): 1 365-1 370.
JIA HJ,LI XK,TANG SC,et al, 2009. Allelopathic effects ofEupatoriumadebophorumon seed germination of three woody plants in karst region [J]. Guihaia,29(5): 631-634. [賈海江,李先琨,唐賽春,等, 2009. 紫莖澤蘭對三種巖溶地區木本植物種子萌發的化感作用 [J]. 廣西植物,29(5): 631-634.]
JIANG YX,ZHAO KF, 2002. The Distribution of Pb and Pb-stressed relative proteins in Pb-polluted phragmites austral is seedlings [J]. J Plant Physiol Mol Biol,28(3): 169-174. [江行玉,趙可夫, 2002. 鉛污染下蘆葦體內鉛的分布和鉛脅迫相關蛋白 [J]. 植物生理與分子生物學學報,28(3): 169-174.]KE W, XIONG Z, XIE M, et al, 2007. Accumulation, subcellular localization and ecophysiological responses to copper stress in twoDaucuscarotaL. populations [J]. Plant & Soil, 292(1): 291-304.
KUPPER H, JIEZHAO F, MCGRATH SP, 1999. Cellular compartmentation of zinc in leaves of the hyperaccumulatorThlaspicaerulescens[J]. Plant Physiol, 119(1): 305-312.
KUPPER H, LOMBI E, ZHAO FJ, et al, 2000. Cellular compartmentation of cadmium and zinc in relation to other elements in the hyperaccumulatorArabidopsishalleri[J]. Planta, 212(1): 75-84.
LI B,ZHANG CH, 2008. Studies on the bryophytes and analysis of ecological restore potential in the Lannigou Gold Deposit, Guizhou Province [J]. J Trop & Subtrop Bot,16(6): 511-515. [李冰,張朝暉, 2008. 貴州爛泥溝金礦區苔蘚植物及其生態修復潛力分析 [J]. 熱帶亞熱帶植物學報,16(6): 511-515.]
LI XY,YING H, 2009. The harms prevention and comprehensive utilization ofEupatoriumadenophorum[J]. Biomass Chem Engin,43(1): 57-60. [李雪瑤,應浩, 2009. 紫莖澤蘭的危害、防除及綜合利用 [J]. 生物質化學工程,43(1): 57-60.]
LIU J,LI XN, 2002. Chemical form and distribution of lead in medical plants [J]. Agro-environm Protect,21(2): 143-145. [劉軍,李先恩, 2002. 藥用植物中鉛的形態和分布研究 [J]. 農業環境保護,21(2): 143-145.]LIU XM,WANG QH, 2001. The latest development about phytoremediation of heavy metal contaminated soil [J]. J Gansu Agric Univ,36(1): 8-13. [劉秀梅,王慶仁, 2001. 植物修復重金屬污染土壤的研究進展 [J]. 甘肅農業大學學報,36(1): 8-13.]
LU P,SANG WG,MA KP, 2005. Progress and prospects in research of an exotic invasive species,Eupatoriumadenophorum[J]. Acta Phytoecol Sin,29(6): 1 029-1 037. [魯平,桑衛國,馬克平, 2005. 外來入侵種紫莖澤蘭研究進展與展望 [J]. 植物生態學報,29(6): 1 029-1 037.]NIU YF,FENG YL,XIE JL,et al, 2011. Effects of disturbance intensity on seed germination, seedling establishment and growth ofAgeratinaadenophora[J]. Guihaia,31(6): 795-800. [牛燕芬,馮玉龍,謝建磊,等, 2011. 干擾強度對群落中紫莖澤蘭種子萌發、幼苗定居和生長的影響 [J]. 廣西植物,31(6): 795-800. ]
QIAO L, HO G, 1996. The effect of clay amendment on speciation of heavy metals in sewage sludge [J]. Water Sci & Technol, 34(7-8): 413-420.
RAMOS I, ESTEBAN E, LUCENA J, et al, 2002. Cadmium uptake and subcellular distribution in plants ofLactucasp. Cd-Mn interaction [J]. Plant Sci, 162(5): 761-767.
SALT DE, BLAYLOCK M, KUMAR NPBA, et al, 1995. Phytoremediation: a novel strategy for the removal of toxic metals from the environment using plants [J]. Nature, 13(5): 468-474.
WANG WY,ZHANG CH, 2008. Study on the bryophytes from Shuiyindong carlin gold deposit in Guizhou [J]. Gold,29(6): 8-12. [汪文云,張朝暉, 2008. 貴州水銀洞卡林型金礦苔蘚植物研究 [J]. 黃金,29(6): 8-12.]
Weigel HJ,Jager HJ, 1980. Subcellular distribution and chemical form of cadmium in bean [J]. Plant Physiol, 65:480-482.
WU F, DONG J, QIAN QQ, et al, 2005. Subcellular distribution and chemical form of Cd and Cd-Zn interaction in different barley genotypes [J]. Chemosphere, 60(10): 1 437-1 446.WU FB, ZHANG GP, DOMINY P, 2003. Four barley genotypes respond differently to cadmium:lipid peroxidation and activities of antioxidant capacity [J]. Environ & Exp Bot, 50(1): 67-78.
XIA XY,YANG LQ,ZHAI FQ,et al, 2007. Organic acids alleviating the toxicity of Cd to young wheat plants [J]. J Agro-Environ Sci,26(3): 990-995. [夏小燕,楊麗琴,翟福勤,等, 2007. 有機酸對小麥幼苗鎘毒的緩解作用 [J]. 農業環境科學學報,26(3): 990-995.]
XU JL,SONG WC, 1991. Chemieal forms of Pb, Cd and Cu in crops [J]. Chin J Appl Ecol,2(3): 244-248. [許嘉琳,宋文昌, 1991. 農作物體內鉛, 鎘, 銅的化學形態研究 [J]. 應用生態學報,2(3): 244-248.]YANG JR,HUANG Y, 1994. Mechanism of heavy metal tolerance of plants [J]. Chin J Ecol,13(6): 20-26. [楊居榮,黃翌, 1994. 植物對重金屬的耐性機理 [J]. 生態學雜志,13(6): 20-26.]
YANG WD,CHEN YT,QU MH,et al, 2009. Subcellular distribution and chemical forms of cadmium inSalixmatsudana[J]. Acta Bot Boreal-Occident Sin,(7): 1 394-1 399. [楊衛東,陳益泰,屈明華,等, 2009. 鎘在旱柳中亞細胞分布及存在的化學形態 [J]. 西北植物學報,(7): 1 394-1 399.]ZHOU XY,CHOU RL,LI QF,et al, 2008. Effects of zinc on distribution and chemical for m of lead inPotentillagriffithiivar.velutina[J]. Acta Sci Circumst,28(10): 2 064-2 071. [周小勇,仇榮亮,李清飛,等, 2008. 鋅對長柔毛委陵菜中鉛的分布和化學形態的影響 [J]. 環境科學學報,28(10): 2 064-2 071.]
ZORNOZA P, VáZQUEZ S, ESTEBAN E, et al, 2002. Cadmium-stress in nodulated white lupin: strategies to avoid toxicity [J]. Plant Physiol Biochem, 40(12): 1 003-1 009.
ZU YQ, LI Y, CHEN JJ, et al, 2005. Hyperaccumulation of Pb,Zn and Cd in herbaceous grown on lead-zinc mining area in Yunnan, China [J]. Environm Inter, 31(5): 755-762.
Subcellular distribution and chemical forms of lead inEupatoriumadenophorumat different lead levels
LIU Xiao-Wen1,2,3, QI Cheng-Mei1, LI Yuan2,3, YAN Dong-Dong2,3,WANG Qiu-Xia2,3, GUO Mei-Xia2,3, CAO Ao-Cheng2,3*
( 1.CollegeofBiologicalandChemicalEngineering,HunanUniversityofScienceandTechnology, Yongzhou 425199, China;2.StateKeyLaboratoryforBiologyofDiseasesandInsertPests, Beijing 100193, China; 3.InstituteofPlantProtection,ChineseAcademyofAgriculturalSciences, Beijing 100193, China )
Abstract:The differential centrifugation technique and sequential chemical extraction method were used to study the subcellular distribution and chemical forms of lead in the roots, leaves and stems of Eupatorium adenophorum. The results showed that with the increasing of lead levels, the contents of lead in subcellular distribution of leaves, stems and roots significant increased. The lead distributed mainly in the soluble partition and cell wall in leaves, occupied 75.34%-84.63% of the total. Lead distributed in stems were similar to it in leaves. It mainly accumulated in the soluble partition and cell wall, which occupied 36.10%-57.14% and 20.07%-36.52%, respectively. And it distributed mainly in cell wall and soluble partition in roots, which occupied 39.20%-49.78% and 28.27%-37.62%, respectively. There was less lead in other organelles. The chemical forms of lead in leaves of E. adenophorum were mainly in HCl and water, occupied 58.74%-73.04% of the total. In stems, the main chemical forms were HAc and NaCl, and the chemical forms of HAc and HCl were predominant in roots, occupied 39.15%-52.91% of the total.
Key words:lead, Eupatorium adenophorum, subcellular distribution, chemical forms
DOI:10.11931/guihaia.gxzw201404010
收稿日期:2014-07-18修回日期: 2014-12-18
基金項目:國家公益性行業(農業)科技專項(201103027); 湖南省教育優秀青年項目(14B071);中國博士后科學基金第56批面上項目(2014M561110);湖南省自然科學基金(13JJ6080);湖南省重點學科建設項目(2011-76);湖南省高校科技創新團隊支持計劃項目(2012-318)[Supported by National Public Welfare Fund for Agricultural Research(201103027); Preject for Advanced Youth of Hunan Provincial Education Department(14B071); Postdoctoral Science Foundation of China(2014M561110); Key Discipline Construction Project of Hunan Province(2011-76); Science and Technology Innovation Team Plan of Hunan Provincial Colleges and Universities(2012-318)]。
作者簡介:劉小文(1983-),男,湖南耒陽人,博士,副教授,研究方向為生物安全和環境毒理學,(E-mail)lxw1110@126.com。
*通訊作者:曹坳程,博士,研究員,研究方向為生物入侵和土壤消毒,(E-mail)caoac@vip.sina.com。
中圖分類號:Q945.79
文獻標識碼:A
文章編號:1000-3142(2016)03-0335-07
劉小文,齊成媚,李園,等. 不同鉛水平下紫莖澤蘭細胞內鉛的分布和化學形態的分析 [J]. 廣西植物, 2016, 36(3):335-341
LIU XW, QI CM, LI Y, et al. Subcellular distribution and chemical forms of lead inEupatoriumadenophorumat different lead levels [J]. Guihaia, 2016,
36(3):335-341
