
里氏木霉QM9414尿嘧啶缺陷型轉化體系改進和β-葡萄糖苷酶的過量表達
2016-07-07 12:13:02鐘立霞錢遠超戴美學鐘耀華山東師范大學生命科學學院山東濟南5004山東大學生命科學學院微生物技術國家重點實驗室山東濟南5000
化工學報 2016年6期
鐘立霞,錢遠超,戴美學,鐘耀華(山東師范大學生命科學學院,山東 濟南 5004;山東大學生命科學學院,微生物技術國家重點實驗室,山東 濟南 5000)
?
里氏木霉QM9414尿嘧啶缺陷型轉化體系改進和β-葡萄糖苷酶的過量表達
鐘立霞1,2,錢遠超2,戴美學1,鐘耀華2
(1山東師范大學生命科學學院,山東 濟南 250014;2山東大學生命科學學院,微生物技術國家重點實驗室,山東 濟南 250100)
摘要:里氏木霉是廣泛應用于纖維素酶生產的工業真菌,但高產突變株遺傳操作困難,限制了菌種改良。首先利用定點整合策略敲除里氏木霉突變株QM9414的pyr4基因,成功構建尿嘧啶營養缺陷型菌株QP4,其遺傳轉化效率顯著提高,而且產酶能力不受影響。進一步在QP4中過量表達β-葡萄糖苷酶(BGL)基因bgl1,經大量平板顯色篩選獲得2株BGL活力明顯增強的工程菌QPB4和QPB5,其酶活分別提高10.01倍和8.26倍。利用發酵酶液對2種不同預處理的玉米芯底物進行水解糖化實驗,結果顯示以酸處理玉米芯為底物時QPB4和QPB5的葡萄糖得率比QP4分別提高60.98%和52.44%,而以脫木素處理玉米芯為底物時其葡萄糖得率分別提高80.01%和86.00%。研究表明改進里氏木霉高產突變株遺傳轉化體系可以顯著促進菌株改良,提高糖化應用效果。
關鍵詞:纖維素酶;里氏木霉高產突變株;遺傳轉化;水解糖化;尿嘧啶營養缺陷型標記;構建和表達;生物技術;生物工程
2015-11-03收到初稿,2015-12-26收到修改稿。
聯系人:戴美學,鐘耀華。第一作者:鐘立霞(1989—),女,碩士研究生。
Received date: 2015-11-03.
Foundation item: supported by the National Natural Science Foundation of China (31370135), the Agricultural Science and Technology Achievement Transformation Fund of Shandong Province (2014No.45) and the Fundamental Research Funds of Shandong University (2015CJ005).
引 言
將生物質轉變為燃料乙醇是獲得可再生性能源的重要途徑,越來越受到世界各國的關注。而纖維素酶系是纖維素乙醇化過程中的關鍵酶,其生產成本過高限制了纖維乙醇的發展[1]。改良纖維素酶系,提高底物糖化效率,對降低酶生產成本、促進纖維素轉化具有重要意義。目前纖維素酶生產的工業真菌包括木霉、青霉和曲霉等[2],其中里氏木霉(Trichoderma reesei)以蛋白質分泌量大而成為纖維素酶生產的主要工業菌株[3]。因此,里氏木霉的菌種改良是構建高效纖維素酶系,提高糖化效率的最有效的途徑。
里氏木霉野生型菌株QM6a是第二次世界大戰期間從腐爛的軍人服裝和帆布帳篷中分離到的一種深綠色真菌[4]。隨后經過不斷的紫外線、NTG、離子誘變等篩選,獲得了NG14、RUT-C30和QM9414等高產突變菌株[5]。RUT-C30和QM9414是當前常用的里氏木霉突變菌株。過度的誘變在獲得高纖維素酶的同時也給菌株帶來了諸多負面影響,如經過多輪誘變獲得的RUT-C30雖然比QM9414的胞外蛋白質和纖維素酶活都大幅度提高[5],但其生長緩慢、生孢延遲、生孢減少。而QM9414菌株生長快、產孢多、發酵成熟,是目前進行酶系改良和纖維素酶表達調控機制研究的重要模式菌株。
當前QM9414的遺傳轉化操作主要依賴潮霉素B(hygromycin B)等抗性篩選,但抗生素價格昂貴、篩選背景較高且不方便實現無痕遺傳操作,而營養缺陷型篩選標記可以克服上述的不足,有利于構建高效遺傳轉化體系[6-7]。營養缺陷型篩選標記乳清酸核苷-5′-磷酸脫羧酶(Pyr4/PyrG:木霉為Pyr4,曲霉為PyrG)是尿嘧啶核苷酸合成的一個關鍵酶,該基因的缺失將導致尿嘧啶核苷酸合成受阻,因此缺乏該酶的營養缺陷型菌株需要添加尿嘧啶/尿苷才能生長。當pyr4基因成功轉化進pyr4基因缺失菌株后,該基因的表達使受體菌能夠自身合成尿嘧啶/尿苷,從而不依賴尿嘧啶/尿苷生長,起到正向篩選的作用。另一方面,5′-氟乳清酸(5′-fluorooroticacid,5′-FOA)是尿嘧啶合成前體的類似物,在pyr4的作用下會產生對細胞有毒害物質,所以野生型菌株在5′-FOA下不能生長,而變成pyr4基因缺失菌時就呈現5′-FOA抗性,從而實現反向篩選[8-9]。目前常用的里氏木霉尿嘧啶營養缺陷型菌株是TU6,該菌是由QM9414經射線誘變而來[10],盡管TU6遺傳轉化效率較高,但菌株生長緩慢、生孢量少,尤其是嚴重影響了纖維素酶的表達和分泌。因此,在QM9414中定點敲除pyr4基因,建立高效遺傳轉化體系,有利于通過基因工程手段實現工業菌株的纖維素酶系改良[6,9-13]。
里氏木霉分泌的纖維素酶主要包括3類:外切纖維素酶(CBH,EC3.2.1.91)、內切纖維素酶(EG,EC3.2.1.4)和β-葡萄糖苷酶(BGL,EC3.2.1.21)。在降解復雜底物時BGL能夠將纖維寡糖水解為葡萄糖,從而解除了纖維寡糖對外切酶的抑制。普遍認為里氏木霉降解纖維素的瓶頸是BGL的不足[14]。向里氏木霉酶系中外源補加BGL能夠顯著提高水解纖維素作底物時的葡萄糖得率[15]。另外,在里氏木霉中本源和異源表達BGL1可以提高其纖維素酶系降解纖維素底物的能力[16-17],但目前纖維素酶系組分中的BGL表達水平仍然不高。因此,在里氏木霉中高水平表達BGL是構建高產纖維素酶菌株和提升纖維素酶系整體糖化效率的關鍵。
為了實現遺傳改良里氏木霉從而提高纖維素底物的糖化能力,本研究首先利用里氏木霉QM9414構建了用于高效遺傳轉化的尿嘧啶營養缺陷型菌株,在此基礎上過表達β-葡萄糖苷酶BGL1,通過大量篩選轉化子獲得高水平表達β-葡萄糖苷酶的工程菌株,并進一步利用不同預處理的玉米芯底物進行了糖化應用。
1 實驗材料和方法
1.1材料
1.1.1菌株與質粒里氏木霉(Trichoderma reesei)QM9414來源于美國菌種保藏中心(ATCC26921),TU6菌株來自奧地利維也納技術大學Christian Kubicek教授惠贈。質粒pTHB[18]、pAB4-1和pAN7-1為本實驗室保存。
1.1.2培養基PDA產孢培養基(g·L?1):去皮土豆薄片200,加水煮沸30 min,8層紗布過濾,濾液添加葡萄糖20,并補足水分至1 L,自然pH,2%瓊脂粉。115℃滅菌30 min。
MM培養基(g·L?1):葡萄糖20,KH2PO415,(NH4)2SO45,CaCl20.6,MgSO4·7H2O 0.6,蛋白胨2,FeSO4·7H2O 0.005,MnSO4·H2O 0.0016,ZnSO4·7H2O 0.0014,CoCl2·2H2O 0.002,調節pH 至4.5~5.5,配固體培養基需添加2%瓊脂粉,菌株表型分析時將葡萄糖替換為其他碳源。115℃滅菌30 min。
里氏木霉轉化培養基(g·L?1):葡萄糖 10,MgSO4·7H2O 1,KH2PO410,(NH4)2SO46,C6H5O7Na3·2H2O 3,FeSO4·7H2O 0.005,MnSO4·H2O 0.0016,ZnSO4·7H2O 0.0014,CoCl2·2H2O 0.002,調節pH 至4.5~5.5,轉化培養基加入滲透壓穩定劑山梨醇182.18、瓊脂粉12。115℃滅菌30 min。
LB培養基(g·L?1):酵母粉5,蛋白胨10,NaCl 10,用NaOH調節pH至7.2,固體培養基添加2%瓊脂粉。115℃滅菌30 min。若需要加入氨芐青霉素至終濃度100 μg·ml?1。
七葉苷培養基(g·L?1):CMC-Na 10,七葉苷3,檸檬酸鐵0.5,(NH4)2SO42,MgSO4·7H2O 0.5,K2HPO41,酵母膏 1,2.5%瓊脂粉。115℃滅菌30 min。
種子培養基(g·L?1):葡萄糖 10,CaCl21,MgSO4·7H2O 0.6,KH2PO45,(NH4)2SO42.5,玉米漿20。115℃滅菌30 min。
發酵培養基(g·L?1):結晶纖維20,CaCl21,MgSO4·7H2O 0.6,KH2PO45,(NH4)2SO42.5,玉米漿20。115℃滅菌30 min。
以上培養基在培養尿嘧啶缺陷型時均需添加10 mmol·L?1的尿嘧啶。
1.1.3玉米芯底物用于纖維素酶糖化效率測定的酸處理玉米芯和脫木素處理玉米芯均來自山東龍力生物科技股份有限公司,其中酸處理玉米芯的處理方式采用Schell等[19]的方法,脫木素處理玉米芯的處理方式參照Zhao等[20]的方法。酸處理玉米芯的處理方式其化學成分為:纖維素62.6%,半纖維素2.4%,木素17.7%,灰分6.8%,其他成分為10.5%;脫木素處理玉米芯的處理方式其化學成分為纖維素65.7%,半纖維素1.8%,木素3.2%,灰分5.9%,其他成分為23.4%[21]。
1.1.4酶和試劑里氏木霉原生質體制備所用的哈茨木霉裂解酶,酶活測定所用的pNPC、pNPG、CMC-Na購自SIGMA;酶活測定所用濾紙為Whaterman 1號濾紙;瓊脂糖凝膠回收純化、PCR純化和質粒小提試劑盒均購自OMEGA;DNA聚合酶HiFiTaq polymerase、EasyTaq polymerase 購自TransGen;DNA GeneRuler 1 kb購自ThermoFisher Scientific;潮霉素B(Hygromycin B)購自Roche;尿嘧啶(Uracil)、5′-氟乳清酸(5′-FOA)購自BB;引物由北京六合華大基因科技股份有限公司合成。1.1.5儀器設備臺式離心機(上海安亭科學儀器廠),高速冷凍離心機(Eppendorf),恒溫搖床(上海博彩生物科技有限公司),DYY-Ⅱ電泳儀(北京君意東方電泳設備有限公司),PCR儀(Eppendorf)。
1.2方法
1.2.2里氏木霉基因組提取里氏木霉染色體的提取參照文獻[22]的方法進行。
1.2.3引物設計在里氏組數據庫(http://genome. jgi-psf.org/Trire2/Trire2.home.html)中,使用BLASTN程序搜索與釀酒酵母pryG的同源基因ura3(GenBank accession no. NC_001137.3)的基因序列同源性最高的序列。結果顯示編號為74020的蛋白同源性最高,根據注釋為乳清酸-5′-單磷酸脫羧酶,即pyr4基因序列信息,運用Primer Premier 5軟件設計引物,本研究所用的引物序列見表1。
1.2.4pyr4敲除盒的構建和黑曲霉pyrG基因片段的擴增以里氏木霉QM9414基因組為模板,利用引物pyr4-UF1/pyr4-UR1和pyr4-DF1/pyr4-DR1分別擴增得到1.3 kb的pyr4基因上游同源臂和1.3 kb 的pyr4基因下游同源臂;以PAN7-1質粒為模板,利用引物hph-F1/hph-R1擴增得到2.8 kb的潮霉素B基因。然后,利用Double-joint PCR[23]將pyr4基因上下游同源臂和潮霉素B基因利用引物pyr4-UF3/pyr4-UR3融合得到5.2 kb的pyr4敲除盒。

表1 本研究所用引物Table 1 Primers used in this study
以質粒pAB4-l為模板,利用引物pyrG-S/pyrG-A擴增得到2.7 kb的黑曲霉pyrG基因片段,用于回補pyr4敲除菌株以及與質粒pTHB共轉化pyr4敲除菌株,從而構建BGL1過表達菌株。
1.2.5pyr4敲除菌株、pyrG回補菌株和BGL1過表達菌株的構建里氏木霉原生質體的制備和轉化實驗主要參照文獻[22]的方法進行。
pyr4敲除菌株的構建:將1μg pyr4敲除盒與適量里氏木霉QM9414原生質體、100 μg·ml?1潮霉素B、1.5 mg·ml?15′-FOA混合,傾倒在已凝固的轉化平板上,30°C培養至轉化子出現。
pyrG回補菌株的構建:1 μg pyrG基因片段與尿嘧啶營養缺陷型菌株QP4原生質體傾倒在不含有尿嘧啶的轉化下層平板上,30℃培養至轉化子出現。
對于初中生而言,年齡相對較小,對任何事情都具有著很大的好奇心,因此只要初中語文教師注重活躍課堂氛圍,就很容易將學生帶到相應的情境之中,激發他們的學習興趣。在過去傳統的初中語文教學中,教師并不重視課堂氛圍,只是簡單地講授知識點,造成學生認為語文學習枯燥乏味,出現排斥心理,影響到初中語文的教學質量。因此,為了打破過去教學方法的束縛,教師應該積極使用多元化的教學方法。在講課的過程中,教師可以面帶微笑,在課堂上營造出輕松、活躍的課堂氛圍,通過提問或是小故事等形式引導學生積極發言,多與學生進行互動。例如在講述詩歌古文時,教師可以以小組為單位舉行朗誦比賽,讓學生們扮演詩人來抒發自己的情懷等。
BGL1過表達菌株的構建:將2 μg含有Pcbh1啟動子控制表達bgl1表達盒(Pcbh1-bgl1)的pTHB質粒[18]和1 μg pyrG基因片段同時與適量尿嘧啶營養缺陷型菌株QP4原生質體傾倒在不含有尿嘧啶的轉化平板上,30℃培養至轉化子出現。
1.2.6PCR驗證、單孢分離和菌株表型分析PCR驗證:參照1.2.2節的方法提取染色體,pyr4敲除菌株的驗證利用pyr4敲除盒中潮霉素B基因驗證引物Yhph-F1/Yhph-R1,上游染色體錨定引物Ypyr4-UF2/Yhph-R1和下游染色體錨定引物Yhph-F2/pyr4-DR1分別擴增800 bp、3.1 kb、2.1 kb大小片段。回補菌株的驗證PCR引物為YpyrG-F1/YpyrG-R1擴增600 bp左右。BGL過表達菌株的驗證以Y1/Y2為引物擴增Pcbh1-bgl1表達盒中橫跨啟動子和編碼區的1.0 kb片段。
單孢分離:將孢子懸液稀釋一定倍數后,在添加有0.5%的TritonX-100、1 mg·ml?1的尿嘧啶的MM固體培養基上涂覆或劃線培養,挑取單菌落進行PCR驗證,將分離純化得到的陽性轉化子命名為QP4并生孢保存。回補菌株及BGL過表達菌株單孢分離時MM培養基上不需添加1 mg·ml?1的尿嘧啶。
表型驗證:取1 μl新鮮孢子(或取菌絲塊),在加有尿嘧啶的MM平板上30℃培養4 d。
1.2.7生長及碳源利用情況測定菌落大小測定:取1 μl新鮮孢子點至不同碳源(葡萄糖、甘油、乳糖、纖維素)MM培養基和PDA培養基上,30℃培養60 h,記錄菌落形態并測量菌落直徑來表征菌落的大小。
1.2.8纖維素酶活測定取新鮮孢子懸液,以106個·ml?1的接種量接種于種子培養基,30℃、200 r·min?1振蕩培養36 h,以10%的接種量轉接至發酵培養基,30℃,200 r·min?1振蕩培養。誘導一定時間后取上清測定濾紙酶活(FPA)、內切葡聚糖酶活(EG)、外切葡聚糖酶活(CBH)、β-葡萄糖苷酶活(BGL),FPA、EG活力測定參照曲音波等[24]的方法進行,CBH、BGL活力測定參照文獻[25]。
1.2.9BGL表達菌株的平板定性檢測將β-葡萄糖苷酶過表達菌株在β-葡萄糖苷酶篩選培養基上30℃培養12 h、24 h,記錄菌落形態并測量菌落直徑大小來表征β-葡萄糖苷酶活。
1.2.10不同底物糖化分析糖化實驗參照Cheng 等[26]的方法進行。反應體系為30 ml,糖化底物為5%的酸處理玉米芯、脫木素處理玉米芯,纖維素酶添加量為10 FPU·g?1底物,為了防止染菌加入0.1% NaN3,用0.1 mol·L?1pH4.8的醋酸緩沖液補齊至30 ml,50℃、150 r·min?1進行糖化,取糖化24 h、48 h糖化液,利用SBA-40C(BISAS,China)生物傳感儀進行葡萄糖的測定。

圖1 里氏木霉pyr4敲除菌株QP4的PCR驗證圖及表型差異比較Fig.1 PCR confirmation and phenotypic assay of pyr4 deletion strain
2 實驗結果與討論
2.1里氏木霉菌株的pyr4敲除
為了在里氏木霉QM9414中敲除pyr4基因,首先利用該菌基因組擴增得到均為1.3 kb的pyr4基因上下游同源臂與pAN7-1質粒擴增得到的2.8 kb潮霉素B抗性表達元件經融合PCR得到5.2 kb的pyr4敲除盒。然后以pyr4敲除盒轉化里氏木霉QM9414原生質體,在100 μg·ml?1潮霉素B、1.5 mg·ml?15′-FOA雙篩選壓力下初步得到2株轉化子,經過復篩、單孢分離,隨后挑取1株遺傳穩定的轉化子命名為QP4,并進行PCR驗證(圖1)。結果顯示,QP4轉化子中可以擴增得到敲除盒潮霉素編碼區800 bp[圖1(a)左]、染色體上游錨定片段3.0 kb[圖1(a)中]和染色體下游錨定片段1.8 kb左右的條帶[圖1(a)右],而出發菌QM9414染色體和陰性對照H2O為模板時沒有擴展片段的條帶,與預期結果符合。這表明pyr4敲除盒已經發生同源交換,成功插入里氏木霉QM9414染色體中,并且染色體中沒有其他位點的片段整合,從而在染色體水平驗證了pyr4基因被定點敲除。
為了進一步確認pyr4基因的敲除,用來自黑曲霉(Aspergillus niger)的pyr4同源基因pyrG對QP4菌株進行回補。轉化后在無尿嘧啶篩選平板上得到17個轉化子,經過PCR擴增pyrG基因驗證均為陽性轉化子,表明利用pyr4作為正向篩選基因可大大提高轉化效率。隨機挑取一株轉化子命名為R-pyrG,經單孢分離傳代后進行表型分析。圖1(b)結果顯示,出發菌株QM9414和回補菌株R-pyrG表型相似,在加有尿嘧啶的MM平板和不含尿嘧啶的MM平板上都生長,而敲除pyr4的QP4菌株在加有尿嘧啶的MM上平板生長,在不含尿嘧啶的MM平板上不生長。該結果說明R-pyrG能夠恢復到野生型狀態,QP4菌株的表型變化確實是由pyr4基因敲除造成的。因此,本研究成功構建了里氏木霉pyr4基因敲除的并可用于高效遺傳轉化的尿嘧啶營養缺陷型菌株QP4。
2.2不同碳源條件下QP4生長情況分析
為驗證pyr4的敲除是否影響菌株的生長,本研究分析了QP4在不同碳源條件下的生長情況(圖2)。與出發菌株QM9414相比,pyr4敲除菌株QP4的菌落大小、菌落顏色和菌落蔓延狀態在不同碳源(葡萄糖、甘油、乳糖、纖維素和PDA)上均變化不大,而誘變的尿嘧啶營養缺陷型菌株TU6菌落明顯變小且菌落顏色也有差異。其中各菌株在葡萄糖、PDA上的生長速度最快,乳糖、纖維素上次之,甘油上最慢。另外,TU6的生孢量明顯低于QP4和QM9414。對菌落大小的測定結果顯示,菌株QP4菌落大小與QM9414基本一致,而TU6菌落在葡萄糖、甘油、乳糖、纖維素條件下分別減小了35.8%、28.1%、15.5%和12.5%[圖2(b)]。以上結果顯示,不同碳源條件下pyr4敲除菌株QP4的生長并沒有受到影響,可用于后續研究。
2.3搖瓶發酵條件下QP4產酶情況分析
為進一步確認pyr4敲除對產酶能力的影響,本研究對QP4在產酶發酵條件下7 d的纖維素酶活及各組分酶活進行測定。如圖3所示,pyr4敲除菌株QP4的濾紙酶活(FPA)、內切葡聚糖酶活(EG)、外切葡聚糖酶活(CBH)和β-葡萄糖苷酶活(BGL)與出發菌株QM9414相比并未發生明顯變化,卻都顯著高于誘變菌株TU6。其中,QP4的FPA和CBH分別是TU6的2.13倍和2.12倍,而EG和BGL分別是TU6的1.36倍和1.35倍。該結果表明,pyr4的敲除并未影響里氏木霉原有纖維素酶的表達和分泌,而且明顯優于誘變菌株TU6。因此,QP4可用于對里氏木霉進行高產纖維素酶菌種改良的后續遺傳操作。
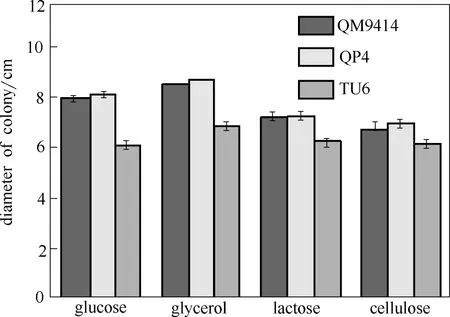
圖2 里氏木霉pyr4敲除菌株QP4在不同碳源下的表型分析和生長差異比較Fig.2 Phenotypic analysis and growth comparison of QM9414, QP4 and TU6 strains on different carbon sources after 60 h (Data are represented as mean of three independent experiments. Error bars express standard deviations)
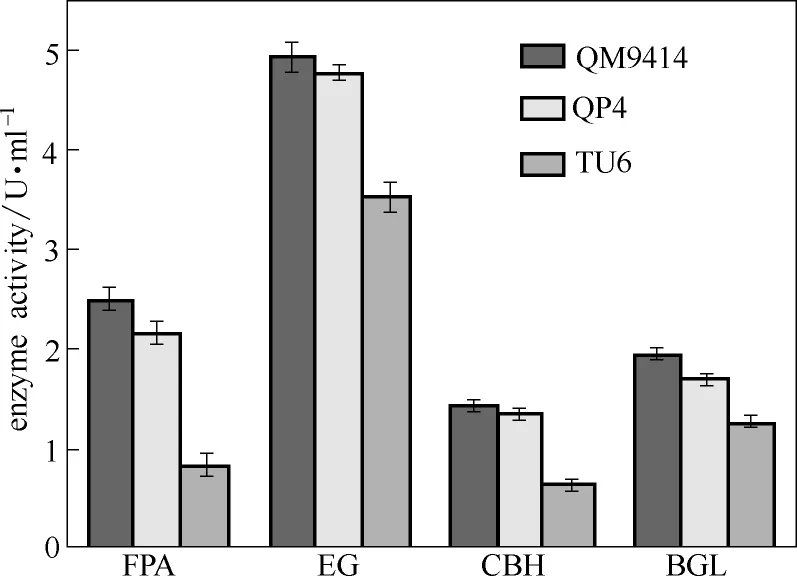
圖3 里氏木霉pyr4敲除菌株纖維素酶活力分析Fig.3 Cellulase activities of T. reesei QP4, TU6 and QM9414 [Filter paper activity (FPA), endoglucanase activity (EG), cellobiohydrolase activity (CBH) and β-glucosidase activity (BGL) were measured after being induced with cellulase production medium for 7 d. Data are represented as mean ofthree independent experiments. Error bars expressstandard deviations]
2.4QP4菌株過表達β-葡萄糖苷酶改良纖維素酶系
為進一步驗證QP4菌株作為高效遺傳轉化體系的可操作性,同時改良里氏木霉纖維素酶系組分,本研究針對里氏木霉β-葡萄糖苷酶(BGL)表達量低的問題,利用帶有強啟動子Pcbh1控制表達β-葡萄糖苷酶基因bgl1表達盒(Pcbh1-bgl1)的質粒pTHB[15]與來自黑曲霉的pyrG片段共轉化QP4。轉化平板上獲得了135個轉化子,經過PCR驗證,123個為pyrG整合入染色體的陽性轉化子,陽性轉化子率達到91.1%,遠高于潮霉素B抗性篩選的陽性轉化子效率(63%)[12]。這表明QP4可以作為pyr4篩選標記的高效遺傳轉化菌株。
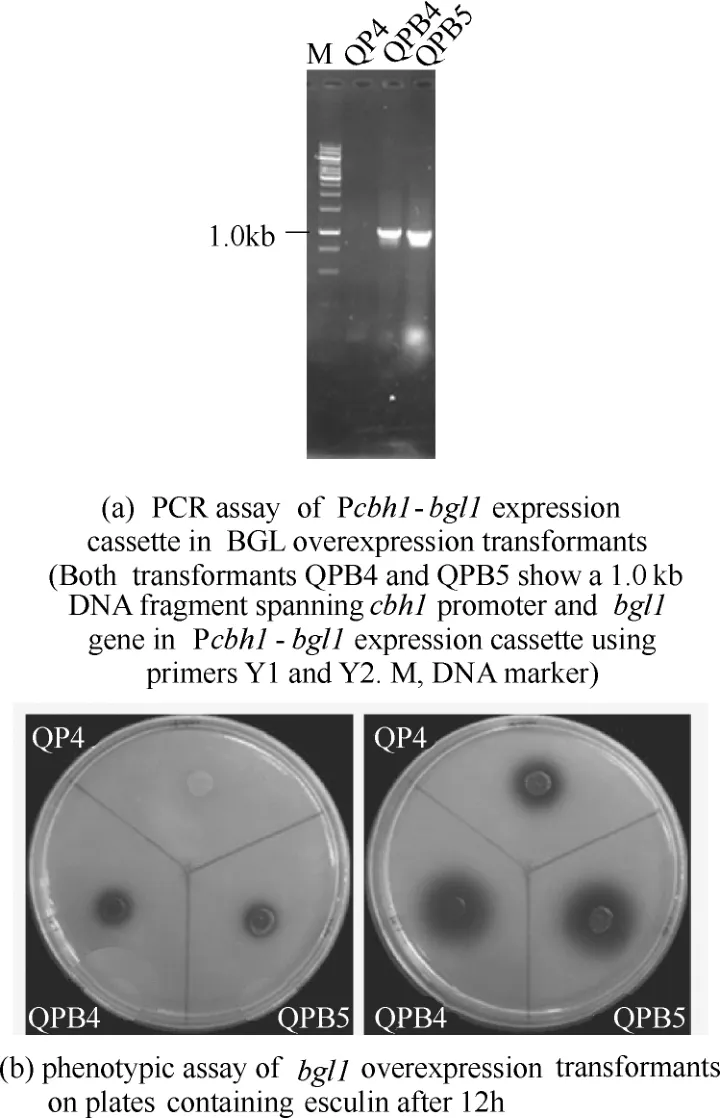
圖4 里氏木霉β-葡萄糖苷酶過表達菌株的分子驗證和平板BGL活力檢測Fig.4 PCR analysis and detection of BGL activity on esculin substrate in bgl1 overexpression transformants
β-葡萄糖苷酶(BGL)能夠將七葉苷分解為七葉素,與Fe3+螯合形成黑色的螯合物,在培養基平板上出現明顯的黑色暈圈,因此利用七葉苷培養基平板可以進行高通量BGL表達菌株的篩選。本研究將獲得的pyrG整合陽性轉化子進行了快速七葉苷平板檢測,挑選出2株BGL酶活顯著增強的轉化子,分別命名為QPB4和QPB5。然后進行單孢分離和PCR驗證,證明Pcbh1控制表達的bgl1基因成功整合到染色體中[圖4(a)]。表型分析表明,QPB4和QPB5轉化子在無尿嘧啶培養基上可以生長,而QP4不能生長,說明篩選標記pyrG基因在轉化子中穩定遺傳。七葉苷平板測定顯示,QPB4 和QPB5在12h時能夠形成明顯的黑色暈圈,而QP4菌株未發現明顯的黑色暈圈;24 h時QPB4和QPB5的黑色暈圈的直徑分別達到了1.3 cm和1.6 cm,而QP4的黑色暈圈的直徑為0.7 cm[圖4(b)],這說明BGL在QP4中成功實現了過表達。
2.5β-葡萄糖苷酶過表達菌株的產酶分析
為進一步確認BGL過表達菌株的纖維素酶產生情況,取搖瓶發酵培養5 d的酶液進行酶活測定。如圖5所示,過表達菌株QPB4和QPB5的β-葡萄糖苷酶活力分別達到了13.65 U·ml?1和11.49 U·ml?1,比QP4(1.24 U·ml?1)分別提高了10.01倍和8.26倍。但其濾紙酶活變化有差異,QPB5的FPA為2.14 U·ml?1,比QP4提高了43.6%,而QPB4的FPA為1.29 U·ml?1,比QP4卻降低了13.4%。QPB4產生這種情況的原因可能是由于其染色體中bgl1表達盒的不合理插入或cbh1啟動子過表達引起轉錄調控因子滴定效應等導致的[27]。但測定QPB4的纖維素外切酶活和內切酶活,發現基本沒有發生變化[圖5(b)]。這種過表達一個纖維素酶基因而引起總纖維素酶活降低的現象在前期研究中也有報道[14]。而通過大量篩選得到的另一菌株QPB5并沒有發生這種情況,BGL和FPA活力均顯著提高(如上所述)。
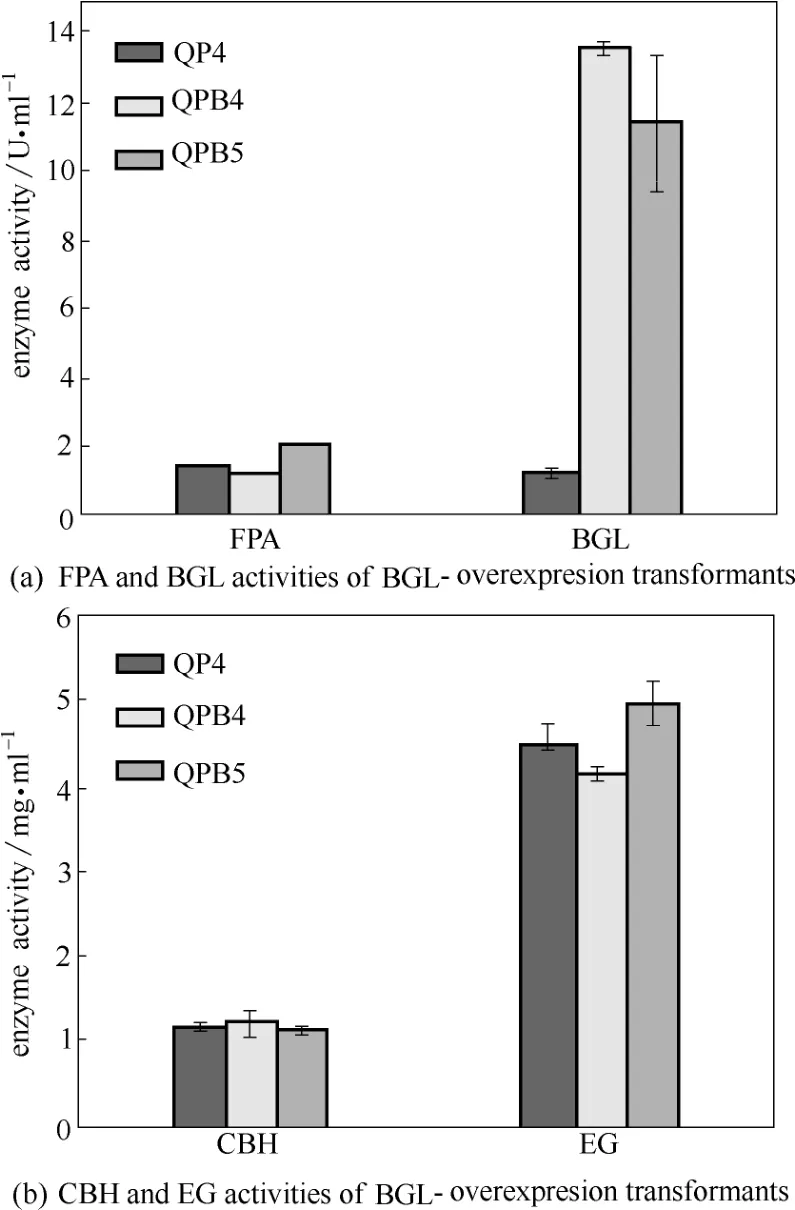
圖5 里氏木霉β-葡萄糖苷酶過表達菌株纖維素酶活測定Fig.5 Enzyme activities of BGL-overexpresion transformants (FPA, BGL, CBH and EG were measured after being induced in cellulase fermentation medium for 5 d. Data are represented as mean of three independent experiments. Error bars express standard deviations)
2.6β-葡萄糖苷酶過表達菌株的糖化應用
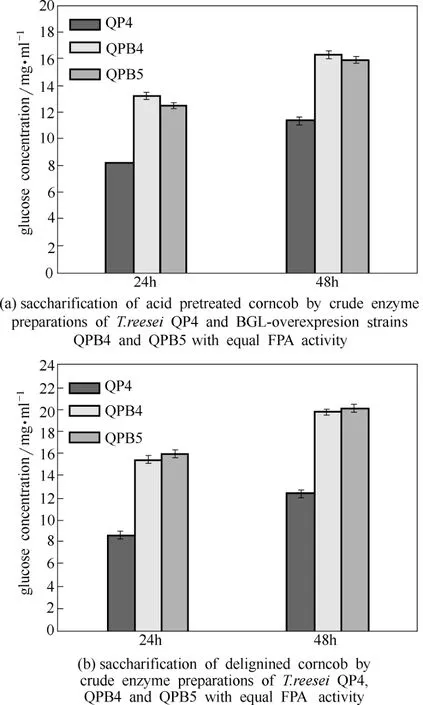
圖6 過表達β-葡萄糖苷酶菌株發酵酶液對玉米芯材料的糖化應用Fig.6 Saccharification of different pretreated corncob residues by crude enzyme preparations of BGL-overexpression strains (Data are represented as mean of three independent experiments. Error bars express standard deviations)
為了檢測里氏木霉高水平過表達β-葡萄糖苷酶對改良纖維素酶系的效果,本研究采用2種不同預處理的玉米芯材料作為糖化底物,測定了β-葡萄糖苷酶過表達菌株發酵酶液的糖化效率(圖6)。結果顯示,當以酸處理玉米芯作為底物時,糖化效率在24 h和48 h時均有大幅提升,尤其是24 h時更加明顯,QPB4和QPB5發酵酶液的葡萄糖得率分別為13.20 mg·ml?1和12.50 mg·ml?1,比QP4 (8.20 mg·ml?1)分別提高了60.98%和52.44%[圖6(a)]。當以脫木素處理玉米芯作為底物糖化時,過表達β-葡萄糖苷酶菌株提高糖化效率更加明顯,24 h時QPB4(15.56 mg·ml?1)和QPB5(16.07 mg·ml?1)比QP4(8.64 mg·ml?1)分別提高了80.09%和86.00%[圖6(b)]。QPB4和QPB5在糖化2種底物時,雖然葡萄糖得率均有大幅提升,但也出現一定差異,如QPB4糖化酸處理玉米芯底物效果比QPB5好,而QPB5糖化脫木素處理玉米芯底物效果比QPB4好,這可能與兩個工程菌株酶系差異有關。總之,本研究成功實現了里氏木霉QM9414遺傳操作體系的改進,并以此為基礎改良纖維素酶組分,獲得了糖化效率顯著提高的工程菌,為下一步實際工業應用打下基礎。
3 結 論
(1)通過定點敲除pyr4基因建立了里氏木霉高產突變株QM9414的尿嘧啶缺陷菌株QP4,顯著提高了遺傳轉化效率,而且沒有影響菌株生長和纖維素酶表達,成為替換尿嘧啶缺陷誘變菌株TU6進行遺傳改良和纖維素酶表達機制研究的優良菌株。
(2)利用構建的尿嘧啶缺陷菌株QP4進行本源bgl1過表達,通過高效轉化和篩選,僅進行一輪轉化便獲得了135個轉化子,其中123株為遺傳穩定轉化子,從中進一步篩選出2株BGL活力最高的菌株,活力分別提高了10.01倍和8.26倍。這說明本研究對里氏木霉QM9414菌株的遺傳轉化體系改進是成功的,同時高水平過表達了β-葡萄糖苷酶。在進一步糖化應用方面,本研究獲得的QPB5菌株發酵酶液將玉米芯底物水解的葡萄糖得率在24 h時就達到了16.07 mg·ml?1,比出發菌株提高了86.00%,這在本源過表達β-葡萄糖苷酶菌株提高糖化效率方面是非常顯著的。
本研究借助轉化體系改進和高水平過表達β-葡萄糖苷酶解決了里氏木霉高產突變株遺傳操作困難和酶系組分缺陷兩個限制菌種改良的因素,顯著提升了里氏木霉的糖化應用能力。同時,高效遺傳轉化菌株的構建和高產菌株的獲得也為研究里氏木霉纖維素酶高產機制以及進一步的實際生產應用打下基礎。
References
[1]鐘耀華, 錢遠超, 任美斌, 等.絲狀真菌降解轉化纖維素的機制與遺傳改良前景 [J]. 生物加工過程, 2014, 12 (1): 46-54. ZHONG Y H, QIAN Y C, REN M B, et al. Progress in bioconversion of cellulose by filamentous fungi and related genetic engineering strategies [J]. Chinese Journal of Bioprocess Engineering, 2014, 12 (1): 46-54
[2]CHANDELA K, CHANDRA S G, SILVA M B, et al. The realm of cellulases in biorefinery development [J]. Crit. Rev. Biotechnol., 2012, 32 (3): 187-202.
[3]LYND L R, WEIMER P J, VAN W H, et al. Microbial cellulose utilization: fundamentals and biotechnology [J]. Microbiol. Mol. Biol. Rev., 2002, 66 (3): 506-577.
[4]MANDELS M, REESE E T. Induction of cellulase in Trichoderma viride as influenced by carbon sources and metals [J]. J. Bacteriol., 1957, 73 (2): 269-278.
[5]PETERSON R, NEVALAINEN H. Trichoderma reesei RUT-C30: thirty years of strain improvement [J]. Microbiology, 2012, 158 (1): 58-68.
[6]陳永, 林良才, 肖冬光, 等. 里氏木霉RL-P37尿嘧啶營養缺陷型菌株的構建 [J].中國釀造, 2014, 33 (12): 58-62. CHEN Y, LIN L C, XIAO D G, et al. Construction of uracil-auxotrophic strain of Trichoderma reesei RL-P37 [J]. China Brewing, 2014, 33 (12): 58-62.
[7]LONG H, WANG T H, ZHANG Y K. Isolation of Trichoderma reesei pyrG negative mutant by UV mutagenesis and its application in transformation [J]. Chem. Res. Chinese Universities, 2008, 24 (5): 565-569.
[8]DU Y, XIE G, YANG C, et al. Construction of brewing-wine Aspergillus oryzae pyrG-mutant by pyrG gene deletion and its application in homology transformation [J]. Acta Biochim. Biophys. Sin. (Shanghai), 2014, 46 (6): 477-483.
[9]HIRASHIMA K, IWAKII T, TAKEGAWA K, et al. A simple and effective chromosome modification method for large-scale deletion of genome sequences and identification of essential genes in fission yeast [J]. Nucleic Acids Research, 2006, 34 (2): e11.
[10]GRUBER F, VISSER J, KUBICEK C P, et al. The development of a heterologous transformation system for the cellulolytic fungus Trichoderma reesei based on a pyrG-negative mutant strain [J]. Curr. Genet., 18 (1): 71-76.
[11]DELMAS S, LLANOS A, PARROU J L, et al. Development of an unmarked gene deletion system for the filamentous fungi Aspergillus niger and Talaromyces versatilis [J]. Appl. Environ. Microbiol., 2014, 80 (11): 3484-3487.
[12]DERNTL C, KIESENHOFER D P, MACH R L, et al. Novel strategies for genomic manipulation of Trichoderma reesei with the purpose of strain engineering [J]. Appl. Environ. Microbiol., 2015, 81 (18): 6314-6323.
[13]安莉穎, 易欣欣, 秦麗娜, 等. 里氏木霉RutC30常溫常壓等離子體(ARTP)誘變篩選pyr4基因缺陷型菌株及轉化系統的建立 [J].菌物學報, 2014, 33 (6):1672-6472. AN L Y, YI X X, QIN L N, et al. Isolation of Trichoderma reesei RutC30 pyr4 gene mutant by atmospheric room temperature plasma (ARTP) and establishment of a transformation system [J]. Mycosystema, 2014, 33 (6): 1672-6472.
[14]SIPOS B, BENKO Z, DIENES D, et al. Characterisation of specific activities and hydrolytic properties of cell-wall-degrading enzymes produced by Trichoderma reesei Rut-C30 on different carbon sources [J]. Appl. Biochem. Biotechnol., 2010, 161 (1-8): 347-364.
[15]SALOHEIMO M, KUJA-PANULA J, YLOSMAKI E,et al.Enzymatic properties and intracellular localization of the novel Trichoderma reesei β-glucosidase BGLII(Cel1A) [J]. Appl. Environ. Microbiol., 2002, 68 (9): 4546-4553.
[16]ZHANG J W, ZHONG Y H, ZHAO X N, et al. Development of the cellulolytic fungus Trichoderma reesei strain with enhanced beta-glucosidase and filter paper activity using strong artificial cellobiohydrolase 1 promoter [J]. Bioresour. Technol., 2010, 101 (24): 9815-9818.
[17]MA L, ZHANG J, ZOU G, et al. Improvement of cellulase activity in Trichoderma reesei by heterologous expression of a beta-glucosidase gene from Penicillium decumbens [J]. Enzyme Microb. Technol., 2011, 49 (4): 366-371.
[18]LIU T, WANG T, LI X, et al. Improved heterologous gene expression in Trichodenna reesei by chhl promoter optimization [J]. Acta Biochim. Biophys. Sin. (Shanghai)., 2008, 40 (2): 158-165.
[19]SCHELL D J, FARMER J, NEWMAN M, et al. Dilute-sulfuric acid pretreatment of corn stover in pilot-scale reactor: investigation of yields, kinetics, and enzymatic digestibilities of solids [J]. Appl. Biochem. Biotechnol., 2003, 105-108: 69-85.
[20]ZHAO X, ZHANG L , LIU D . Comparative study on chemical pretreatment methods for improving enzymatic digestibility of crofton weed stem [J]. Bioresour. Technol., 2008, 99 (9): 3729-3736.
[21]LIU K, LIN X H, YUE J, et al. High concentration ethanol production from corncob residues by fed-batch strategy [J]. Bioresour. Technol., 2010, 101 (13): 4952-4958.
[22]PENTTILA M, NEVALAINEN H, RATTO M, et al. A versatile transformation system for the cellulolytic filamentous fungus Trichoderma reesei [J]. Gene, 1987, 61 (2): 155-164.
[23]YU J H, HAMARI Z, HAN K H, et al. Double-joint PCR: PCR-based molecular tool for gene manipulations in filamentous fungi [J]. Fungal Genet. Biol., 2004, 41 (11): 973-981.
[24]曲音波, 高培基, 王祖農. 青霉的纖維素酶抗降解物阻遏突變株的選育 [J]. 真菌學報, 1984, 3 (4): 238-243. QU Y B, GAO P J,WANG Z N. Screening of catabolite repressionresistant mutants of cellulase producing Penicillium spp [J]. AcfaMycol. Sin., 1984, 3 (4): 238-243.
[25]MURRAY P, ARO N, COLLINS C, et al. Expression in Trichoderma reesei and characterisation of a thermostable family 3 beta-glucosidase from the moderately thermophilic fungus Talaromyces emersonii [J]. Protein Expr. Purif., 2004, 38 (2): 248-257.
[26]CHENG Y, SONG X, QIN Y, et al. Genome shuffling improves production of cellulase by Penicillium decumbens JU-A10 [J]. J. Appl. Microbiol., 2009, 107 (6): 1837-1846.
[27]RAHMANZL, SHIDA Y, FURUKAWA T, et al. Application of Trichoderma reesei cellulase and xylanase promoters through homologous recombination for enhanced production of extracellular beta-glucosidase Ⅰ [J]. Biosci. Biotechnol. Biochem., 2009, 73 (5): 1083-1089.
Improvement of uracil auxotrophic transformation system in Trichoderma reesei QM9414 and overexpression of β-glucosidase
ZHONG Lixia1,2, QIAN Yuanchao2, DAI Meixue1, ZHONG Yaohua2
(1College of Life Sciences, Shandong Normal University, Jinan 250014, Shandong, China;2State Key Laboratory of Microbial Technology, School of Life Sciences, Shandong University, Jinan 250100, Shandong, China)
Abstract:Trichoderma reesei is an industrial filamentous fungus widely used for cellulase production. However, the difficulty in genetic modification of the high-production mutants limits the strain improvement. Firstly, the pyr4 gene was successfully deleted by target integration strategy in T. reesei QM9414 to construct a uracil auxotroph strain QP4. Compared with those in the parental strain, not only the genetic transformation efficiency was improved, but also the cellulase production was not influenced in QP4. The β-glucosidase (BGL) encoding gene bgl1 was further overexpressed in the QP4 strain. The transformants QPB4 and QPB5 with increased BGL activity were obtained by rapid screening based on the esculin plate. The BGL activities in the culture supernatants of QPB4 and QPB5 exhibited 10.01- and 8.26- fold higher than that in QP4, respectively. The crude enzyme preparations were used to saccharify two types of differently pretreated corncob residues. The glucose yields from saccharification of acid-pretreated corncob by QPB4 and QPB5 were 60.98% and 52.44% higher than that by QP4,respectively, while using the delignined corncob as substrate, the glucose yields by QPB4 and QPB5 were improved 80.01% and 86.00%, respectively. These results suggested that improvement of the genetic manipulation system in T. reesei greatly facilitated the strain improvement and increased the saccharification efficiency.
Key words:cellulases; Trichoderma reesei high-production mutant; genetic transformation; saccharification; uracil auxotroph; construction and expression; biotechnology; biological engineering
中圖分類號:TQ 812
文獻標志碼:A
文章編號:0438—1157(2016)06—2510—09
DOI:10.11949/j.issn.0438-1157.20151654
基金項目:國家自然科學基金面上項目(31370135);山東省農業科技成果轉化資金項目(2014No.45);山東大學基本科研業務費專項資金項目(2015CJ005)。
Corresponding author:DAI Meixue, daimeixue@sdnu.edu.cn; ZHONG Yaohua, zhongyaohua@sdu.edu.cn
