CYP2C8及CYP3A4細胞表達體系的構建及其在小分子激酶藥物對紫衫醇代謝途徑抑制研究中的應用
2016-06-23 13:49:52李遠黃潔瓊李佳俊汪維鵬張洪建
生物技術通報 2016年7期
李遠 黃潔瓊 李佳俊 汪維鵬 張洪建
(蘇州大學藥學院,蘇州 215123)
CYP2C8及CYP3A4細胞表達體系的構建及其在小分子激酶藥物對紫衫醇代謝途徑抑制研究中的應用
李遠 黃潔瓊 李佳俊 汪維鵬 張洪建
(蘇州大學藥學院,蘇州 215123)
構建CYP2C8及其3種突變體細胞表達體系,以紫杉醇為底物研究CYP2C8基因多態性對其酶活性的影響,以及構建CYP2C8和CYP3A4共轉染細胞體系研究小分子激酶抑制劑對紫杉醇代謝途徑的抑制。根據基因文庫分別合成CYP3A4以及CYP2C8及其3種突變體CYP2C8*2(805A>T)、CYP2C8*3(416G>A,1196A>G)、CYP2C8*4(792C>G)的基因編碼片段,將其連接到PEGFP-N1表達質粒,測序驗證。將CYP2C8野生型及其突變體分別轉染HepG2細胞,24 h后加入紫杉醇進行孵育,通過建立好的LC-MS/MS方法對代謝物進行定量分析。同時,也將野生型CYP2C8和CYP3A4質粒按一定的濃度比轉入HepG2細胞構建共表達體系。并篩選出合適的質粒濃度比轉染細胞,在加入紫杉醇孵育時,同時加入小分子激酶抑制劑,考察小分子激酶抑制劑對紫杉醇代謝途徑的抑制作用。結果表明,CYP2C8*4代謝酶對紫杉醇的代謝能力存在明顯差異,其中CYP2C8 *2和CYP2C8 *3代謝活性分別是野生型的81%(P<0.05)和87%(P<0.05),而CYP2C8*4則是野生型的65%(P<0.01)。 尼洛替尼完全抑制了紫杉醇的代謝,阿法替尼對紫杉醇的兩條代謝途徑抑制達30%,而伊馬替尼選擇性抑制了CYPD3A4的活性。不同基因型CYP2C8對紫杉醇的代謝存在差異,可能是導致臨床療效不同的原因。小分子激酶抑制劑在與紫杉醇聯合使用時,對紫杉醇代謝的抑制各不相同。
紫杉醇;代謝酶;基因多態性;小分子激酶抑制劑;藥物相互作用
CYP2C8是CYP2C家族中最晚發現的一個成員,大約占CYP2C家族的26%。 CYP2C8主要分布在肝臟,占肝CYP代謝酶總量的7%;其在肝臟外的組織中也存在少量的分布[1]。CYP2C8參與催化大約5%的臨床藥物,并且存在著明顯的基因多態性,對藥物的代謝清除有著明顯的影響[2]。常見的CYP2C8基因多態性包括CYP2C8*2(805A>T)、*3(416G>A,1196A>G)、*4(792C>G),其中CYP2C8*2只在非洲人中被發現,等位基因頻率為19%,CYP2C8*3和CYP2C8*4只在白種人中被發現,等位基因頻率分別為23.5%和5.8%,其他CYP2C8基因多態的頻率較低[3]。目前已經知道CYP2C8的不同基因型對抗糖尿病藥物[4],抗腫瘤藥物[5],非甾體抗炎藥[6]等藥物的代謝過程有著重要的影響。
紫杉醇是一種抗微管細胞毒素藥物,它通過促進微管蛋白聚合、抑制解聚、保持微管蛋白穩定、抑制細胞有絲分裂和促進腫瘤細胞凋亡發揮抗腫瘤作用。紫杉醇具有較廣的抗癌譜,近年來被廣泛應用于卵巢癌、肺癌、乳腺癌、胃癌、頭頸部癌等惡性腫瘤[7-9]。紫杉醇在人體內能夠被CYP3A4和CYP2C8代謝,分別生成p-3'-羥基紫杉醇和6α-羥基紫杉醇[10,11]。在人肝微粒體中,6α-羥基紫杉醇的生成量高于p-3'-羥基紫杉醇,表明CYP2C8在紫杉醇體內代謝清除過程中扮演著重要角色[12]。為了達到更好的治療效果,紫杉醇經常與一些小分子激酶抑制劑等新型抗腫瘤藥物聯合使用,以使抗腫瘤功效最大化。雖然聯合療法已較多報道,但是紫杉醇在與這些小分子激酶抑制劑聯用時,藥物之間的相互作用目前還鮮有報道。因而有必要研究清楚各種小分子激酶抑制劑及藥物對酶的抑制作用。
在不同的病人體內,紫杉醇的藥代動力學的變化各不相同,這種變化與CYP3A4和CYP2C8表達及酶活性的不同有著緊密的聯系。近年來也進行了大量的實驗研究CYP酶的多態性,以及它們與個體化差異的關系在紫杉醇的藥效和毒性中起到的作用[13]。例如,CYP2C8*3與高效的臨床反應以及神經毒性風險的提高有關[14-16]。CYP3A4*22基因型能夠影響有紫杉醇引起的神經毒性[17]。這 些都表明不同基因型的CYP酶對紫杉醇的代謝情況存在差異,這些差異將會影響到紫杉醇的抗癌作用和毒副作用。而本課題將研究紫杉醇在不同基因型的CYP2C8代謝酶中的代謝情況。
為了更好地研究紫杉醇與CYP2C8、CYP3A4代謝酶以及小分子激酶抑制劑的關系,需要構建一個合適的系統。目前常用的系統有細菌表達系統[18]、酵母表達系統[19]、昆蟲表達系統[20]和哺乳動物表達系統[21]。P450酶在不同的表達系統中有著各自的表達特點。最終,本研究選擇了哺乳動物表達系統——HepG2細胞株。HepG2細胞株與其它主要的人肝細胞相比,更加的經濟和使用方便。由于它來源于原發性肝癌的活體組織,保留了許多外源性物質的P450代謝活性[22],但CYP酶活性較低[23],更為重要的是HepG2細胞內含有P450酶催化活性所必需的CYP450 氧化還原酶和細胞色素b5等輔助因子[24]。因此它是用來研究CYP450介導藥物代謝的一個理想的系統[25]。通過P450酶表達系統,可以避免體內藥物研究存在的安全性低,通量低,易受外界條件影響等缺點[26],從而能夠更加準確的研究藥物之間相互作用以及不同的基因型對藥物的代謝的影響,為臨床用藥安全提供更多的參考和更好的保障。本研究利用生物合成學的方法,構建CYP2C8及其3種突變體細胞表達體系,以紫杉醇為底物研究CYP2C8基因多態性對其酶活性的影響,以及構建CYP2C8和CYP3A4共轉染細胞體系研究小分子激酶抑制劑對紫杉醇代謝途徑的抑制,以期為臨床給藥提供參考依據。
1 材料與方法
1.1 材料
HepG2細胞購自上海生物化學與細胞生物學研究所;FBS(胎牛血清)、DMEM高糖培養基、RPMI-1640培養基、鏈霉素、青霉素以及PBS緩沖液購自Hyclone公司;胰蛋白酶購自Gibco公司;脂質體2000試劑購自Invitrogen公司;甲醇、乙腈購自Merck公司;紫杉醇、6α-羥基紫杉醇、p-3'-羥基紫杉醇以及多西紫杉醇購自美國Sigma公司;阿法替尼、伊馬替尼、尼洛替尼購自上海ChemBest公司。
1.2 方法
1.2.1 細胞培養 采用含10% FBS和1%雙抗(青霉素及鏈霉素)的DMEM高糖培養基進行培養,培養條件為37℃、5% CO2、飽和濕度。
1.2.2 質粒構建 根據NCBI GENE(http://www.ncbi. nlm.nih.gov/gene/)文庫,利用合成生物學方法合成CYP2C8和CYP3A4基因,與載體pEGFP-N1進行組裝、測序,以上步驟由金唯智公司(http://www. genewiz.com.cn/)完成。測序正確后進行功能驗證。
1.2.3 細胞轉染 參考產品說明書,將一定濃度的質粒稀釋至50 μL不含血清和抗生素的RPMI-1640培養基中,2 μL脂質體2000懸液加入50 μL不含血清和抗生素的RPMI-1640培養基中,在室溫孵育5 min。然后將含質粒的RPMI-1640培養基和含脂質體2000的RPMI-1640培養基混合在一起,并充分混勻,靜置20 min,使得含質粒的載體和脂質體能夠充分的混合。同時準備好只加入了脂質體2000的RPMI-1640培養基,作陰性對照;吸去600 μL接種到24孔板中的培養液,每孔加入100 μL上述混合物,陰性對照孔加入400 μL新鮮培養基 及100 μL只加入了脂質體2000的RPMI-1640培養基。經4 h后,吸棄24孔板中的轉染培養液,同時向各孔加入1 mL的含有血清不含抗生素的DMEM培養基,繼續培養。
1.2.4 活性測試 細胞轉染48 h之后,用KHB緩沖液清洗細胞兩次。用KHB緩沖液配制濃度為5 μmol/L的紫杉醇作為探針底物,每孔500 μL進行孵育。4 h后,取出24孔板,每孔吸取100 μL KHB孵育液,加入到含有內標的100 μL有機溶劑中。振蕩2 min后,1.3×104r/min 4℃離心10 min,吸取上層清液進行LC-MS/MS分析。具體過程見圖1。
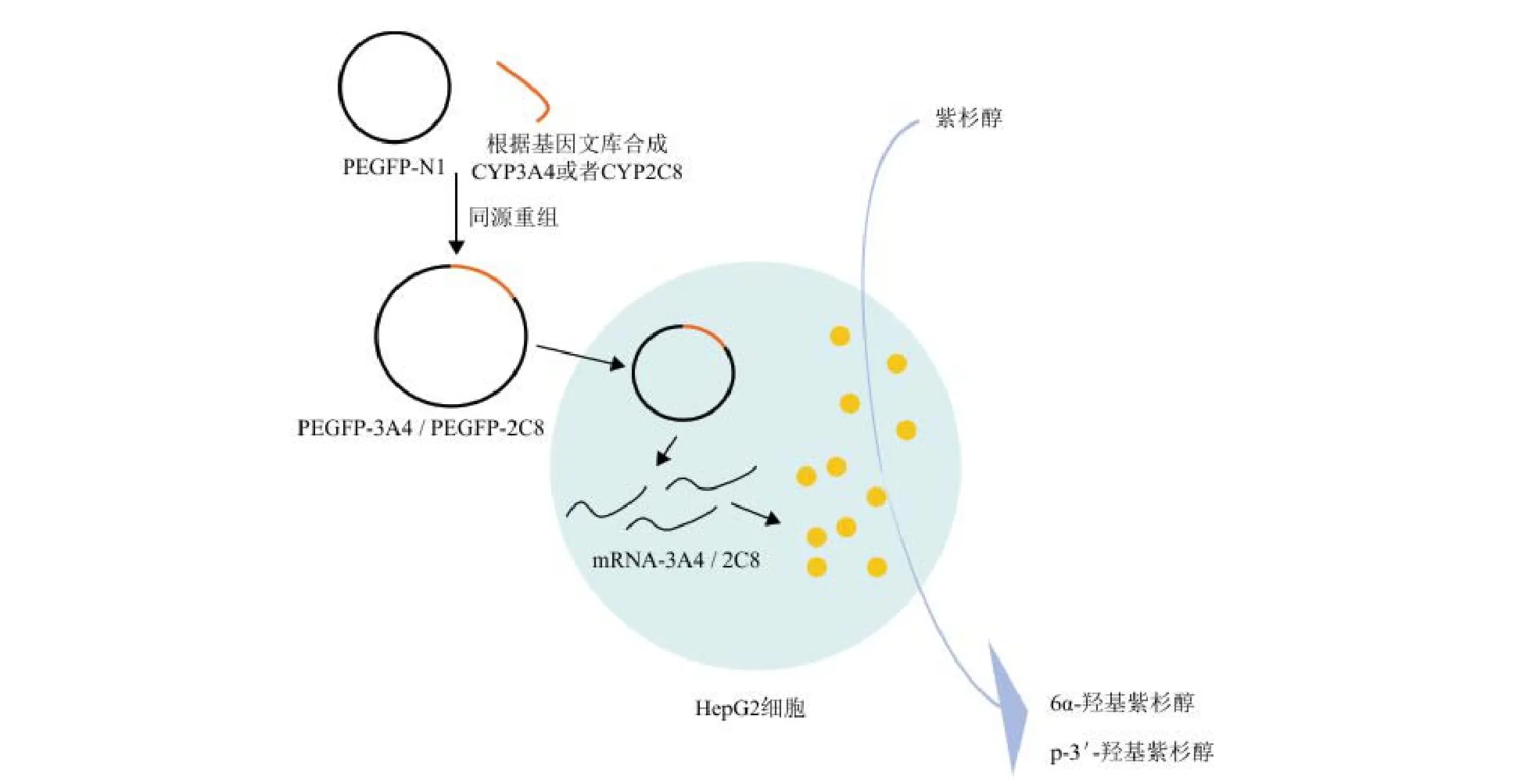
圖1 合成CYP2C8和CYP3A4基因,與載體同源重組以及功能測試
1.2.5 抑制試驗 細胞鋪板24 h后進行細胞轉染。細胞轉染48 h后進行活性測試實驗,在活性測試實驗中,用KHB緩沖液將各個小分子激酶抑制劑配制到臨床使用濃度,并于活性測試時與底物同時加入。將每孔細胞進行裂解,測定蛋白濃度,并利用蛋白濃度來對活性測試結果進行校準。
1.2.6 LC-MS/MS分 析 CYP2C8和CYP3A4的 代謝產物通過LC-MS/MS系統 來進行定量分析。美國Applied Biosystems公司API 4000 Q Trap三重四極桿串聯質譜儀,配有電噴霧離子 源(Turbo I onS prav),高效液相分離包括LC-20AD泵,CBM-20A控制器,SIL-20A自動進樣器,Shimadzu CTO-20A柱溫箱。電離源為ESI源,正離子方式檢測,噴霧電壓為5.5 kV,霧化溫度為500℃,氣簾氣CUR為20 L/min,輔助氣AUX(GAS2)為65 L/min。掃描方式為多反應監測(MRM)。采用Agela Venusil XBP C18 色譜柱(50×2.1 mm;5 μm)實現代謝物分離,柱溫40℃,流速為0.3 mL/min,流動相為A相(水∶乙腈=95∶5)與B相(水∶乙腈=5∶95)梯度混合。
2 結果
2.1 LC-MS/MS檢測方法
建立紫杉醇代謝物(6 α-羥基紫杉醇和p-3'-羥基紫杉醇)專屬靈敏的LC-MS/MS方法,進行定量檢測。采用正離子模式,選擇離子對m/z 892.4→324.1、m/z 892.3→308.1和m/z 830.3→549.4分別測定p-3'-羥基紫杉醇、6α-羥基紫杉醇和多西紫杉醇(內標)的含量;色譜圖如圖2所示。代謝物與內標實現基線分離,峰形良好,無干擾。
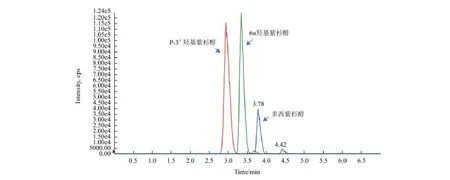
圖2 紫杉醇代謝物LC-MS/MS色譜圖
2.2 CYP2C8的基因多態性對紫杉醇代謝的影響
為了研究CYP2C8的不同基因型對紫杉醇代謝的影響,分別構建了CYP2C8野生型質粒(WT)和3種突變型質粒,包括*2(805A>T)、*3(416G>A,1196A>G)和*4(792C>G)。分別將其轉入HepG2細胞后進行功能測試,結果如圖3所示。轉染了CYP2C8野生型及不同基因型質粒后,6α-羥基紫杉醇的生成量顯著增加,且在不同的CYP2C8基因型細胞之間,6α-羥基紫杉醇的生成量存在差異。其中CYP2C8 *2和*3代謝活性分別是野生型的81%(P<0.05)和87%(P<0.05),而*4則僅為野生型的65%(P<0.01)。
2.3 CYP3A4和CYP2C8共表達體系的構建
紫杉醇能夠同時被CYP2C8和CYP3A4代謝生成6α- 羥基紫杉醇和p-3'-羥基紫杉醇。HepG2細胞中同時轉入不同比例的CYP2C8和CYP3A4后的功能測試結果如圖4所示。轉染了CYP2C8和CYP3A4的細胞中的代謝產物生成顯著增加,并且隨著CYP2C8比例的提高,6α-羥基紫杉醇生成量與p-3'-羥基紫杉醇生成量之比也隨之升高。同時,當CYP3A4與CYP2C8質粒濃度比為1∶5時,CYP3A4和CYP2C8參與紫杉醇代謝比例(活性比)與體外人肝微粒體結果相似[13],因此用該轉染比例進行后續的抑制試驗。
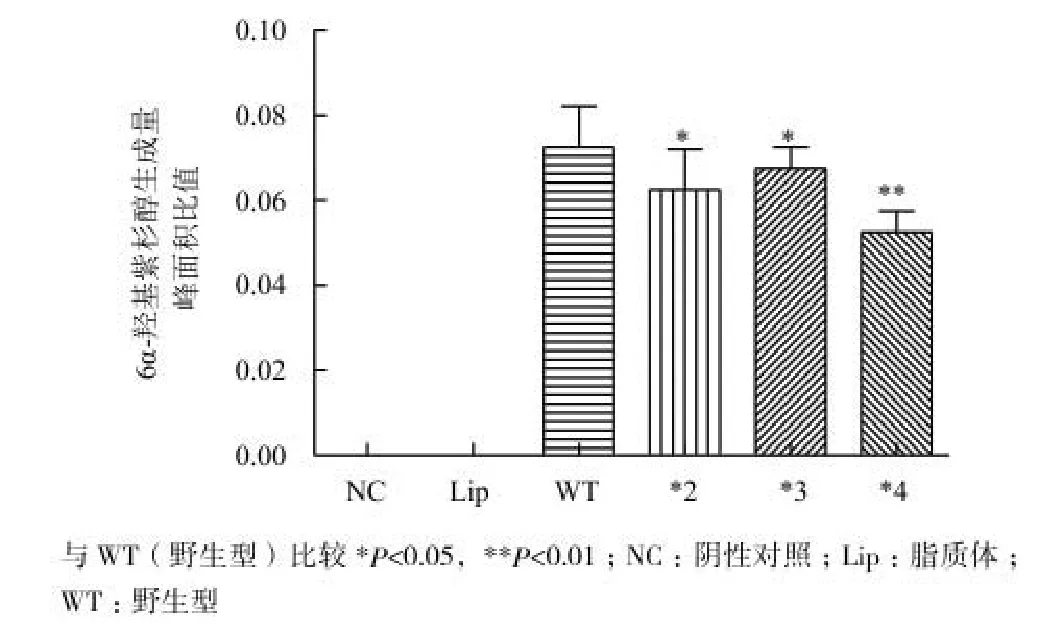
圖3 轉染CYP2C8不同基因型質粒的HepG2細胞的6α-羥基紫杉醇生成量
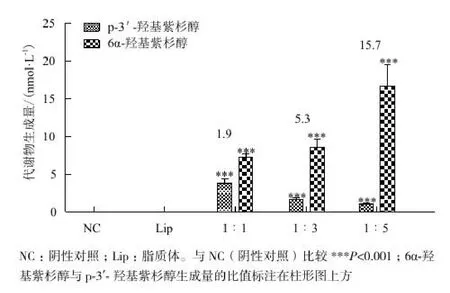
圖4 不同濃度比的CYP3A4和CYP2C8質粒,6α-羥基紫杉醇和p-3'-羥基紫杉醇的生成量
2.4 小 分子激酶抑制劑對紫杉醇代謝途徑的抑制
CYP3A4和CYP2C8質粒濃度比為1∶5時(對照組),小分子激酶抑制劑阿法替尼、伊馬替尼和尼洛替尼對紫杉醇代謝途徑的抑制結果如圖5所示。結果顯示,尼洛替尼完全抑制了紫杉醇的代謝,阿法替尼對紫杉醇的兩條代謝途徑抑制達30%,而伊馬替尼選擇性抑制了CYP3A4的活性。
3 討論
體外藥物代謝研究通常以肝微粒體、胞漿、S9或原代肝細胞等為主要體系。較之肝微粒體,原代肝細胞存在著來源、質量、較大的個體差異等不利因素。因此,構建穩定可控的體外細胞代謝體系,不僅可以避免原代肝細胞的各種缺陷,同時也會較大提高研究及藥物篩選的效率。合成生物學具有簡單準確等優勢,可以將目標基因(如CYP同工酶)構建成相對應的質粒并轉染至細胞中,進行表達,獲得單一的高表達的CYP酶或者按照一定比例的不同的CYP酶的共表達體系,從而能夠更加準確、便捷地進行藥物代謝研究。
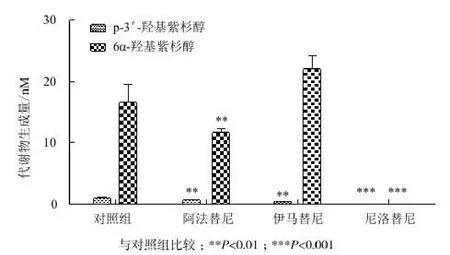
圖5 阿法替尼、伊馬替尼、尼洛替尼對紫杉醇代謝的抑制
代謝研究的另一個挑戰是代謝酶的基因多態性。基因多態性是指同一人群的同一基因位點上存在多個等位基因形態,并且遵循孟德爾遺傳定律[26]。由于超過半數以上的藥物都是由CYP450介導代謝的,CYP450的基因多態性已經成為造成藥物反應個體差異的主要原因。本研究以CYP2C8為目標,合成了CYP2C8*1、*2、*3 及*4的關鍵基因,構建了獨立的HepG2表達體系。在研究過程中,我們發現轉染了CYP2C8質粒的細胞的6α-羥基紫杉醇的生成量與空白和只加了脂質體2000的細胞相比具有顯著的增加,同時在轉染了CYP2C8野生型和突變體質粒的細胞中,6α-羥基紫杉醇的生成量存在著差別。這些結果說明了構建的細胞表達體系具有代謝功能,不同基因型CYP2C8對紫杉醇的代謝作用存在著“基因劑量”的影響。
作為臨床上常用的經典抗癌藥物,紫杉醇能夠同時被CYP3A4和CYP2C8代謝。由于捐獻者之間的基因差異,健康狀況和用藥歷史的不同,肝微粒體中P450酶的活性也不相同,紫杉醇的代謝產物6α-羥基紫杉醇和p-3'-羥基紫杉醇生成量的比值也不相同[27]。在我們之前的研究中,6α-羥基紫杉醇的生成量大約是p-3'-羥基紫杉醇的9倍。這說明盡管CYP2C8在肝中的表達量低于CYP3A4,但其在紫杉醇的代謝清除中起主導作用。甚至在有些捐獻者的肝微粒體中,6α-羥基紫杉醇的生成量能達到p-3'-羥基紫杉醇的13倍[12,28]。在膽汁和尿液中,6α-羥基紫杉醇的生成量能達到p-3'-羥基紫杉醇的4倍,紫杉醇引起的神經毒性的程度和CYP2C8的活性以及6α-羥基紫杉醇的生成量一致。這說明P450酶介導的紫杉醇代謝是影響其臨床藥效的主要因素,因此應該重視藥物與藥物之間的相互作用,并尋求基于紫杉醇的聯合療法來達到更好的治療效果。
由于紫杉醇能夠同時被CYP3A4和CYP2C8代謝,所以本研究選擇紫杉醇作為一個合適的探針底物來研究藥物對CYP2C8和CYP450抑制作用。本實驗中,我們成功構建了CYP2C8和CYP3A4的HepG2的雙轉體系,并驗證了其代謝功能,在該HepG2的雙轉細胞中,根據p-3'-羥基紫杉醇和6α-羥基紫杉醇的生成量可以反映CYP3A4和CYP2C8的代謝活性。我們發現當轉入的CYP3A4與CYP2C8質粒比為1∶5時,CYP3A4 和CYP2C8參與紫杉醇代謝比例與體外微粒體實驗結果相似。故選擇1∶5的比例構建的細胞表達系統來模擬肝微粒體體系,研究小分子激酶抑制劑對P450酶的抑制作用,來進一步探究體內藥物與藥物的相互作用。臨床上使用紫杉醇時常與其他抗腫瘤藥物聯合使用,產生了一系列的藥物相互作用,因此進一步的體內研究可集中于以紫杉醇為基礎的更安全有效的聯合療法。而在癌癥治療中,臨床上已有超過10種以上的小分子激酶抑制劑與紫杉醇進行聯用,而與這些小分子激酶抑制劑進行聯用時的相互作用還鮮有報道。本實驗選用了臨床上已使用的3種小分子激酶抑制劑:尼洛替尼、阿法替尼和伊馬替尼進行抑制研究。CYP2C8和CYP3A4酶活性的改變能夠導致代謝產物生成量的改變,這些改變將會影響基于紫杉醇的聯合療法的療效和風險。實驗中我們發現,尼洛替尼完全抑制了紫杉醇代謝途徑,阿法替尼部分抑制了紫杉醇代謝途徑,而伊馬替尼選擇性抑制了CYP3A4的活性,但具體的作用機制還需要進一步的研究。
由于藥物與藥物相互作用以及個體之間的差異,在制定臨床給藥方案時需要考慮的因素也越來越多。我們需要更加著重于藥物之間的相互作用以及個體差異對藥物代謝的影響并以此給臨床用藥提供正確的指導,提高藥效,降低風險。
4 結論
本研究通過運用生物合成技術,成功構建了CYP2C8及其3個變異體以及CYP3A4體外表達體系,以紫杉醇為底物研究了基因多態性以及小分子激酶抑制劑對酶活性的影響,其中CYP2C8 *2、CYP2C8 *3、CYP2C8 *4活性分別為野生型活性的81%、87%和65%;而在抑制實驗中,尼洛替尼完全抑制了紫杉醇的代謝,阿法替尼對紫杉醇的兩條代謝途徑抑制達30%,而伊馬替尼選擇性抑制了CYP3A4的活性。
[1]Klose TS, Blaisdell JA, Goldstein JA. Gene structure of CYP2C8 and extrahepatic distribution of the human CYP2Cs[J]. Journal of Biochemical & Molecular Toxicology, 1999, 13(6):289-295.
[2]Nebot N, Crettol S, D’Esposito F, et al. Participation of CYP2C8 and CYP3A4 in the N-demethylation of imatinib in human hepatic microsomes[J]. Br J Pharmacol, 2010, 161(5):1059-1069.
[3]Daily EB, Aquilante CL. Cytochrome P450 2C8 pharmacogenetics:a review of clinical studies[J]. Pharmacogenomics, 2009, 10(9):1489-1510.
[4]Gertz M, Tsamandouras N, S?ll C, et al. Reduced physiologicallybased pharmacokinetic model of repaglinide:impact of OATP1B1 and CYP2C8 genotype and source of in vitro data on the prediction of drug-drug interaction risk[J]. Pharmaceutical Research, 2014, 31(9):2367-2382.
[5]Khan MS, Barratt DT, Somogyi AA. Impact of CYP2C8*3 polymorphism on in vitro metabolism of imatinib to N-desmethyl imatinib[J]. Xenobiotica, 2016, 46(3):278-287.
[6]Yu L, Shi D, Ma L, et al. Influence of CYP2C8 polymorphisms on the hydroxylation metabolism of paclitaxel, repaglinide and ibuprofen enantiomers in vitro[J]. Biopharmaceutics & Drug Disposition, 2013, 34(5):278-287.
[7]Fountzilas G, Kalofonos HP, Dafni U, et al. Paclitaxel and epirubicin versus paclitaxel and carboplatin as first-line chemotherapy in patients with advanced breast cancer:a phase III study conducted by the Hellenic Cooperative Oncology Group[J]. Annals of Oncol, 2004, 15(10):1517-1526.
[8]Ramalingam SS, Mlfrankel M. Carboplatin and Paclitaxel in combination with either vorinostat or placebo for first-line therapy of advanced non-small-cell lung cancer[J]. J Clin Oncol, 2010, 28(1):56-62.
[9]郭培鳳, 郭云利. 紫杉醇應用相關不良反應觀察[J]. 醫學信息, 2014(2):643.
[10]Monsarrat B, Mariel E, Cros S, et al. Taxol metabolism. Isolation and identification of three major metabolites of taxol in rat bile[J]. Drug Metabolism & Disposition, 1990, 18(6):895-901.
[11]Walle T, Kumar GN, McMillan JM, et al. Taxol metabolism in rat hepatocytes[J]. Biochemcal Pharmacology, 1993, 46(9):1661-1664.
[12]Taniguchi R, Kumai T, Matsumoto N, et al. Utilization of human liver microsomes to explain individual differences in paclitaxel metabolism by CYP2C8 and CYP3A4[J]. Journal of Pharmacological Sciences, 2005, 97(1):83-90.
[13]Wang Y, Wang M, Qi H, et al. Pathway-dependent inhibition of paclitaxel hydroxylation by kinase inhibitors and assessment of drug-drug interaction potentials[J]. Drug Metabolism & Dispos, 2014, 42(4):782-795.
[14]Leskel? S, Jara C, Leandro-García LJ, et al. Polymorphisms in cytochromes P450 2C8 and 3A5 are associated with paclitaxel neurotoxicity[J]. Pharmacogenomics Journal, 2011, 11(2):121-129.
[15]Hertz DL, Motsinger-Reif AA, Amy Drobish, et al. CYP2C8*3 predicts benefit/risk profile in breast cancer patients receiving neoadjuvant paclitaxel[J]. Breast Cancer Research & Treatment, 2012, 134(1):401-410.
[16]Hertz DL, Roy S, Motsinger-Reif AA, et al. CYP2C8*3 increases risk of neuropathy in breast cancer patients treated with paclitaxel[J]. Ann Oncol , 2013, 24(6):1472-1478.
[17]Graan AJMD, Laure E, Sprowl JA, et al. CYP3A4*22 genotype and systemic exposure affect paclitaxel-induced neurotoxicity[J]. Clin Cancer Res, 2013, 19(12):3316-3324.
[18]Behrendorff JB, Moore CD, Kim KH, et al. Directed evolution reveals requisite sequence elements in the functional expression of P450 2F1 in Escherichia coli[J]. Chemical Research in Toxicology, 2012, 25(9):1964-1974.
[19]Imaoka S, Yamada T, Hiroi T, et al. Multiple forms of human P450 expressed in Saccharomyces cerevisiae[J]. Biochemical Pharmacology, 1996, 51(8):1041-1050.
[20]路珂, 曾蘇, 姚形煒. 利用細菌/桿狀病毒系統在昆蟲細胞中表達人CYP2E1[J]. 浙江大學學報:醫學版, 2008, 37(2):118-125.
[21]Gerets HHJ, Tilmant K, Gerin B, et al. Characterization of primary human hepatocytes, HepG2 cells, and HepaRG cells at the mRNA level and CYP activity in response to inducers and their predictivity for the detection of human hepatotoxins[J]. Cell Biology & Toxicology, 2012, 28(2):69-87.
[22]薛正楷. 穩定表達人CYP3A4基因與Bama小型豬CYP3A基因的HepG_2細胞株的建立及探針藥物代謝表征的比較研究[D]. 重慶:重慶醫科大學, 2009.
[23]Westerink WMA, Schoonen WGEJ. Cytochrome P450 enzyme levels in Hepg2 cells and cryopreserved primary human hepatocytes and their induction in Hepg2 cells[J]. Toxicology in Vitro, 2007, 21(8):1581-1591.
[24]謝章明, 陳樞青. 人CYP的異源表達及其在新藥研發早期的作用[J]. 浙江大學學報:醫學版, 2013, 1(1):109-113.
[25]Vignati L, Turlizzi E, Monaci S, et al. An in vitro approach to detect metabolite toxicity due to CYP3A4-dependent bioactivation of xenobiotics[J]. Toxicology, 2005, 216:154-167.
[26]梁艷, 李彥, 白清清. CYP450基因多態性研究進展[J]. 武警醫學, 2014, 25(5):529-532.
[27]Sonnichsen DS, Liu Q, Schuetz EG, et al. Variability in human cytochrome P450 paclitaxel metabolism[J]. J Pharmacol Exp Ther, 1995, 275(2):566-575.
[28]Václavíková R, Horsky S, ?imek P, et al. Paclitaxel metabolism in rat and human liver microsomes is inhibited by phenolic antioxidants[J]. Naunyn-Schmiedeberg’s Archives of Pharmacology, 2003, 368(3):200-209.
(責任編輯 李楠)
The Construction of CYP2C8 and CYP3A4,and the Inhibition of Paclitaxel Metabolism by Small Kinase Inhibitor
LI Yuan HUANG Jie-qiong LI Jia-jun WANG Wei-peng ZHANG Hong-jian
(College of Pharmaceutical Sciences,Soochow University,Suzhou 215123)
The aims of this work are to establish the cell expression system of CYP2C8 and its 3 mutants,to study the effects of gene polymorphisms on its enzymatic activities,and to investigate the effects of small molecular kinase inhibitors on paclitaxel metabolism with the established CYP2C8 and CYP3A4 co-transfected cell lines. The gene fragments of CYP3A4,CYP2C8 and its three mutants(CYP2C8*2(805A>T),CYP2C8*3(416G>A,1196A>G),and CYP2C8*4(792C>G))were synthesized based on gene bank. Then those fragments were ligated to expression plasmid PEGFP-N1 for sequencing evaluation. Twenty-four hours after the wild and mutant plasmids were transfected into HepG2 cells,paclitaxel was added and incubation proceeded,and then the metabolites were quantitatively detected by well-constructed LC-MS/MS. Concurrently,the wild CYP2C8 and CYP3A4 plasmids in certain concentration ratios were transfected into HepG2 cells,to establish a co-expression system. Screening the plasmids with proper concentration ratio and transfecting them to the cells,adding small molecule kinase inhibitor while adding paclitaxel for incubation,the inhibition of paclitaxel metabolism by small molecule kinase inhibitors was investigated. The results showed that the metabolic enzyme of different CYP2C8 caused the significant difference of paclitaxel metabolicactivity,of which the metabolic activity of CYP2C8*2,CYP2C8*3,CYP2C8*4 remained at about 80%(P < 0.05),87%(P < 0.05),and 65%(P < 0.01),respectively,as compared to the wild type. Nilotinib completely inhibited the metabolism of paclitaxel and axitinib showed a 30% inhibition,while imatinib didn’t have any inhibitory effect. As conclusion,different genotypes of CYP2C8 differentially affect the overall paclitaxel metabolism,which might be the reason that results in the varied treatment effect in clinic. Small molecule kinase inhibitors while combined use with paclitaxel may inhibit the metabolism of paclitaxel to different extents.
paclitaxel;metabolism enzyme;gene polymorphism;small molecule kinase inhibitors;drug-drug interaction
10.13560/j.cnki.biotech.bull.1985.2016.07.032
2015-10-27
國家自然科學基金項目(81473278)
李遠,男,學士,研究方向:藥物分析,E-mail:yuan722914@163.com;黃潔瓊為本文并列第一作者
汪維鵬,男,博士,研究方向:藥物分析,E-mail:wangweipeng@suda.edu.cn;張洪建,男,博士,研究方向:藥物分析,E-mail:zhanghongjian@suda.edu.cn
猜你喜歡
世界科學技術-中醫藥現代化(2022年2期)2022-05-25 13:17:14
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
昆明醫科大學學報(2021年12期)2021-12-30 07:00:10
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
實用口腔醫學雜志(2017年6期)2017-09-19 02:51:06
汽車工程學報(2017年2期)2017-07-05 08:13:02
中外醫療(2016年15期)2016-12-01 04:25:50
