基于生物量的冬小麥越冬前植株地上部形態結構模型
2016-07-15 05:45:18陳昱利楊平張文宇張偉欣諸葉平李世娟鞏法江畢海濱岳霆曹宏鑫
作物學報 2016年5期
關鍵詞:模型
陳昱利楊 平張文宇張偉欣諸葉平李世娟鞏法江畢海濱岳 霆曹宏鑫,*
1淄博市農業科學研究院, 山東淄博 255033;2江蘇省農業科學院農業經濟與信息研究所/數字農業工程技術研究中心, 江蘇南京210014;3中國農業科學院農業信息研究所, 北京 100081
?
基于生物量的冬小麥越冬前植株地上部形態結構模型
陳昱利1楊 平1張文宇2張偉欣2諸葉平3李世娟3鞏法江1畢海濱1岳 霆1曹宏鑫2,*
1淄博市農業科學研究院, 山東淄博 255033;2江蘇省農業科學院農業經濟與信息研究所/數字農業工程技術研究中心, 江蘇南京210014;3中國農業科學院農業信息研究所, 北京 100081
摘 要:越冬前植株地上部形態建成是冬小麥株型構建和可視化的重要基礎。為定量分析冬小麥越冬前植株地上部形態結構參數與器官生物量的關系, 以濟麥22、泰農18和魯原502為材料, 于2013—2014和2014—2015年度開展品種和施氮試驗。利用2013—2014年度越冬前植株地上部形態結構參數和器官生物量等數據構建了冬小麥越冬前植株地上部形態結構模型。經2014—2015年度數據檢驗, 除葉鞘長和葉弦長模型精度略低外, 葉長、最大葉寬、葉切角和葉弦角模型精度均較高, 所建模型可較好地模擬不同品種與各施氮水平冬小麥越冬前植株地上部形態結構。
關鍵詞:冬小麥; 形態結構; 生物量; 植株地上部; 模型
本研究由國家高技術研究發展計劃(863計劃)項目(2013AA102305-1)資助。
This study was supported by the National High Technology Research and Development Program of China (2013AA102305-1).*通訊作者(Corresponding author)∶ 曹宏鑫, E-mail∶ caohongxin@hotmail.com
第一作者聯系方式∶ E-mail∶ luckydogcyl@163.com
URL∶ http∶//www.cnki.net/kcms/detail/11.1809.S.20160314.1444.008.html
合理株型結構是小麥高產、穩產、優質的有效保證, 地上部形態結構是小麥生長發育過程與株型結構的全面展現, 而小麥生長模型與形態結構模型則是小麥生長發育過程與株型結構的定量表達。因此, 將小麥生長模型與形態模型有機結合, 建立基于生物量的小麥植株形態結構參數模型是功能-結構小麥模型研究的重要內容, 對小麥株型設計與栽培調控具有重要參考意義。
目前, 關于小麥形態結構模型的研究, 主要集中在小麥形態與環境因素的定量關系和小麥植株可視化方面。Evers等[1-2]通過分析群體密度和遮陰對春小麥生長發育的影響, 構建了春小麥形態參數模型, 并對小麥發育的三維結構模型 ADEL-wheat參數進行了量化; Fournier等[3]在ADEL-wheat模型基礎上, 分析了冬、春小麥初始參數的差異。張文宇等[4-5]利用系統分析方法和動態建模技術, 以生長度·日為尺度, 構建了小麥主莖葉片及莖鞘夾角動態模擬模型; 并通過分析小麥主莖葉型和莖型指標與環境因素的定量關系, 模擬分層葉面積、葉向值等株型指標動態變化規律。陳國慶等[6]通過分析小麥莖鞘生長過程與環境因素的關系, 構建了小麥葉鞘和節間生長過程的動態模擬模型。Mabille等[7]構建了小麥籽粒形態參數模型。伍艷蓮等[8]借助于OpenGL圖形平臺, 實現了小麥器官-個體-群體三層次的形態可視化; 談峰等[9]在構建基于形態特征參數的小麥根系三維形態模型基礎上, 實現了根系生長可視化。雷曉俊等[10]通過對小麥穗形態結構觀測分析,提出了基于形態特征參數的麥穗幾何模型及可視化實現方法。上述研究中, 所建小麥形態模型大多未考慮同化物分配與小麥植株形態模型的定量關系,缺乏冬小麥生長模型和形態模型的有效結合。
隨著作物形態結構模型不斷發展, 功能-結構小麥模型已成為小麥形態模型與可視化研究的一個重要方向[11]。目前, 基于生物量的作物功能-結構模型已有研究, 主要是通過確定作物器官生物量與形態參數的定量關系和作物形態參數之間的內在聯系, 進而建立形態結構參數模型和實現植株可視化[12-16],而基于生物量的小麥植株地上部形態結構參數模型報道較少。本文在前人研究基礎上, 以器官生物量為尺度, 通過分析冬小麥越冬前地上部植株形態參數與器官生物量的定量關系, 將冬小麥生長模型和形態模型結合, 建立冬小麥越冬前地上部植株形態結構模型。為冬小麥生長模型與植株地上部形態結構模型結合, 進而為建立功能—結構小麥模型奠定基礎。
1 材料與方法
1.1 供試材料
選取 3個代表性小麥品種, 即濟麥 22 (株型緊湊, 山東省農業科學院作物研究所育成)、泰農 18(中間型, 泰安市泰山區瑞豐作物育種研究所、山東農業大學農學院育成)和魯原 502 (株型松散, 山東省農業科學院原子能農業應用研究所、中國農業科學院作物科學研究所育成)。
1.2 試驗設計
于2013年10月至2015年6月在山東省淄博市農業科學研究院試驗農場開展品種與施氮試驗, 設3個品種, 5個氮肥處理, 3次重復, 共計45個小區。土壤為褐土, 0~30 cm耕層含有機碳32.14g kg-1、全氮1.62 g kg-1、速效磷14.40 mg kg-1、速效鉀150.32 mg kg-1、pH 8.16。采用裂區設計, 主區為品種(V),即V1 (濟麥22)、V2 (泰農18)和V3 (魯原502), 副區為施氮水平(N)。小區面積10.0 m × 1.5 m=15.0 m2, 行道寬0.5 m。基施有機肥15 000 kg hm-2、P2O5112.5 kg hm-2、K2O 112.5 kg hm-2。其他管理同大田高產栽培管理。
2013年10月至2014年6月, 設N1 (0 kg hm-2)、N2 (84.375 kg hm-2)、N3 (168.75 kg hm-2)、N4 (253.125 kg hm-2)和N5 (337.5 kg hm-2) 5個施氮水平。50%作基肥, 在播種前施入; 50%作追肥, 在拔節期施入。2013年10月 7日播種, 基本苗 225萬hm-2。2014年10月至2015年6月, 根據上年度試驗結果, 為了更切合生產實際, 設N1 (0 kg hm-2)、N2 (75 kg hm-2)、N3 (150 kg hm-2)、N4 (225 kg hm-2)和N5 (300 kg hm-2) 5個施氮水平。50%作基肥, 在播種前施入; 50%作追肥, 在拔節期施入。2014年10 月12日播種, 基本苗225萬 hm-2。第1年試驗數據用于建立模型, 第2年試驗數據用于模型檢驗。
1.3 植株形態參數采集與分析
自三葉期標記葉位, 每隔10 d選取每處理長勢一致的植株 5株, 用直尺、游標卡尺等測定主莖葉片長度、葉片最大寬度、葉鞘長、葉弦長、莖葉夾角(葉切角、葉弦角)(圖 1)等形態指標。然后, 分不同葉位將葉片、葉鞘等分裝, 105℃下殺青30 min, 再80℃烘至恒重后稱重。
葉片長度為葉片伸直狀態下自葉片基部至葉尖的直線距離; 葉片最大寬度為葉片的最大葉寬值,一般在葉片中部; 葉弦長為葉片自然伸展狀態下自葉片基部至葉尖的空間直線距離; 葉切角為葉片基部切線與主莖的夾角(∠AOP, 圖 1); 葉弦角為葉弦與主莖的夾角(∠BOP, 圖1)。各參數均以3次測量取平均值。
采用Microsoft Excel 2007和Sigmaplot v10.0統計分析軟件處理試驗數據。
1.4 模型檢驗
利用根均方差(RMSE)、平均絕對誤差(da)和平均絕對誤差與觀察值平均數的比值(dap)統計量檢驗模型, 并繪制觀測值與模擬值的1︰1關系圖, 以檢驗模型。
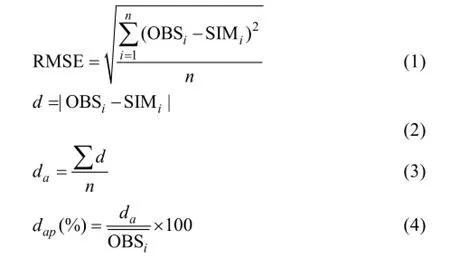
式中, OBSi為實測值,為實測值平均值, SIMi為模擬值, d為絕對誤差, n為樣本容量。
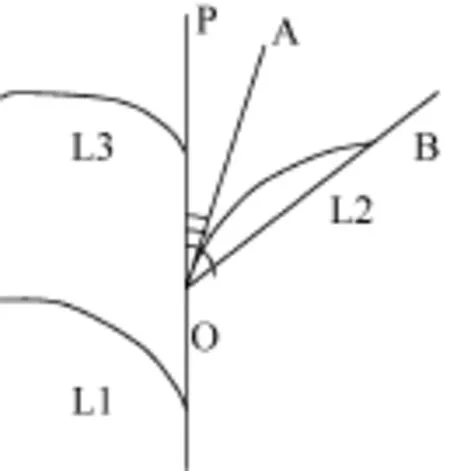
圖1 葉片角度結構示意圖Fig.1 Schematic diagram of leaf blade angles
2 結果與分析
2.1 模型描述
2.1.1 葉長模型 生物量作為小麥葉片生長發育的物質基礎, 是小麥葉片形態形成的關鍵。2013—2014年試驗數據表明, 越冬前冬小麥不同葉位葉片長度隨其生物量增加呈線性增長的趨勢, 品種間有較大差異(圖2)。根據葉片伸長與葉片生物量變化的關系, 主莖葉片長度可由如下公式計算。
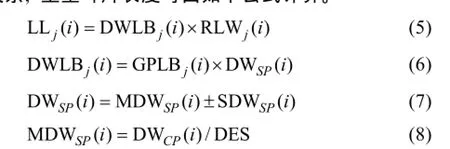
式中, j表示主莖葉位, i表示出苗后的天數, LLj(i)為出苗后第i天主莖第j葉位葉片長度(cm), DWLBj(i)為出苗后第 i天主莖第 j葉位葉片干生物量(g),RLWj(i)為出苗后第i天主莖第j葉位葉片長度和干生物量的比值(cm g-1), CPLBj(i)為出苗后第i天主莖第 j葉位葉片干生物量與單株地上部干生物量的比值(g g-1), DWSP(i)為出苗后第i天單株地上部干生物量(g), MDWSP(i)為出苗后第 i天單株地上部平均干生物量(g), SDWSP(i)為出苗后第i天單株地上干生物量標準誤差(g), DWCP(i)為出苗后第i天單位面積植株地上部干生物量(g m-2), DES為種植密度(m-2)。

圖2 2013-2014年不同處理葉片長度隨葉片干重的變化趨勢Fig.2 Changes in the average single blade length by the average single blade dry weight for different treatments in 2013-2014
隨著葉位的增加, 越冬前冬小麥主莖不同葉位RLW 逐漸變小, 呈指數函數變化趨勢, 品種間無顯著差異(圖3-A); 而越冬前主莖不同葉位CPLB先變大后變小, 在第3和第4葉位達到最大, 呈現二次曲線變化趨勢, 品種間無顯著差異(圖3-B)。不同葉位葉片RLW和CPLB可用如下公式表示。
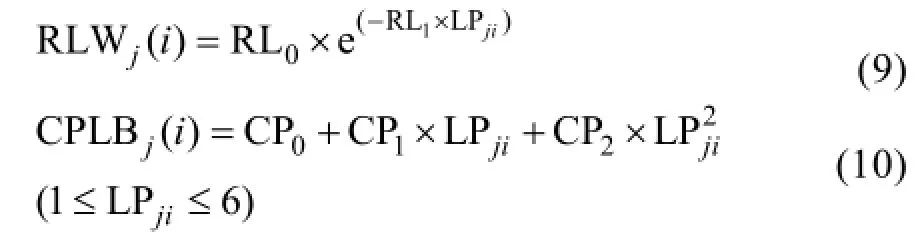
式中, RLWj(i)為出苗后第i天主莖第j葉位葉片長度和干生物量的比值(cm g-1); CPLBj(i)為出苗后第i天主莖第 j葉位葉片干生物量與單株地上部干生物量的比值(g g-1); LPji為出苗后第i天主莖第j葉片的葉位; RL0、RL1、CP0、CP1、CP2為模型參數, 由試驗數據獲得(表1)。
2.1.2 最大葉寬、葉鞘長和葉弦長模型 越冬前冬小麥主莖不同葉位葉片最大葉寬、葉鞘長和葉弦長均隨該葉位葉片伸長呈一次函數的增長趨勢, 品種間無顯著差異(圖4)。
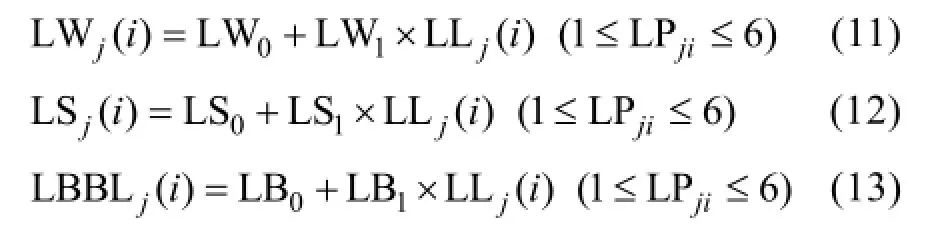
式中, LWj(i)、LSj(i)和LBBLj(i)分別為出苗后第i天主莖第j葉片的最大葉寬(cm)、葉鞘長(cm)和葉弦長(cm); LLj(i)為出苗后第 i天主莖第 j葉位葉片長度(cm); LPji為出苗后第i天主莖第j葉片的葉位; LW0、LW1、LS0、LS1、LB0、LB1為模型參數(表1)。
2.1.3 葉片角度模型 葉片生物量是葉片角度形成的重要生理學和物理學基礎[10], 因此將某葉位葉片葉切角和葉弦角引入模型, 并計算該葉位葉片干生物量與葉切角和葉弦角的比值 RTWj(i)和RBWj(i)。越冬前冬小麥主莖不同葉位葉片的RTWj(i)和 RBWj(i)隨葉位的增加而降低, 呈指數函數變化趨勢(圖5)。
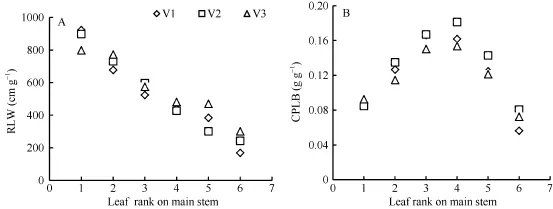
圖3 葉片長度與地上部干生物量比值(A)和葉片干生物量與單株地上部干生物量比值(B)隨葉位的變化趨勢(2013-2014)Fig.3 Changes in RLW(A) and CPLB (B) with the leaf rank on main stem (2013-2014)
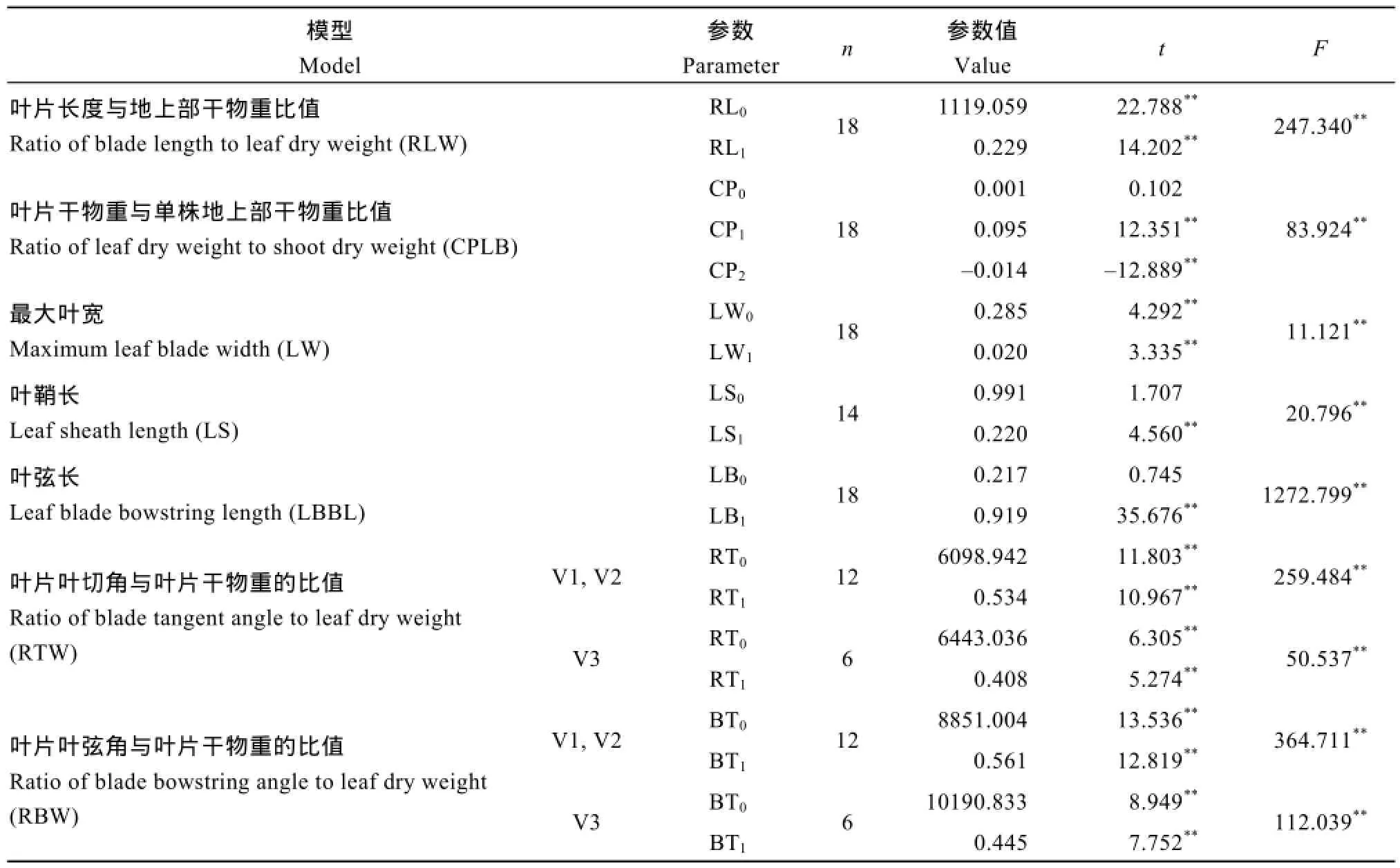
表1 各模型參數值及其統計檢驗Table 1 Parameters of various models and their statistical test
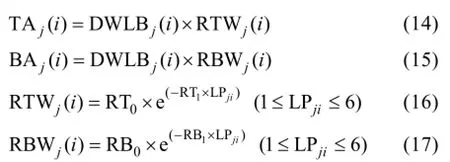
式中, TAj(i)和BAj(i)分別為第j葉第i天的葉切角(°)和葉弦角(°), DWLBj(i)為出苗后第i天第j葉位葉片干生物量(g), RTWj(i)和RBWj(i)分別為第i天第j葉的葉切角和葉弦角與葉片干物重的比值(° g-1), LPji為出苗后第i天主莖第j葉片的葉位, RT0、RT1、RB0、RB1為模型參數(表1)。
2.2 模型檢驗
采用獨立試驗資料對所建冬小麥越冬前植株形態參數模型進行檢驗, 結果冬小麥越冬前植株地上部主莖形態結構參數葉長、最大葉寬、葉鞘長、葉弦長、葉切角和葉弦角的實測值與模擬值的吻合程度較好(表2和圖6), 各模型r值在0.708~0.998之間(P < 0.001)。葉長、最大葉寬、葉弦長以及葉切角模型的dap值在10%~20%之間, 表明模型精度較好, 而葉鞘長及葉弦角模型精度較低(dap> 20%), 表明模型有改進空間。
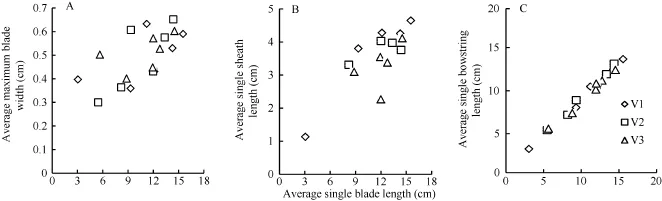
圖4 最大葉寬(A)、葉鞘長(B)和葉弦長(C)隨葉長的變化趨勢(2013-2014)Fig.4 Changes in the maximum single blade width (A), single sheath length (B), and single bowstring length (C) with the leaf blade length (2013-2014)

圖5 葉片葉切角與葉片干生物量比值(A)和葉弦角與葉片干生物量比值(B)隨葉位的變化趨勢(2013-2014)Fig.5 Changes in RTW (A) and RBW (B)with the leaf rank on main stem (2013-2014)
3 討論
隨著作物形態模型不斷發展, 功能—結構植物模型(functional-structural plant model, FSPM)已經成為作物形態模型與可視化研究的一個重要方向[17]。FSPM 是作物生長模型之功能與作物形態模型之結構的有機結合, 通過將作物生理過程描述與 3D顯示相結合, 不斷強化對作物生理機制、品種基因型與環境響應研究[11]。本研究通過分析小麥地上部植株形態參數與生物量的定量關系, 將小麥生長模型和形態模型結合, 建立了基于生物量的小麥植株地上部植株形態結構參數模型。生物量形成受小麥品種和環境因素(光、溫度、水分、CO2、施肥等)綜合影響, 而生物量可通過小麥生長模型估算并響應品種和環境因素差異[18]。目前, 針對小麥形態結構參數與環境因素的定量關系以及植株可視化已開展了較多形態結構模型方面的研究[19-21], 例如, 譚子輝[19]通過分析小麥葉、莖、蘗、穗等形態建成與其生理發育時間的關系, 初步研究了小麥植株形態建成的模擬模型。然而, 現有形態結構模型缺少與生長模型的結合, 而生物量作為小麥生長發育的一個重要指標, 對小麥植株形態建成具有重要作用, 是小麥生長模型和形態結構模型有機結合的橋梁。除生物量外, 本研究中在模型中引入了RLW、CPLB、RTW、RBW等參數, 均具有一定生物學意義。
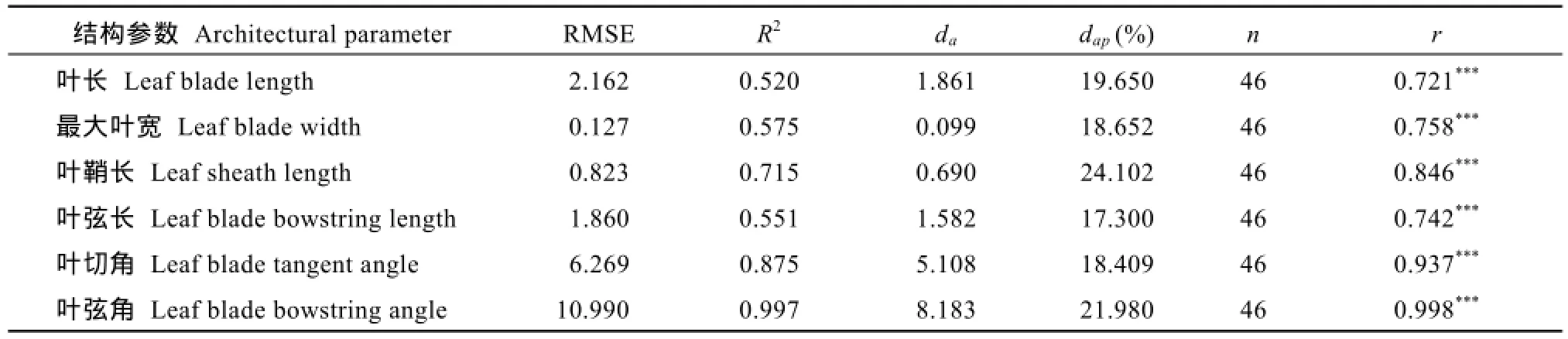
表2 冬小麥越冬前植株地上部形態結構模型觀察值與模擬值比較的統計參數Table 2 Comparison of statistical parameters of simulation and observation in winter wheat aboveground architectural parameter models before overwintering
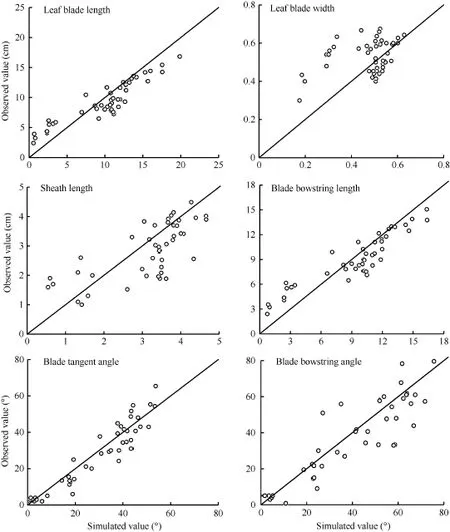
圖6 實測值與模擬值比較(2014-2015)Fig.6 Comparison between observed and simulated values (2014-2015)
本研究表明, 冬小麥葉片長度和生物量存在線性關系, 這與曹宏鑫等[12]在水稻和張偉欣等[18]在油菜上的研究結果相似。葉片角度是小麥株型結構的一個重要指標, 隨著分蘗的發生, 葉鞘與主莖之間也會形成一定角度[3], 使葉片角度發生變化, 故在葉片角度(葉切角和葉弦角)的模擬上存在一定誤差。由于在越冬前, 小麥植株分蘗較小, 本文只涉及小麥單株主莖形態結構, 所建模型是否適合小麥分蘗,還有待檢驗; 小麥植株形態結構除了受品種、氮素等因素影響外, 還受水分[22]、播期[23]、密度[4]等多種因素影響。特別是播期和種植密度對小麥形態結構有較大影響[24], 下一步擬將把播期和密度作為研究重點, 在不同種植密度下, 確定小麥不同葉位葉片干物質積累與分配的變化規律并建立相應的形態結構參數模型。此外, 葉形和葉曲線模型也是小麥葉片形態結構模型研究的重要內容[25-29], 本文尚未涉及。
冬小麥越冬前地上部植株形態結構指標主要包括數量指標和質量指標2個部分。本研究只考慮了冬小麥越冬前地上部植株形態數量指標與生物量的定量關系, 并未涉及冬小麥越冬前地上部植株形態質量指標(葉片含氮量、含水量、葉片厚度等)的差異。
4 結論
在研究冬小麥植株地上部器官生物量分配的基礎上, 通過分析冬小麥越冬前植株地上部各形態參數指標與生物量的定量關系及其各形態參數之間的內在聯系, 建立了基于生物量的冬小麥越冬前植株地上部各形態參數模型。除葉鞘長和葉弦角模型精度較低外, 所建其他形態結構參數模型準確性均較高, 具有一定的機理性和普適性。
References
[1]Evers J B, Vos J, Fournier C, Andrieu B, Chelle M, Struik P C.An architectural model of spring wheat∶ evaluation of the effects of population density and shading on model parameterization and performance.Ecol Model, 2007, 200∶ 308-320
[2]Evers J B, Vos J, Fournier C, Andrieu B, Chelle M, Struik P C.Towards a generic architectural model of tillering in Gramineae,as exemplified by spring wheat (Triticum aestivum).New Phytol,2005, 166∶ 801-812
[3]Fournier C, Andrieu B, Ljutovac S, Saint-Jean S.ADEL-wheat∶ a 3D architectural model of wheat development.In∶ Hu B G, Jaeger M, eds.2003 International Symposium on Plant Growth Modeling, Simulation, Visualization, and Their Applications.Beijing,Tsinghua University Press, 2003.pp 54-63
[4]張文宇, 湯亮, 朱相成, 楊月, 曹衛星, 朱艷.基于過程的小麥莖鞘夾角動態模擬.應用生態學報, 2011, 22∶ 1765-1770 Zhang W Y, Tang L, Zhu X C, Yang Y, Cao W X, Zhu Y.Dynamic simulation of wheat stem-sheath angle based on process.Chin J Appl Ecol, 2011, 22∶ 1765-1770 (in Chinese with English abstract)
[5]張文宇, 湯亮, 姚鑫鋒, 楊月, 曹衛星, 朱艷.基于過程的小麥株型指標動態模擬.中國農業科學, 2012, 45∶ 2364-2374 Zhang W Y, Tang L, Yao X F, Yang Y, Cao W X, Zhu Y.Process-based simulation model for growth dynamics of plant type index in wheat.Sci Agric Sin, 2012, 45∶ 2364-2374 (in Chinese with English abstract)
[6]陳國慶, 朱艷, 曹衛星.小麥葉鞘和節間生長過程的模擬研究.麥類作物學報, 2005, 25∶ 71-74 Chen G Q, Zhu Y, Cao W X.Modeling leaf sheath and internode growth dynamics in wheat.J Triticeae Crops, 2005, 25∶ 71-74 (in Chinese with English abstract)
[7]Mabille F, Abecassis J.Parametric modelling of wheat grain morphology∶ a new perspective.J Cereal Sci, 2003, 37∶ 43-53
[8]伍艷蓮, 曹衛星, 湯亮, 朱艷, 劉慧.基于 OpenGL的小麥形態可視化技術.農業工程學報, 2009, 25(1)∶ 121-126 Wu Y L, Cao W X, Tang L, Zhu Y, Liu H.OpenGL-based visual technology for wheat morphology.Trans CSAE, 2009, 25(1)∶121-126 (in Chinese with English abstract)
[9]談峰, 湯亮, 胡軍成, 姜海燕, 曹衛星, 朱艷.小麥根系三維形態建模及可視化.應用生態學報, 2011, 22∶ 137-143 Tan F, Tang L, Hu J C, Jiang H Y, Cao W X, Zhu Y.Three-dimensional morphological modeling and visualization of wheat root system.Chin J Appl Ecol, 2011, 22∶ 137-143 (in Chinese with English abstract)
[10]雷曉俊, 湯亮, 張永會, 姜海燕, 曹衛星, 朱艷.小麥麥穗幾何模型構建與可視化.農業工程學報, 2011, 27(3)∶ 179-184 Lei X J, Tang L, Zhang Y H, Jiang H Y, Cao W X, Zhu Y.Geometric model and visualization of wheat spike.Trans CSAE, 2011,27(3)∶ 179-184 (in Chinese with English abstract)
[11]曹宏鑫, 趙鎖勞, 葛道闊, 劉永霞, 劉巖, 孫金英, 岳延濱, 張智優, 陳昱利.作物模型發展探討.中國農業科學, 2011, 44∶3520-3528 Cao H X, Zhao S L, Ge D K, Liu Y X, Liu Y, Sun J Y, Yue Y B,Zhang Z Y, Chen Y L.Discussion on development of crop models.Sci Agric Sin, 2011, 44∶ 3520-3528 (in Chinese with English abstract)
[12]Cao H X, Liu Y, Liu Y X, Hanan J S, Yue Y B, Zhu D W, Lu J F,Sun J Y, Shi C L, Ge D K, Wei X F, Yao A Q, Tian P P, Bao T L.Biomass-based rice (Oryza sativa L.) aboveground architectural parameter models.J Integr Agric, 2012, 11∶ 1621-1632
[13]劉巖, 陸建飛, 曹宏鑫, 石春林, 劉永霞, 朱大威, 孫金英, 岳延濱, 魏秀芳, 田平平, 包太林.基于生物量的水稻葉片主要幾何屬性模型研究.中國農業科學, 2009, 42∶ 4093-4099 Liu Y, Lu J F, Cao H X, Shi C L, Liu Y X, Zhu D W, Sun J Y, Yue Y B, Wei X F, Tian P P, Bao T L.Main geometrical parameter models of rice blade Based on biomass.Sci Agric Sin, 2009, 42∶4093-4099 (in Chinese with English abstract)
[14]宋有洪, 郭焱, 李保國, de Reffye P.基于器官生物量構建植株形態的玉米虛擬模型.生態學報, 2003, 23∶ 2579-2586 Song Y H, Guo Y, Li B G, de Reffye P.Virtual maize model∶ II.Plant morphological constructing based on organ biomass accumulation.Acta Ecol Sin, 2003, 23∶ 2579-2586 (in Chinese with English abstract)
[15]劉永霞, 岳延濱, 劉巖, 曹宏鑫, 葛道闊, 魏秀芳.不同品種和氮肥條件下水稻根系主要幾何參數動態的量化研究.中國農業科學, 2010, 43∶ 1782-1790 Liu Y X, Yue Y B, Liu Y, Cao H X, Ge D K, Wei X F.Quantitative research of dynamic models of the main geometric parameters of rice root system of different varieties under different nitrogen conditions.Sci Agric Sin, 2010, 43∶ 1782-1790 (in Chinese with English abstract)
[16]陳超, 潘學標, 張立禎, 龐艷梅.棉花地上部生長的功能—結構模型研究.作物學報, 2012, 38∶ 2237-2245 Chen C, Pan X B, Zhang L Z, Pang Y M.Functional and structural model for above-ground growth in cotton.Acta Agron Sin,2012, 38∶ 2237-2245 (in Chinese with English abstract)
[17]曹宏鑫, 石春林, 金之慶.植物形態結構模擬與可視化研究進展.中國農業科學, 2008, 41∶ 669-677 Cao H X, Shi C L, Jin Z Q.Advances in researches on plant morphological structure simulation and visualization.Sci Agric Sin, 2008, 41∶ 669-677 (in Chinese with English abstract)
[18]張偉欣.基于生物量的油菜植株地上部形態結構模型研究.南京農業大學碩士學位論文, 江蘇南京, 2013 Zhang W X.Study on Biomass-Based Rapeseed Aboveground Morphological Structure Model.MS Thesis of Nanjing Agricultural University, Nanjing, China, 2013 (in Chinese with English abstract)
[19]譚子輝.小麥植株形態建成的模擬模型研究.南京農業大學碩士學位論文, 江蘇南京, 2006 Tan Z H.Researches of Simulation Models for Wheat Plant Morphological Forming.MS Thesis of Nanjing Agricultural University,Nanjing, China, 2006 (in Chinese with English abstract)
[20]劉炳成, 劉偉, 劉俐華, 金弋.冬小麥根系生長的三維仿真模擬.華中科技大學學報(自然科學版), 2005, 33(9)∶ 65-67 Liu B C, Liu W, Liu L H, Jin G.Three-dimensional visual simulation of wheat root system growing.J Huazhong Univ Sci Technol (Nat Sci Edn), 2005, 33(9)∶ 65-67 (in Chinese with English abstract)
[21]趙春江, 王紀華, 吳華瑞, 黃文江, 鄭文剛.小麥葉形空間分布的模擬模型及推理系統.農業工程學報, 2002, 18(5)∶221-225 Zhao C J, Wang J H, Wu H R, Huang W J, Zheng W G.Simulation models and deduction system for interspace description of wheat leaf shape.Trans CSAE, 2002, 18(5)∶ 221-225 (in Chinese with English abstract)
[22]田夢雨, 李丹丹, 戴廷波, 姜東, 荊奇, 曹衛星.水分脅迫下不同基因型小麥苗期的形態生理差異.應用生態學報, 2010 ,21∶ 41-47 Tian M Y, Li D D, Dai T B, Jiang D, Jing Q, Cao W X.Morphological and physiological differences of wheat genotypes at seedling stage under water stress.Chin J Appl Ecol, 2010, 21∶ 41-47 (in Chinese with English abstract)
[23]李存東, 曹衛星, 戴廷波, 嚴美春.小麥不同品種和播期對發育階段的效應.應用生態學報, 2001, 12∶ 218-222 Li C D, Cao W X, Dai T B, Yan M C.Effects of different varieties and sowing dates on development stages of wheat.Chin J Appl Ecol, 2001, 12∶ 218-222 (in Chinese with English abstract)
[24]趙志范.冬小麥分蘗缺位產生原因及其控制的研究.北京農業科學, 1984, (3)∶ 34-40 Zhao Z F.Reason for tiller deficiency in winter wheat and its management.Beijing Agric Sci, 1984, (3)∶ 34-40 (in Chinese)
[25]鄭文剛, 郭新宇, 趙春江, 王紀華.玉米葉片幾何造型研究.農業工程學報, 2004, 20(1)∶ 152-154 Zheng W G, Guo X Y, Zhao C J, Wang J H.Geometry modeling of the maize leaf canopy.Trans CSAE, 2004, 20(1)∶ 152-154 (in Chinese with English abstract)
[26]孟軍, 郭新宇, 趙春江.小麥地上部器官幾何造型與可視化研究.麥類作物學報, 2009, 29∶ 106-109 Meng J, Guo X Y, Zhao C J.Geometry modeling and visualization of above-ground organs of wheat.J Triticeae Crops, 2009,29∶ 106-109 (in Chinese with English abstract)
[27]趙春江, 鄭文剛, 郭新宇, 王紀華.玉米葉片三維形態的數學模擬研究.生物數學學報, 2004, 19(4)∶ 493-496 Zhao C J, Zheng W G, Guo X Y, Wang J H.The computer simulation of maize leaf.J Biomath, 2004, 19(4)∶ 493-496 (in Chinese with English abstract)
[28]鄧旭陽, 郭新宇, 周淑秋, 鄭文剛.玉米葉片形態的幾何造型研究.中國圖象圖形學報, 2005, 10∶ 637-641 Deng X Y, Guo X Y, Zhou S Q, Zheng W G.Study on the geometry modeling of corn leaf morphological formation.J Image & Graphics, 2005, 10∶ 637-641 (in Chinese with English abstract)
[29]石春林, 朱艷, 曹衛星.水稻葉曲線特征的機理模型.作物學報, 2006, 32∶ 656-660 Shi C L, Zhu Y, Cao W X.A quantitative analysis on leaf curvature characteristics in rice.Acta Agron Sin, 2006, 32∶656-660 (in Chinese with English abstract)
DOI:10.3724/SP.J.1006.2016.00743
收稿日期Received()∶ 2015-08-12; Accepted(接受日期)∶ 2016-03-02; Published online(網絡出版日期)∶ 2016-03-14.
Aboveground Architecture Model Based on Biomass of Winter Wheat before Overwintering
CHEN Yu-Li1, YANG Ping1, ZHANG Wen-Yu2, ZHANG Wei-Xin2, ZHU Ye-Ping3, LI Shi-Juan3, GONG Fa-Jiang1, BI Hai-Bin1, YUE Ting1, and CAO Hong-Xin2,*
1Zibo Academy of Agricultural Sciences, Zibo 255033, China;2Institute of Agricultural Economics and Information/Engineering Research Center for Digital Agriculture, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China;3Agricultural Information Institute, Chinese Academy of Agricultural Sciences, Beijing 100081, China
Abstract:The aboveground morphogenesis is an important basis of plant morphological construction and visualization for winter wheat before overwintering.For quantitatively analyzing the relationship between the aboveground architectural parameters and organ biomass of winter wheat before overwintering, field experiments with different varieties (Jimai 22, Tainong 18, and Luyuan 502) and nitrogen levels were carried out in 2013-2014 and 2014-2015 wheat growth seasons.Simulation models for aboveground architectural of winter wheat before overwintering were built with the 2013-2014 dataset of aboveground architectural parameters before overwintering and organ biomass and validated by the 2014-2015 dataset, showing the models exhibited satisfactory predictions for leaf blade length, leaf maximum blade width, leaf blade tangent angle, and leaf blade bowstring angle,except for leaf sheath length and leaf bowstring length.The models built in this study are suitable to simulate the aboveground architecture of winter wheat varieties before overwintering under different nitrogen levels.
Keywords:Winter wheat; Morphological structure; Biomass; Aboveground plant; Model
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
