兔肝細胞中與RHDV VP60 P2亞區相互作用蛋白質的篩選和初步鑒定
2016-07-13 01:31:05宋艷華范志宇魏后軍仇汝龍
畜牧獸醫學報 2016年3期
李 騰,姜 平,王 芳*,宋艷華,胡 波,范志宇,魏后軍,仇汝龍,劉 星
(1.南京農業大學動物醫學院,南京 210095;2.江蘇省農業科學院獸醫研究所,農業部獸用生物制品工程技術重點實驗室,國家獸用生物制品工程技術研究中心,南京 210014)
?
兔肝細胞中與RHDV VP60 P2亞區相互作用蛋白質的篩選和初步鑒定
李騰1,2,姜平1*,王芳1,2*,宋艷華2,胡波2,范志宇2,魏后軍2,仇汝龍2,劉星2
(1.南京農業大學動物醫學院,南京 210095;2.江蘇省農業科學院獸醫研究所,農業部獸用生物制品工程技術重點實驗室,國家獸用生物制品工程技術研究中心,南京 210014)
摘要:兔出血癥病毒(RHDV)衣殼蛋白VP60 P2亞區在病毒感染宿主細胞過程中可能起重要作用,為找尋RHDV靶細胞——兔肝細胞中與P2亞區相互作用的蛋白質,以原核表達的P2蛋白為誘餌蛋白質,通過免疫共沉淀試驗(Co-IP)結合質譜分析篩選與之有相互作用的肝細胞蛋白質。經分析選取層黏連蛋白受體(LamR)蛋白做進一步研究。提取兔肝細胞總RNA,反轉錄擴增LamR基因,將其插入pGEX-6p-1載體,經IPTG誘導表達,獲得GST-LamR融合蛋白,并通過GST pull-down試驗驗證LamR蛋白與P2蛋白的結合。結果顯示,以P2蛋白為誘餌蛋白質,免疫共沉淀試驗結合質譜分析共篩選到26個與P2存在相互作用的肝細胞蛋白質,其中LamR蛋白是多種病毒的受體。通過原核表達系統表達了LamR蛋白,經pull-down試驗驗證了LamR蛋白與P2蛋白有直接的相互作用。本研究證實兔肝細胞LamR蛋白與RHDV VP60 P2蛋白的相互作用,為深入研究RHDV的感染過程、揭示RHDV的致病機制提供了線索。
關鍵詞:兔出血癥病毒(RHDV);VP60 P2亞區;免疫共沉淀;層黏連蛋白受體(LamR)
兔出血癥(rabbit hemorrhagic disease,RHD)是由兔出血癥病毒(rabbit hemorrhagic disease virus,RHDV)引起的兔的一種急性、敗血性、高度接觸傳染性、致死性疾病,是兔的一種毀滅性傳染病。RHDV屬杯狀病毒科兔病毒屬,無囊膜,為20面體對稱結構。病毒衣殼由180個VP60蛋白形成的90個二聚體組裝而成。VP60蛋白是RHDV的主要結構蛋白,包含形成病毒衣殼內殼的S區和在衣殼外表面形成刺突結構的P區。P區變異性較高,與病毒的抗原性相關,是宿主抗體識別的主要區域并在病毒粒子吸附宿主細胞及隨后的感染致病過程中起重要作用[1-3]。該區又可進一步劃分為P1亞區和P2亞區。P2亞區位于病毒衣殼表面,具有突出的β桶核心結構,推測P2區在病毒整個感染過程中起關鍵作用。此外,P2區還包含7個loop環,其loop區與其他杯狀病毒有顯著不同,這可能是病毒感染宿主特異性的主要決定因素[4]。因此,本研究選用P2亞區蛋白作為誘餌蛋白質,以RHDV的主要靶器官兔肝作為研究對象,通過Co-IP法釣取兔肝細胞中與P2相互作用的蛋白質,以期為研究RHDV的致病機制提供線索。
1材料與方法
1.1載體、蛋白質、菌株及細胞
感受態細胞E.coliDH5α、BL21購自北京全式金生物技術有限公司;供體質粒pET-32a(+)、pGEX-6p-1及真核表達的VP60蛋白[5]、pET-32a(+)表達的重組IFN-γ蛋白均由本實驗室保存。
1.2試劑
Platinum?Pfx高保真DNA 聚合酶(50 U·μL-1)購自Invitrogen公司;抗GST標簽單抗抗體購自北京全式金生物技術有限公司;抗LamR山羊多抗購自Santa Cruz公司;識別VP60-P2區的單克隆抗體1B8由本實驗室制備;柱式DNA質粒OUT試劑盒購自北京天恩澤基因科技有限公司;HiTrapTMProtein G HP純化柱試劑盒、His TrapTMHP 純化柱購自GE公司;Pierce?交聯IP試劑盒購自Thermo Fisher Scientific公司;GST-resin純化樹脂購自上海點創生物科技有限公司;其他試劑均為國產分析純級。
1.3P2亞區基因的克隆
根據GenBank數據庫RHDV皖阜株基因序列(FJ794180,RHDVa型),以pMD19-T-VP60為模板,應用Primer 5.0軟件設計P2亞區基因引物(表1)擴增RHDVVP60 P2亞區基因。
1.4pET-32a-P2重組載體的構建
用瓊脂糖凝膠DNA回收試劑盒回收P2亞區基因擴增產物。pET-32a(+)供體質粒及目的基因擴增產物經EcoRⅠ和XhoⅠ 37 ℃雙酶切3 h,用T4 DNA Ligase 4 ℃連接12 h,將連接產物轉化DH5α感受態大腸桿菌,經質粒提取和雙酶切鑒定后,陽性克隆送上海華津生物科技有限公司測序,測序正確的克隆提取質粒,最終獲得重組質粒,命名為pET-32a-P2。
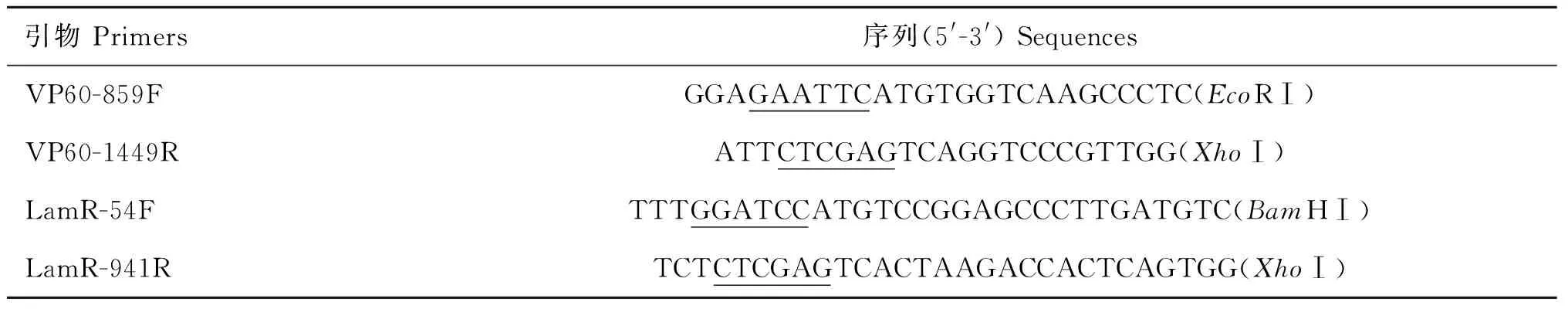
表1 引物序列
下劃線為內切酶位置
The underlined letters show the restriction endonuclease sites
1.5P2重組蛋白質的表達、純化及鑒定
經測序正確的重組質粒轉入BL 21大腸桿菌,以1 mmol·L-1的IPTG 37 ℃誘導表達5 h,離心收集菌體并重懸至原體積的1/5,經超聲波破碎后收集含可溶性表達蛋白質的上清。使用His TrapTMHP 純化柱試劑盒,按照說明對P2重組蛋白質進行純化。SDS-PAGE和Western blotting鑒定P2蛋白的表達及純化效果。
1.6肝細胞全蛋白質的制備
采用膠原酶灌注法分離肝細胞[6]。IP裂解液重懸細胞,4 ℃搖床上孵育10 min以裂解肝細胞,13 000 g離心10 min,去除細胞碎片,取上清備用。超微量核酸蛋白質分析儀測定蛋白質濃度。
1.7間接免疫熒光鑒定P2蛋白與兔肝細胞的結合
分離正常兔肝細胞,PBS洗滌兩次后,稀釋至5×106個·mL-1,鋪覆0.1 mg·mL-1多聚L-賴氨酸包被的細胞板。用含0.1% TritonX100的2%多聚甲醛固定液4 ℃固定肝細胞30 min,PBS洗滌兩次,加入純化的P2蛋白,4 ℃孵育過夜。PBS洗滌三次,加入5~10 μg·mL-11B8單抗200 μL·孔-1,37 ℃孵育1 h。洗滌,加入FITC標記的羊抗鼠IgG,37 ℃ 1 h。PBS洗滌三次,倒置熒光顯微鏡下觀察試驗結果。同時設置陽性對照(VP60蛋白)和陰性對照(重組IFN-γ蛋白)。
1.8Co-IP篩選與P2蛋白相互作用的兔肝細胞蛋白
根據Pierce交聯IP試劑盒使用說明書,P2蛋白、肝細胞蛋白質分別經對照瓊脂糖樹脂預處理,去除與瓊脂糖非特異性結合的蛋白質后,備用。將識別P2區的單克隆抗體1B8結合到protein A/G 瓊脂糖上,經洗滌后,再將誘餌蛋白質P2與之共孵育,洗滌去除未結合的蛋白質。最后將肝細胞蛋白質與結合有1B8和P2的protein A/G瓊脂糖共孵育,4 ℃搖床過夜。低pH洗脫液洗脫蛋白質,并進行SDS-PAGE電泳分析。同時,設置對照A只加入1B8和P2蛋白,用以鑒定抗體和誘餌蛋白質上樣時結合到瓊脂糖上的非特異蛋白;設置對照B以重組IFN-γ蛋白替代P2誘餌蛋白質做陰性對照,以鑒別肝細胞蛋白質與瓊脂糖或1B8抗體的非特異結合蛋白。
1.9質譜鑒定
分別從SDS-PAGE凝膠上切取整個泳道的蛋白質,去除誘餌蛋白質和抗體重輕鏈條帶后,送杭州景杰生物科技有限公司進行LC-MS/MS質譜分析。篩出質譜結果中試驗組與對照組間的差異蛋白質,并通過GO分析注釋差異蛋白質的分子功能和生物進程。通過文獻查閱最終選擇laminin receptor(LamR)進行后期驗證。
1.10LamR基因擴增
以兔肝細胞提取的總RNA反轉錄得到的cDNA為模板,根據GenBank數據庫中 LamR(LOC100354806)的CDS序列設計引物(表1)擴增LamR基因。
1.11pGEX-6p-1-LamR重組表達載體的構建
方法同“1.4”,以pGEX-6p-1為載體,構建pGEX-6p-1-LamR重組質粒,并進行雙酶切和測序鑒定。
1.12GST-LamR重組蛋白質的表達及鑒定
方法同“1.5”,SDS-PAGE及Western blotting鑒定LamR蛋白的表達。
1.13P2蛋白與GST-LamR蛋白的體外pull-down試驗
將GST-LamR或GST(陰性對照)結合到Glutathione樹脂上,再與P2蛋白共孵育,4 ℃ 2 h。還原性谷胱甘肽GSH洗脫并進行SDS-PAGE和Western blotting分析。設置空白對照不加GST蛋白,P2蛋白直接與Glutathione樹脂共孵育。
2結果
2.1pET-32a-P2重組表達載體的構建
以pMD19-T-VP60為模板,PCR擴增P2亞區基因,獲得591 bp與預期大小相符的目的片段(圖1)。雙酶切鑒定重組質粒pET-32a-P2,結果與預期一致(圖2)。測序結果與基因序列完全一致。

M.DL2000 DNA相對分子質量標準; 1.P2亞區基因PCR產物M.2 000 bp ladder;1.The PCR product of P2 subdomain gene圖1 P2亞區基因擴增結果Fig.1 The amplified results of P2 gene
M1.DL2000 DNA相對分子質量標準;1~4.重組質粒pET-32a-P2雙酶切產物;M2.DL15000 DNA相對分子質量標準M1.2 000 bp ladder;1-4.Digestion products of pET-32a-P2 by enzymes;M2.15 000 bp ladder圖2 重組質粒pET-32a-P2的酶切鑒定Fig.2 Enzyme digestion of recombinant plasmid pET-32a-P2
2.2P2重組蛋白質表達、純化及鑒定
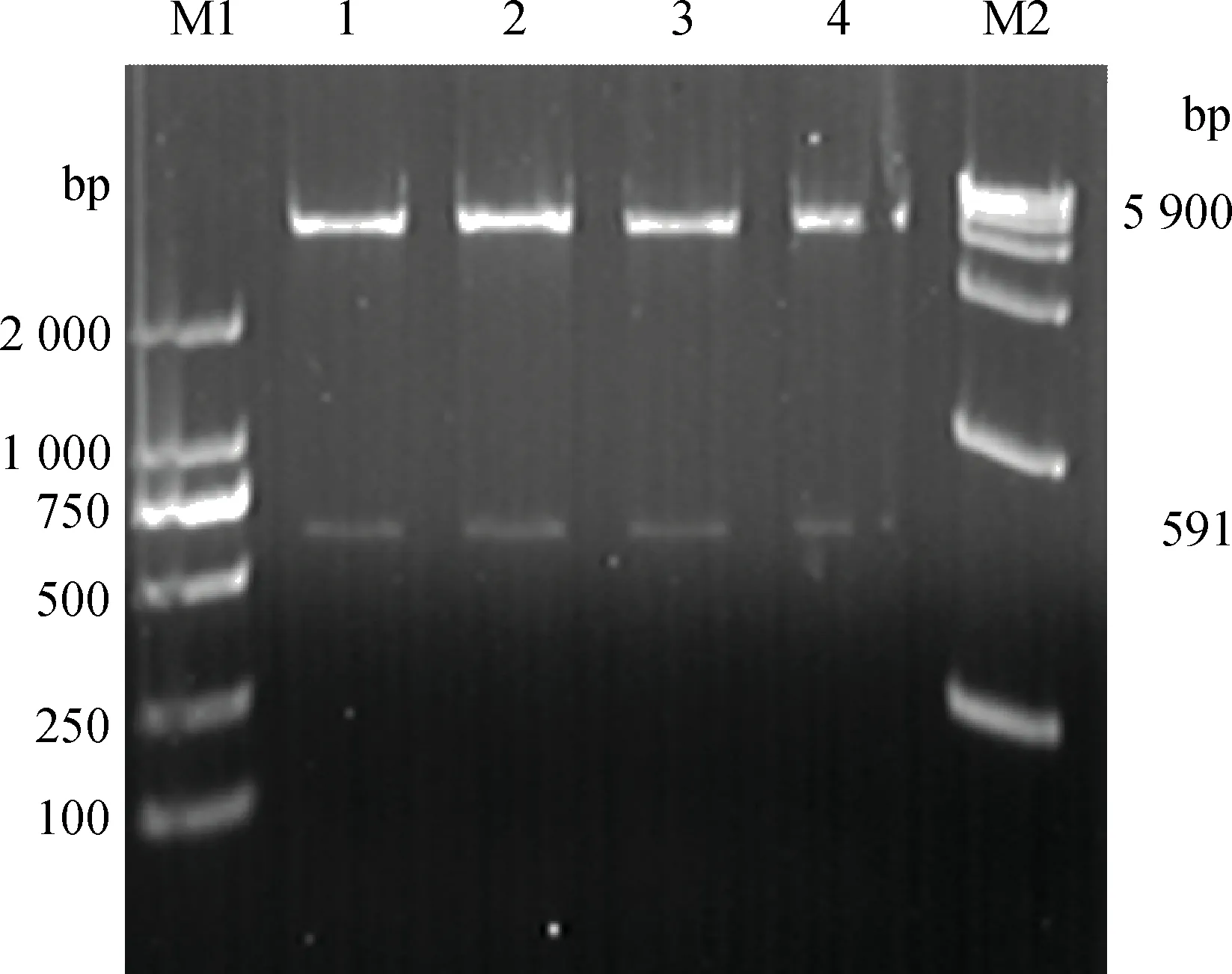
pET-32a-P2重組表達載體轉化BL21感受態細胞,IPTG誘導表達,超聲裂解,離心分離上清和沉淀。SDS-PAGE電泳結果表明,約40 ku處有特異性條帶,與預期大小一致;經HisTrap親和柱純化后,蛋白質雜帶減少,濃度變大(圖3)。Western blotting 鑒定表達的融合蛋白質,結果顯示在40 ku處有特異性條帶。
M.蛋白質相對分子質量標準;1、4.pET-32a(+)空載體表達蛋白質;2、5.重組表達的P2蛋白(未純化);3.純化的P2蛋白M.Protein marker;1,4.Protein expressed by pET-32a(+); 2,5.Recombined P2 protein(unpurified);3.Purified P2 protein圖3 重組蛋白P2的SDS-PAGE和Western blotting鑒定Fig.3 SDS-PAGE and Western blotting analysis of the recombinant P2 protein
2.3P2與肝細胞結合的IFA鑒定
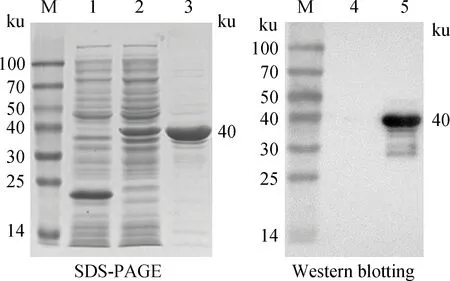
用純化的P2蛋白與兔正常肝細胞共孵育,識別P2的單克隆抗體1B8和羊抗鼠FITC熒光二抗檢測P2蛋白與兔肝細胞的結合。結果顯示P2蛋白能與兔肝細胞發生較強的特異性吸附作用,有特異性綠色熒光,且強于陽性對照組(VP60),而陰性對照組(重組IFN-γ)肝細胞無綠色熒光(圖4)。
A.P2蛋白;B.VP60陽性對照;C.重組IFN-γ陰性對照;D.陰性對照(明場圖)A.P2 protein;B.Positive control (VP60);C.Negative control (IFN-γ);D.Negative control (bright)圖4 P2與肝細胞結合的IFA鑒定Fig.4 Detection the binding of P2 to liver cells by IFA
2.4Co-IP篩選與P2蛋白相互作用的兔肝細胞蛋白質
以pET-32a(+)表達的P2蛋白為誘餌蛋白質,利用Co-IP法釣取與之互作的兔肝細胞蛋白質。SDS-PAGE結果表明,與對照組相比,試驗組有多條肉眼可見的差異條帶,如箭頭所指(圖5)。
2.5質譜分析
分別切下試驗組、對照A、對照B 整個泳道條帶,去除1B8抗體條帶和P2誘餌蛋白質條帶后,分別做LC-MS/MS質譜分析。去除與對照組重復的蛋白質、可信度低的蛋白質(Unique Peptides<2)及未鑒定蛋白質,共篩出26個差異蛋白質,并通過GO分析注釋各蛋白質的分子功能和生物進程(表2)。從分子功能上看,大部分蛋白質為binding蛋白質和具有酶活性的蛋白質。binding蛋白質包括嘌呤核苷酸binding、金屬離子binding、RNA binding、DNA binding、輔因子binding、ATP binding蛋白質。酶活性蛋白質包括具有脫氫酶、氧化酶、轉移酶、磷酸化酶、裂解酶、連接酶等活性的蛋白質。還有小部分蛋白質為核糖體結構成分。從生物進程上來看,這些蛋白質多參與蛋白質的翻譯、修飾、組裝過程以及血紅素、膽固醇等物質的代謝過程,還有部分蛋白質參與細胞凋亡、有氧呼吸及脂質、膽固醇的平衡過程。
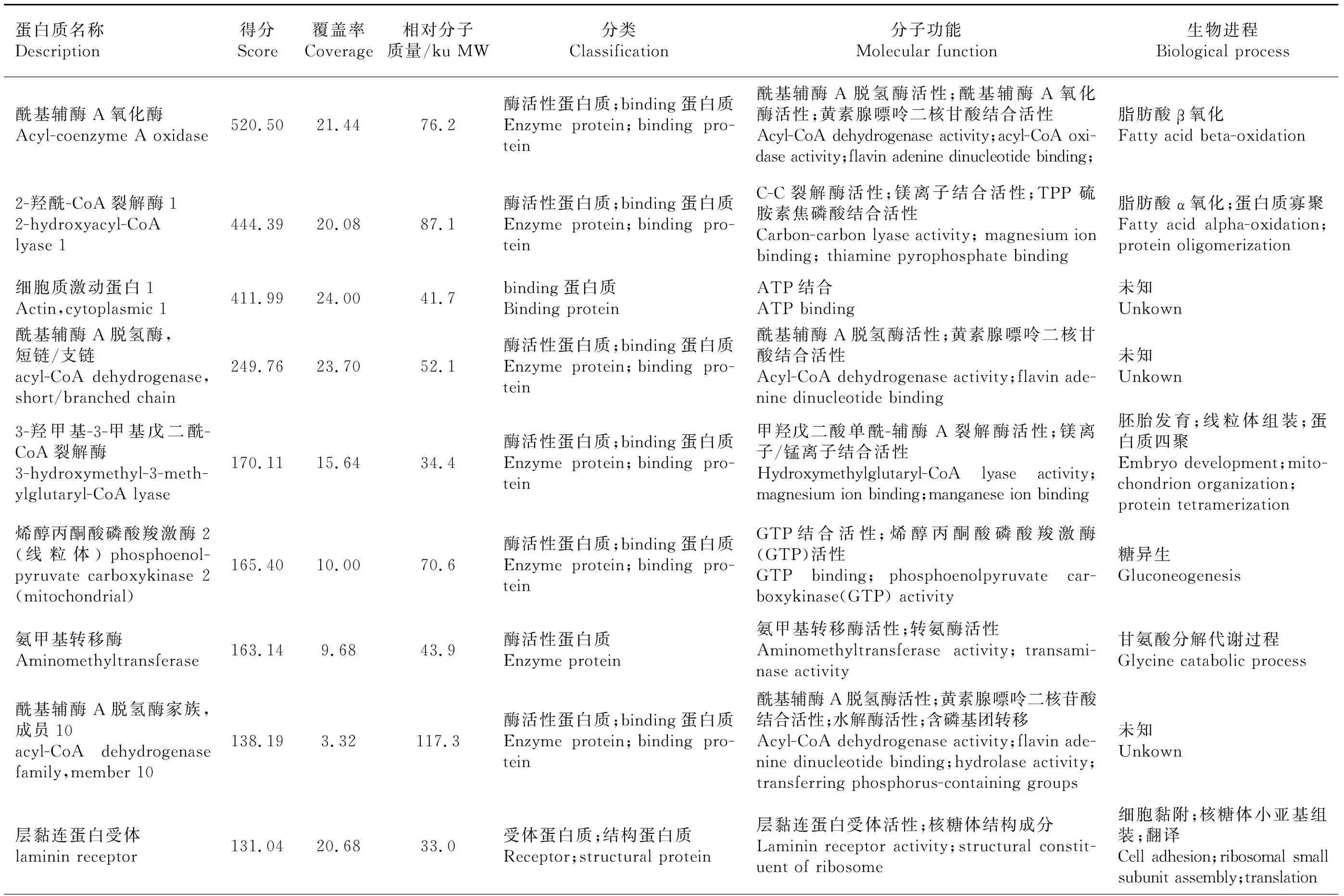
表2 質譜分析結果
(轉下頁)(Continued on the next page)
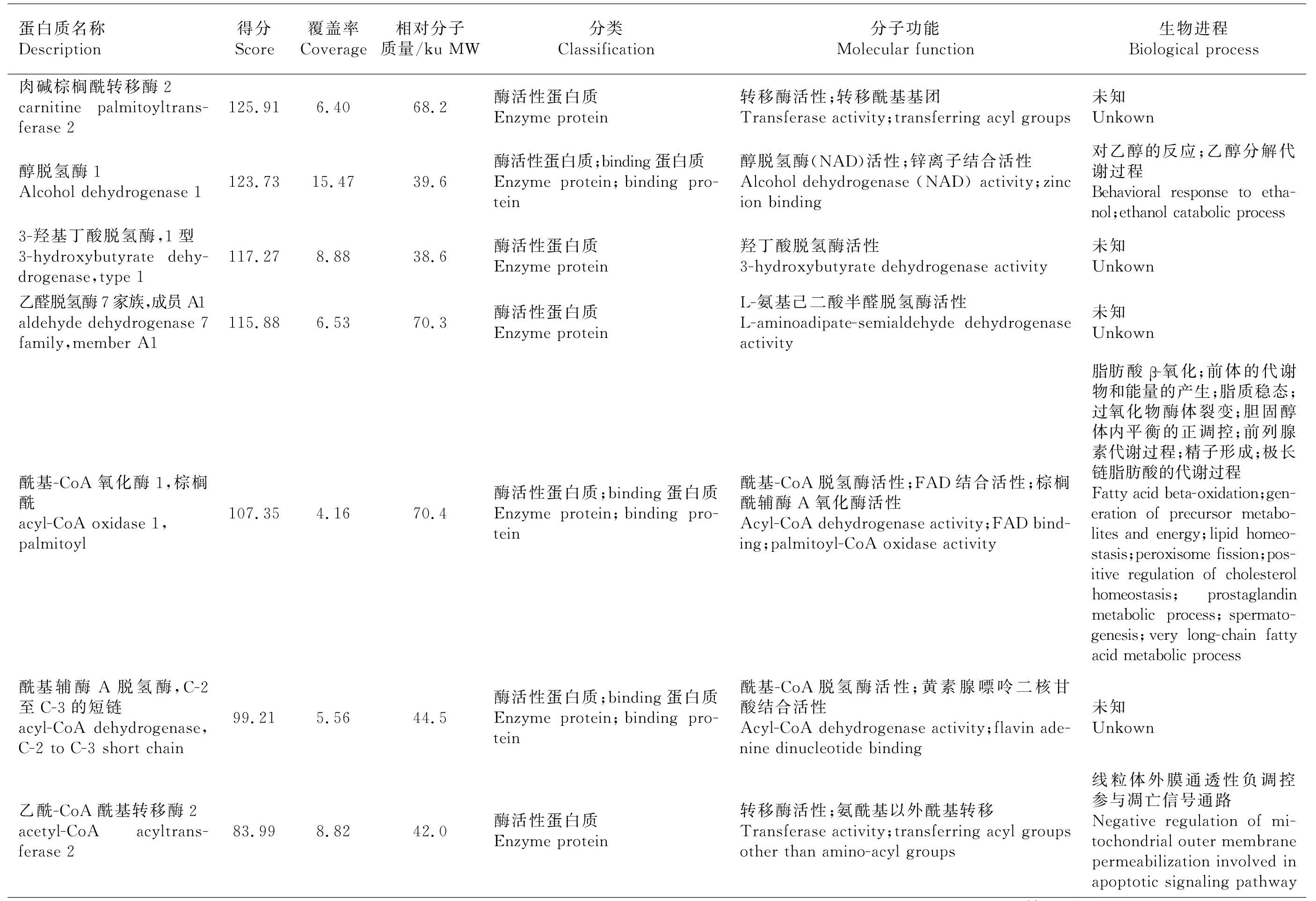
表2(續)Table 2(continued)
(轉下頁)(Continued on the next page)
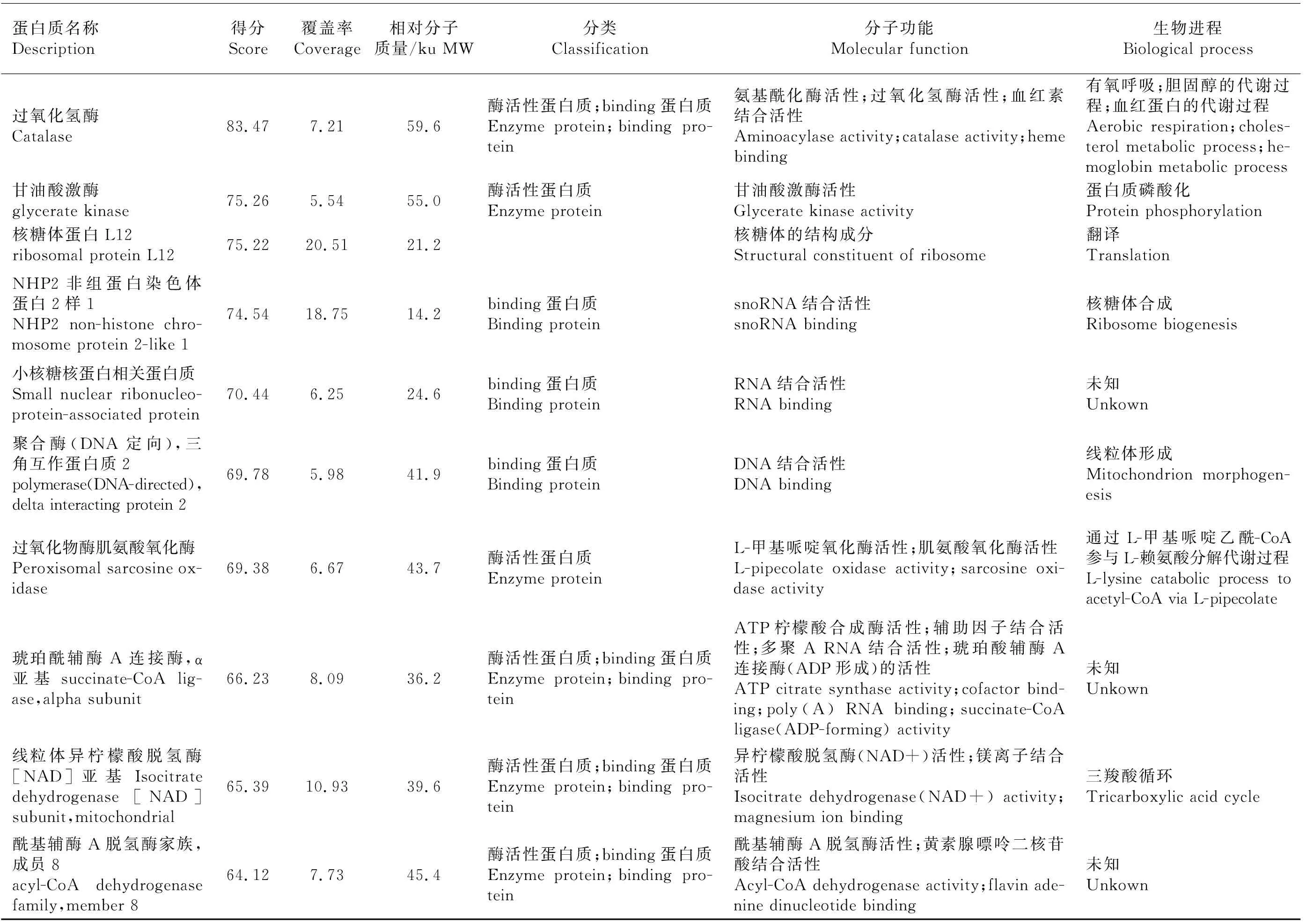
表2(續)Table 2(continued)
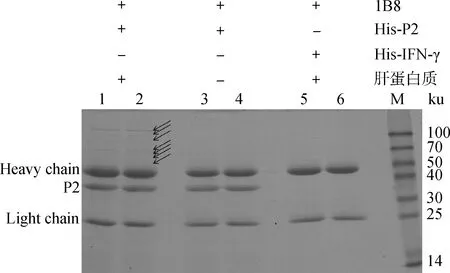
1、2.試驗組樣品;3、4.對照A樣品; 5、6.對照B樣品1,2.The experimental sample;3,4.Control A sample;5,6.Control B sample圖5 Co-IP篩選與P2蛋白相互作用的兔肝細胞蛋白Fig.5 Detection of P2-interacting proteins in rabbit liver cells by Co-IP
通過文獻查閱,選擇laminin receptor(LamR)蛋白進行后期驗證。
2.6pGEX-6p-1-LamR重組表達載體的構建
以兔肝組織總RNA反轉錄得到的cDNA為模板,PCR擴增LamR基因,獲得888 bp與預期大小相符的目的片段(圖6),重組質粒pGEX-6p-1-LamR雙酶切鑒定正確(圖7),測序結果顯示克隆的LamR堿基序列與NCBI兔LamR(LOC100354806)堿基序列相似性為98%,氨基酸序列相似性為97.3%。
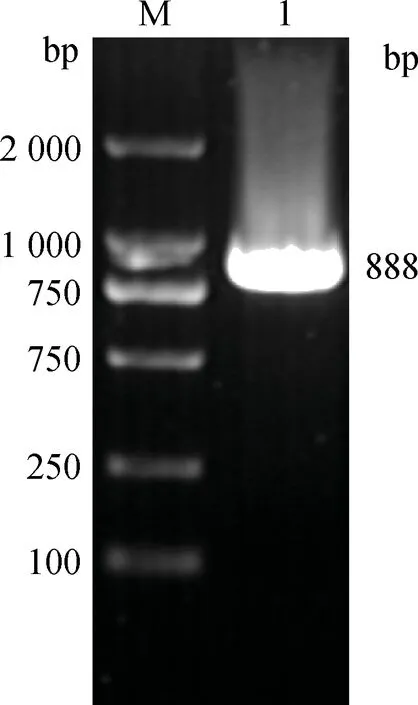
M.DL2000 DNA相對分子質量標準;1.LamR基因RT-PCR產物M.2 000 bp DNA ladder;1.The product of LamR by RT-PCR圖6 LamR基因擴增結果Fig.6 The amplified results of LamR gene
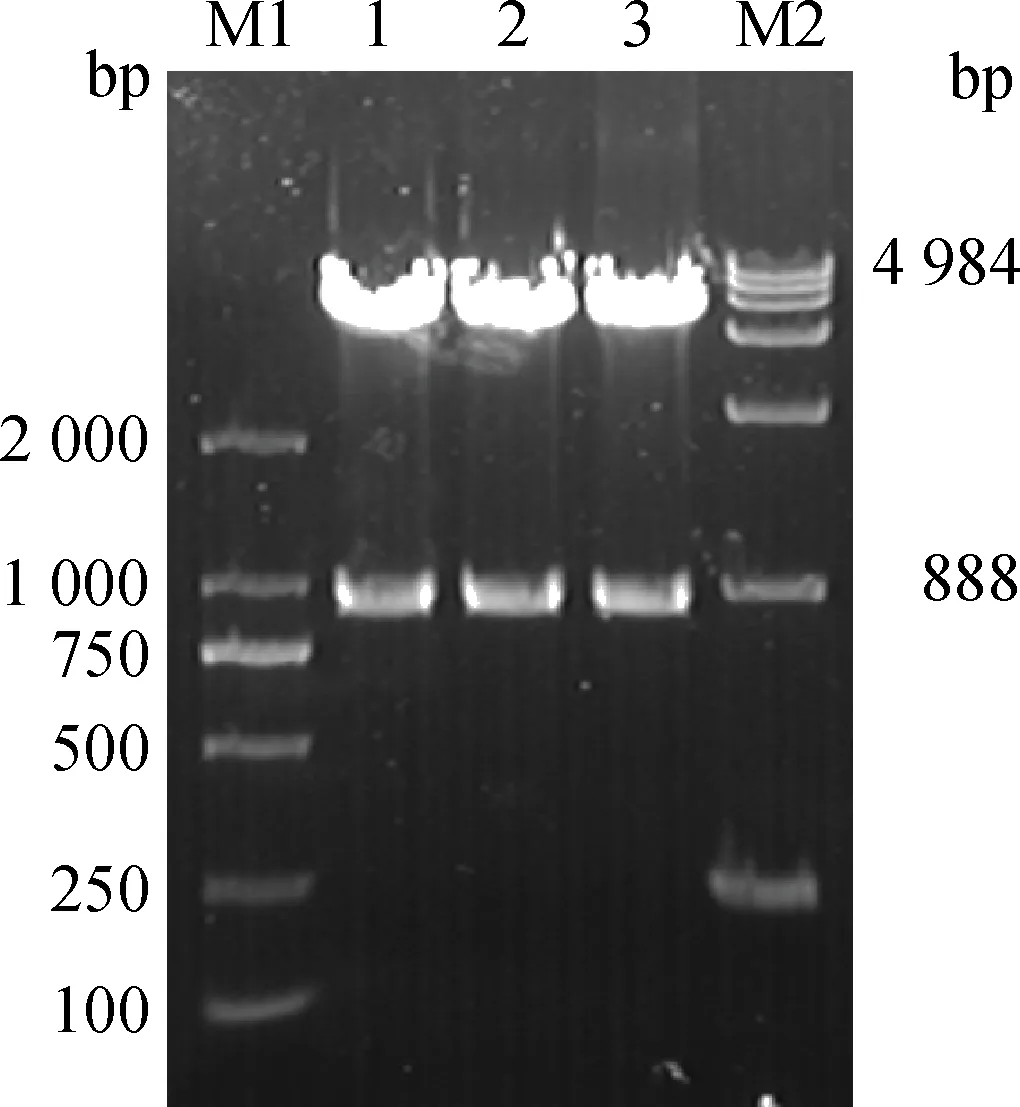
M1.DL2000 DNA相對分子質量標準;1~3.重組質粒pGEX-6p-1-LamR雙酶切產物;M2.DL15000 DNA相對分子質量標準M1.2 000 bp DNA ladder;1-3.Digestion products of pGEX-6p-1-LamR by enzyme;M2.15 000 bp ladder圖7 重組質粒pGEX-6p-1-LamR的酶切鑒定Fig.7 Enzyme digestion of recombinant plasmid pGEX-6p-1-LamR
2.7LamR重組蛋白質表達鑒定
pGEX-6p-1-LamR重組質粒轉化BL21感受態細胞,經IPTG誘導表達,SDS-PAGE和Western blotting分析顯示在60 ku處有GST-LamR特異性條帶,與預期相符(圖8)。
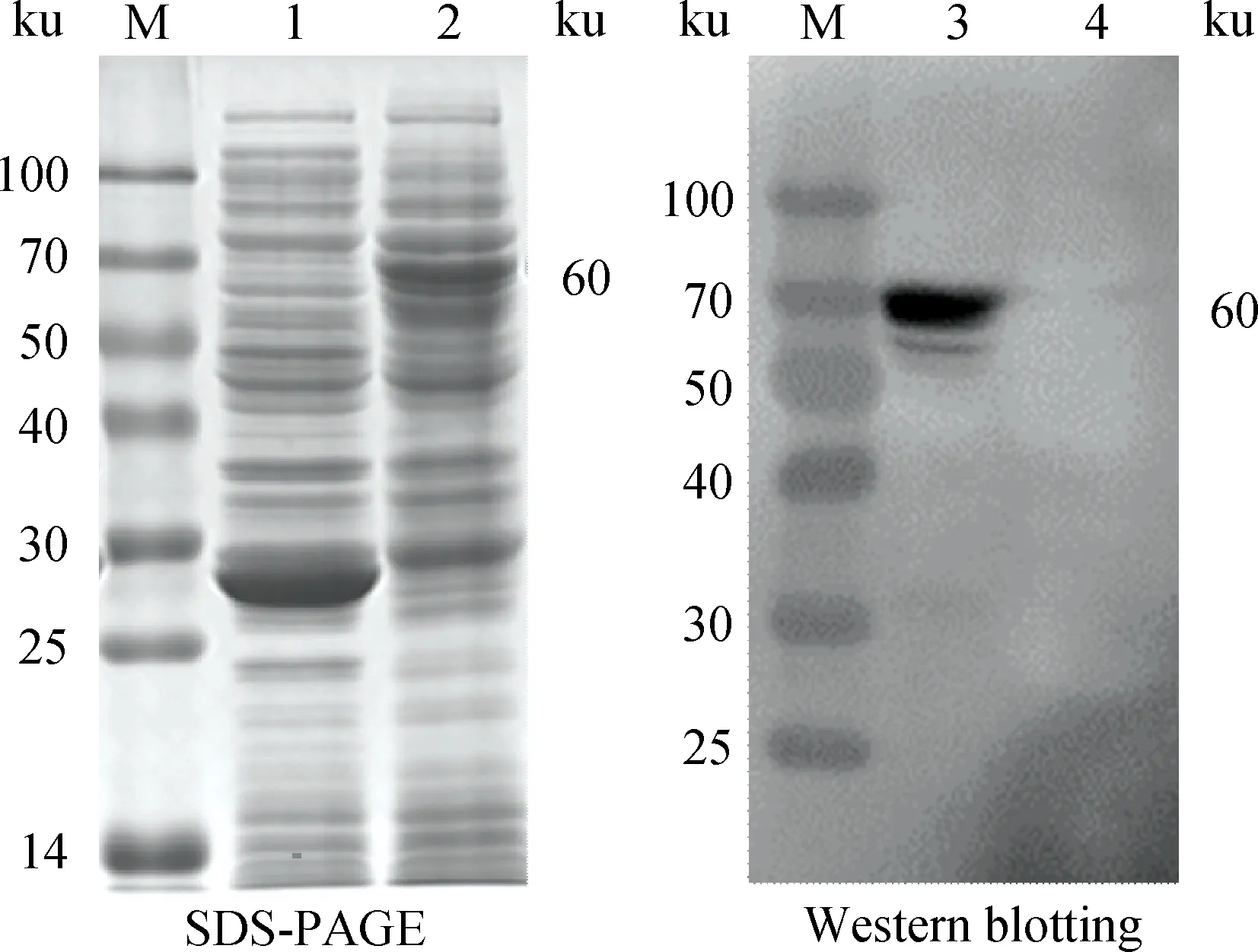
M.蛋白質相對分子質量標準;1、4.pGEX-6p-1空載體誘導表達蛋白質;2、3.重組表達的GST-LamR蛋白質M.Protein marker;1,4.Protein expressed by pGEX-6p-1;2,3.Recombined GST-LamR protein圖8 融合蛋白質GST-LamR的SDS-PAGE和Western blotting鑒定Fig.8 SDS-PAGE and Western blotting analysis of GST-LamR fusion protein
2.8P2蛋白與GST-LamR蛋白的體外pull-down試驗
為驗證LamR與P2是否存在直接相互作用,分別以GST-LamR和GST(陰性對照)為誘餌蛋白質,進行體外GST-pulldown試驗并進行Western blotting分析。結果顯示試驗組有P2蛋白特異性條帶,而陰性對照和空白對照無條帶(圖9)。

1.試驗組(GST-LamR);2.陰性對照(GST);3.空白對照1.Treatment(GST-LamR);2.Negative control(GST);3.Blank control圖9 P2蛋白與LamR蛋白相互作用的體外pull-down驗證Fig.9 Pull-down analysis of P2 interacts with LamR in vitro
3討論
RHDV衣殼蛋白VP60 P區包括P1亞區(238—286、450—466、484—579 aa)和P2亞區(287—449、467—483 aa),其中P2亞區包含7個高變區,推測與受體的結合相關[4]。M.Tan等研究發現諾如病毒(NVs)P2亞區是受體HBGAs結合的關鍵區域[7],R.Chen等也發現杯狀病毒P2亞區與受體識別,宿主特異性和免疫逃避有關[8]。而NVs和RHDV具有相同受體(HBGAs)[9-10]和相似的病毒作用機制。且兩種病毒的衣殼蛋白P2亞區均位于病毒衣殼的最突出區域,最有可能在病毒入侵宿主的過程中率先與宿主細胞接觸并在病毒感染致病過程中發揮重要作用。由此推測RHDV P2亞區可能在病毒感染整個過程中均發揮關鍵作用。本研究構建了P2融合蛋白質,免疫熒光試驗顯示,表達的P2蛋白與肝細胞存在顯著的結合,這表明以P2為誘餌蛋白質釣取的宿主蛋白質可能在病毒結合、侵染和致病過程中具有重要作用。
兔肝是RHDV感染的主要靶器官之一,RHDV能引起兔肝細胞程序性死亡,從而導致嚴重的急性肝炎癥狀[11-12],以兔肝作為研究RHDV P2互作蛋白質的研究對象具有代表性。試驗通過Co-IP法共篩選出兔肝細胞中26個與P2亞區相互作用的蛋白質,GO分析分子功能顯示,這些蛋白質多屬于binding蛋白質和具有酶活性的蛋白質;生物進程顯示,這些蛋白質涉及生理調節、細胞代謝、細胞生長增殖及凋亡等過程。推測RHDV感染肝細胞時,可能通過P2與這些蛋白質的互作,改變肝細胞的正常代謝,致使機體產生一系列的應答反應。
LamR是以二聚體形式存在的跨膜蛋白質,又稱67 ku層黏連蛋白受體,是由37 ku的前體經翻譯后加工形成[13]。LamR被認為是多種病原的受體。E.V.Protopopova等[14]使用獨特型抗體發現LamR是蜱傳腦炎病毒的細胞受體。K.S.Wang等[15]發現LamR是新培斯病毒進入BHK-21的功能受體。蚊源LamR蛋白被認為是委內瑞拉馬腦炎病毒[16]和登革病毒[17-20]的假定受體。在哺乳動物細胞上,LamR又被認為是朊病毒的細胞受體[21-22]。J.Chen等[23]研究發現LamR是經典豬瘟病毒的細胞吸附受體。李茂中通過雙向電泳結合質譜分析對RHDV感染兔和未感染兔肝組織中的差異蛋白質進行分析,也發現LamR是其中的差異蛋白質之一[24],這與本試驗結果一致,但是他們沒有對LamR與RHDV的相互作用進行進一步研究。因此本研究選擇LamR蛋白,對其與P2蛋白的體外結合進行了進一步驗證,證實了兩者在體外確實存在直接的相互作用,但LamR蛋白在病毒入侵宿主的過程中究竟起到怎樣的作用還有待進一步驗證。
4結論
通過Co-IP的方法,以P2為誘餌蛋白質,以RHDV感染的靶器官兔肝為研究對象,結合質譜鑒定共篩選到了26個蛋白質。經分析選取LamR蛋白進行進一步研究。利用GST pull-down試驗驗證了P2與LamR蛋白存在直接的相互作用,為RHDV致病機制的研究提供了線索。
參考文獻(References):
[1]KINNEAR M,LINDE C C.Capsid gene divergence in rabbit hemorrhagic disease virus[J].JGenVirol,2010,91(Pt 1):174-181.
[2]ESTEVES P J,ABRANTES J,CARNEIRO M,et al.Detection of positive selection in the major capsid protein VP60 of the rabbit haemorrhagic disease virus(RHDV)[J].VirusRes,2008,137(2):253-256.
[3]ROHAYEM J,BERGMANN M,GEBHARDT J,et al.Antiviral strategies to control calicivirus infections[J].AntiviralRes,2010,87(2):162-178.
[4]WANG X,XU F,LIU J,et al.Atomic model of rabbit hemorrhagic disease virus by cryo-electron microscopy and crystallography[J].PLoSPathog,2013,9(1):e1003132.
[5]王芳,胡波,任雪楓,等.兔出血癥病毒衣殼蛋白在昆蟲細胞中的表達及對家兔的免疫保護效果[J].畜牧獸醫學報,2008,39(10):1382-1387.
WAGN F,HU B,REN X F,et al.Expression of the capsid protein of rabbit haemorrhagic disease virus in insect cells and its protective efficacy to rabbits[J].ActaVeterinariaetZootechnicaSinica,2008,39(10):1382-1387.(in Chinese)
[6]弗雷謝尼 R I.動物細胞培養—基本技術指南[M].章靜波,徐存拴,等譯.5版.北京:科學出版社,2008:521-522.
R IAN FRESHNEY.Culture of animal cells:a manual of basic technique[M].ZHAGN J B,XU C S,translated.5ed.Beijing:Science Press,2008:521-522.(in Chinese)
[7]TAN M,HUANG P,MELLER J,et al.Mutations within the P2 domain of norovirus capsid affect binding to human histo-blood group antigens:evidence for a binding pocket[J].JVirol,2003,77(23):12562-12571.
[8]CHEN R,NEILL J D,NOEL J S,et al.Inter- and intragenus structural variations in caliciviruses and their functional implications[J].JVirol,2004,78(12):6469-6479.
[9]ABRANTES J,VAN DER LOO W,LE PENDU J,et al.Rabbit haemorrhagic disease(RHD) and rabbit haemorrhagic disease virus(RHDV):a review[J].VetRes,2012,43:12.
[10]NYSTR?M K,LE GALL-RECULé G,GRASSI P,et al.Histo-blood group antigens act as attachment factors of rabbit hemorrhagic disease virus infection in a virus strain-dependent manner[J].PLoSPathog,2011,7(8):e1002188.
[11]ALONSO C,OVIEDO J M,MARTN-ALONSO J M,et al.Programmed cell death in the pathogenesis of rabbit hemorrhagic disease[J].ArchVirol,1998,143(2):321-332.
[12]PARK J H,LEE Y S,ITAKURA C.Pathogenesis of acute necrotic hepatitis in rabbit hemorrhagic disease[J].LabAnimSci,1995,45(4):445-449.
[13]MALINOFF H L,WICHA M S.Isolation of a cell surface receptor protein for laminin from murine fibrosarcoma cells[J].JCellBiol,1983,96(5):1475-1479.
[14]PROTOPOPOVA E V,KONAVALOVA S N,LOKTEV V B.Isolation of a cellular receptor for tick-borne encephalitis virus using anti-idiotypic antibodies[J].VoprVirusol,1997,42(6):264-268.
[15]WANG K S,KUHN R J,STRAUSS E G,et al.High-affinity laminin receptor is a receptor for Sindbis virus in mammalian cells[J].JVirol,1992,66(8):4992-5001.
[16]LUDWIG G V,KONDIG J P,SMITH J F.A putative receptor for Venezuelan equine encephalitis virus from mosquito cells[J].JVirol,1996,70(8):5592-5599.
[17]UPANAN S,KUADKITKAN A,SMITH D R.Identification of dengue virus binding proteins using affinity chromatography[J].JVirolMethods,2008,151(2):325-328.
[18]GAUCZYNSKI S,NIKLES D,EL-GOGO S,et al.The 37-kDa/67-kDa laminin receptor acts as a receptor for infectious prions and is inhibited by polysulfated glycanes[J].JInfectDis,2006,194(5):702-709.
[19]TIO P H,JONG W W,CARDOSA M J.Two dimensional VOPBA reveals laminin receptor(LAMR1) interaction with dengue virus serotypes 1,2 and 3[J].VirolJ,2005,2:25.
[20]THEPPARIT C,SMITH D R.Serotype-specific entry of dengue virus into liver cells:identification of the 37-kilodalton/67-kilodalton high-affinity laminin receptor as a dengue virus serotype 1 receptor[J].JVirol,2004,78(22):12647-12656.
[21]SAKOONWATANYOO P,BOONSANAY V,SMITH D R.Growth and production of the dengue virus in C6/36 cells and identification of a laminin-binding protein as a candidate serotype 3 and 4 receptor protein[J].Intervirology,2006,49(3):161-172.
[22]GAUCZYNSKI S,PEYRIN J M,HAIK S,et al.The 37-kDa/67-kDa laminin receptor acts as the cell-surface receptor for the cellular prion protein[J].EMBOJ,2001,20(21):5863-5875.
[23]CHEN J,HE W R,SHEN L,et al.The laminin receptor is a cellular attachment receptor for classical Swine Fever virus[J].JVirol,2015,89(9):4894-4906.
[24]李茂中.人工感染兔出血癥病毒兔肝臟的蛋白質組學分析[D].北京:中國農業科學院,2012.
LI M Z.Proteomics analysis of rabbit liver after infection by rabbit hemorrhagic disease virus[D].Beijing:Chinese Academy of Agricultural Sciences,2012.(in Chinese)
(編輯白永平)
Screening and Identification of RHDV VP60 P2 Domain Interacting Proteins in Rabbit Liver Cells
LI Teng1,2,JIANG Ping1*,WANG Fang1,2*,SONG Yan-hua2,HU Bo2,FAN Zhi-yu2,WEI Hou-jun2,QIU Ru-long2,LIU Xing2
(1.KeyLaboratoryofAnimalDiseasesDiagnosticandImmunology,CollegeofVeterinaryMedicine,NanjingAgriculturalUniversity,Nanjing210095,China;2.KeyLaboratoryofVeterinaryBiologicalEngineeringandTechnologyofMinistryofAgriculture,NationalCenterforEngineeringResearchofVeterinaryBio-products,InstituteofVeterinaryMedicine,JiangsuAcademyofAgriculturalSciences,Nanjing210014,China)
Abstract:According to reports,VP60 P2 subdomain of RHDV may play an important role in the viral infection. In this study,we used co-immunoprecipitation(Co-IP) with RHDV VP60 P2 expressed inE.colias bait protein and mass spectrometry analysis to screen the RHDV interacting hepatocellular proteins.Laminin receptor (LamR),as receptors of many other viruses,was identified and verified as potential interacting partner of P2.LamRgene was amplified by RT-PCR from rabbit liver cells.The recombinant LamR protein was expressed inE.coliand used in GST pull-down to verify the interaction of LamR and P2.Here we screened 26 proteins as potential P2 interacting proteins.After verification,the protein LamR could interact with P2 directly.We found that RHDV VP60 P2 interacted with LamR which would contributed to further study of the infectious process and pathogenesis of RHDV.
Key words:rabbit hemorrhagic disease virus (RHDV);VP60 P2 subdomain;co-immunoprecipitation; laminin receptor (LamR)
doi:10.11843/j.issn.0366-6964.2016.03.018
收稿日期:2015-08-21
基金項目:現代農業產業技術體系建設兔體系病毒病預防與控制崗位(CARS-44-C-1); 國家自然科學基金(31502073)
作者簡介:李騰(1990-),女,湖南邵東人,碩士生,主要從事家畜傳染病學研究,Tel:025-84390337,E-mail: 2013107045@njau.edu.cn *通信作者:姜平,教授, E-mail:jiangp@njau.edu.cn;王芳,研究員,E-mail:rwangfang@126.com
中圖分類號:S855.3;S852.659.6
文獻標志碼:A
文章編號:0366-6964(2016)03-0549-10
